
若尔盖高寒草地退化沙化过程中土壤养分与团聚体结构的变化特征
2019-09-21 02:40:10陈秋捷张楠楠仲波陈冬明孙庚刘琳
生态科学 2019年4期
陈秋捷, 张楠楠, 仲波, 陈冬明, 孙庚,*, 刘琳
若尔盖高寒草地退化沙化过程中土壤养分与团聚体结构的变化特征
陈秋捷1,2, 张楠楠2, 仲波2, 陈冬明3, 孙庚2,*, 刘琳1,*
1. 四川农业大学动科院草学系, 成都 611130 2. 中国科学院山地生态恢复与生物资源利用重点实验室, 生态恢复与生物多样性保育四川省重点实验室, 中国科学院成都生物研究所, 成都 610041 3. 四川省甘孜藏族自治州畜牧业科学研究所, 康定 626000
若尔盖地处青藏高原东缘, 是黄河上游重要的水源涵养地。近来受过度放牧、鼠虫害等影响, 若尔盖高寒草地出现严重退化, 部分草地已完全沙化。草地沙化已成为若尔盖高寒草地生态安全所面临的重大问题。研究以若尔盖典型的沙化地区麦溪乡为研究对象, 以植被覆盖率为沙化梯度划分标准, 分别划分未退化、轻度退化、中度退化、重度退化和极度退化5种不同退化阶段草地, 探究退化沙化过程中草地土壤养分与团聚体结构变化特征。结果表明: (1)退化沙化过程中, 土壤养分含量总体随着草地沙化程度的加剧而显著下降(<0.05)。(2)退化沙化过程中土壤团聚体结构发生明显变化, 团聚体平均重量直径(MWD)、几何平均直径(GWD)随着沙化程度的加剧而显著降低(<0.05); 各粒级团聚体百分比仅5—2 mm粒级团聚体在重度退化阶段发生显著降低变化(<0.05), 其他粒级并未表现出明显的变化趋势。(3)团聚体MWD与微生物生物量、有机质、全氮、硝态氮、有机碳均呈显著或极显著正相关, 与pH呈极显著负相关(<0.01)。在整个草地退化沙化过程中, 土壤养分含量显著下降, 土壤结构受到严重破坏, 二者之间变化关系紧密。草地沙化治理与防护过程中, 要兼顾土壤结构修复与养分输入, 处理好土壤团聚体结构稳定与养分转化间的交互关系。
若尔盖; 草地沙化; 土壤养分; 土壤团聚体
0 前言
若尔盖高寒草地地处青藏高原东缘, 是我国重要牧区, 也是长江、黄河流域重要的水源涵养及生物多样性集中区, 具有重要的生态地位[1]。近年来受人为及自然因素影响, 若尔盖高寒草地严重退化, 大量沙化地出现。调查表明, 1994—2009年, 若尔盖沙化草地面积增加了28.1 %, 2009年沙化草地面积达到82.19×104hm2, 占川西北幅员面积25.85×106hm2的3.18 %[2]。草地沙化对高寒草地生态有着严重的影响。草地退化沙化过程中, 土壤养分严重流失, 团聚结构遭到严重破坏, 土壤砂粒含量增加, 直接影响着草地植物生长[3], 而草地植物生长受阻又将影响着土壤恢复[4]。在高寒草地草地生态系统退化沙化过程中, 土壤退化往往滞后于植物退化, 其退化后恢复时间要远远长于草地植物的恢复时间[5], 这将严重影响高寒草地生态系统健康与平衡, 给草地生态可持续发展带来极大阻碍[6]。
草地沙化将直接影响草地生产力, 探究土壤养分含量在草地退化沙化过程中变化规律, 对沙化草地的恢复与管理有着重要的理论意义[7-8]。土壤团聚体作为土壤结构的基本单元, 影响着土壤的孔隙性、持水能力, 透气性等, 其稳定性是反应土壤结构状况的重要指标之一[9]。目前对川西北若尔盖高寒草地研究主要集中在退化现状及其驱动力等方面, 对于土壤化学和微生物肥力与土壤物理结构间的变化及相互影响关系的研究较少[8]。因此, 了解掌握若尔盖高寒草地退化沙化过程中, 土壤养分及土壤团聚体变化特征及其相互关系是极其必要的。
本研究以若尔盖沙化草地为研究对象, 探究高寒草地退化沙化过程中, 不同沙化程度草地土壤养分、土壤团聚体结构的变化特征, 以期了解若尔盖高寒草地的退化沙化过程中土壤自身养分与结构间的内在变化关系, 为若尔盖沙化草地恢复提供理论依据。
1 材料与方法
1.1 研究地点
本研究试验点位于四川省北部的若尔盖县麦溪乡(102°11′—102°18′E、33°56′—33°58′N), 平均海拔3430 m, 属典型高原寒温带湿润季风气候。年均气温1.3 ℃, 年降水量615 mm, 年蒸发量1352.4 mm, 无绝对无霜期。该地降雨多集中于5月下旬至7月中旬, 年降雨量656.8 mm, 年均相对湿度69 %。每年10月初土地开始冻结, 5月中旬完全解冻。沙化草地主要出现冬季放牧集中的地区, 包括某些古河道。该地区草地沙化较轻的地段, 植物组成与原生植被组成相似; 严重沙化的地段, 植物组成相对简单, 并出现沙生苔草(spp)、赖草(()Tzvel.)等沙生植物。未经管理沙化草地将最终演变为大型的流动沙丘, 无任何植被类型。
1.2 研究方法
于2016年8月, 在麦溪乡境内随机选择大型裸露沙丘, 以沙丘中心为起点向外围草地扩散取样。以植被覆盖率为沙化梯度划分标准[10], 将植被覆盖率为70%—100 %、50%—70 %、30%—50 %、10%—30 %、0%—10 %分别划分为潜在沙化草地(未沙化地)、轻度沙化草地、中度沙化草地、重度沙化草地、极度沙化草地5个梯度[11-13]。每个处理内随机设置4个1×1 m的样方, 采用直径5 cm的土钻, 在样方内按对角线取样法随机取3钻0—20 cm土层土壤, 3钻混合放入自封袋中, 带回实验室过2 mm筛, 4℃保存备用。
1.3 分析方法
土壤可溶性有机碳、氮用土壤浸提液直接在TOC分析仪上测定[14]; 微生物生物量碳(microbial biomass carbon, MBC)采用氯仿熏蒸—K2SO4浸提法测定, 转换系数K取0.38, 微生物生物量氮(microbial biomass nitrogen, MBN)采用氯仿熏蒸—K2SO4提取测定, 转换系数K取0.45[15]; 有机质采用重铬酸钾—浓硫酸油浴法[16]; 土壤pH值采用1 : 2.5土水比进行混合后, 土壤悬浮液用pH测量仪(PHB—4)进行测定; 硝态氮用酚二磺酸分光光度法(GB7480—87)测定; 铵态氮用纳氏试剂比色法(GB7479—87)测定[14]。
土壤团聚体分析采用湿筛法: 将土壤样品过8 mm的筛子, 称取60 g土壤样品(干重), 小心放入土壤团聚体湿筛测定仪的套筛(>2000 um)进行团聚体分级, 筛套从上至下分别为(2000 um, 250 um, 53 um),土壤完全浸入去离子水中(套筛不完全浸入水中), 静置5分钟, 开启团聚体分析仪 (振幅3 cm, 25 Hz, 2 min), 小心分离不同粒级套筛上的土壤, 用于土壤团聚体比例测定[17-19]。
团聚体的平均重量直径(MWD)和几何平均直径(GWD)计算采用公式[20]:


1.4 数据分析
采用SPSS 17.0软件进行统计分析, 通过单因素方差分析(One—way ANOVA)和LSD多重比较法进行差异显著性检验, 检验不同草地退化程度下若尔盖沙化草地土壤养分含量及土壤团聚体的变化。文中数据均为平均值(mean)±标准误(SE), 文中所有图在Origin(version 8.0)中绘制。
2 结果与分析
2.1 退化沙化过程土壤化学特征变化
由表1可看出, 铵态氮总体变化较为波动, 未出现明显的规律变化, 与未退化处理相比, 仅中度退化与极度退化处理阶段有显著的下降(<0.05), 分别下降了30.07 %和28.85 %。硝态氮呈现出随沙化程度加深而显著降低(<0.05), 较未退化处理相比较, 极度退化减少了83.67 %; 可溶性全氮变化趋势与硝态氮相近, 与未退化相比较, 其轻度退化与极度退化处理阶段分别减少了41.4 %和73.38 %。可溶性有机氮与可溶性有机碳都仅在退化初期(未退化至轻度退化)显著下降(<0.05), 分别下降了56.97 %、51.72 %。土壤pH表现出随沙化程度的加深而显著上升趋势<0.05), 在极度退化阶段达到最大值(8.16±0.02)。较未退化样地, 极度退化样地pH值上升了7.11 %。
图1可知, 随着沙化程度愈深, 土壤有机质及全氮均表现出显著下降趋势(<0.05)。其变化区间分别在(3.15±0.22—11.09±2.43) g·kg–1(图1a)和(0.2±0.01—0.64±0.10) g·kg–1(图1b)之间, 最大值均出现在未退化处理阶段。与未退化阶段相比较, 各退化沙化处理阶段有机质含量分别下降了41.66 %、55.73 %、57.17 %、71.6 %; 全氮含量分别下降26.56 %、45.31 %、50 %、68.75 %。可见有机质、全氮均在极度退化阶段下降百分比最为突出。
2.2 退化沙化过程土壤微生物生物量碳、氮含量的变化
由图2可知, 不同退化沙化阶段土壤微生物碳、氮均表现出显著下降(<0.05)。MBC在轻度退化处理阶段开始出现显著下降(<0.05), 相比未退化处理下降了57.63 %, 但并未在其余退化处理阶段继续出现显著下降变化()。相较未退化处理, MBN在轻度退化处理阶段开始出现显著下降, 并在极度退化处理阶段再次出现显著下降, 下降幅度分别为64.09 %和89.65 %。
2.3 退化沙化过程中土壤团聚体的百分比变化
表2可知, 在整个退化沙化过程中, 2—0.250 mm、0.250—0.053 mm粒级始终显著高于5—2 mm、0.053—0 mm粒级(<0.05), 而5—2 mm及0.053—0 mm粒级之间并未出现显著的变化(>0.05)。同时, 随着沙化程度愈深, 0.250—0.053 mm粒级在中度退化处理阶段开始表现出显著高于2—0.250 mm粒级的变化趋势(<0.05)。与未退化处理相比较, 退化沙化过程中2—0.250 mm、 0.250—0.053 mm及0.053—0 mm粒级并未发生明显的增减变化(P>0.05), 但5—2 mm粒级在重度退化处理阶段出现了显著的降低变化(< 0.05), 下降了67.65 %; 并在极度退化阶段未筛得该粒级团聚体。
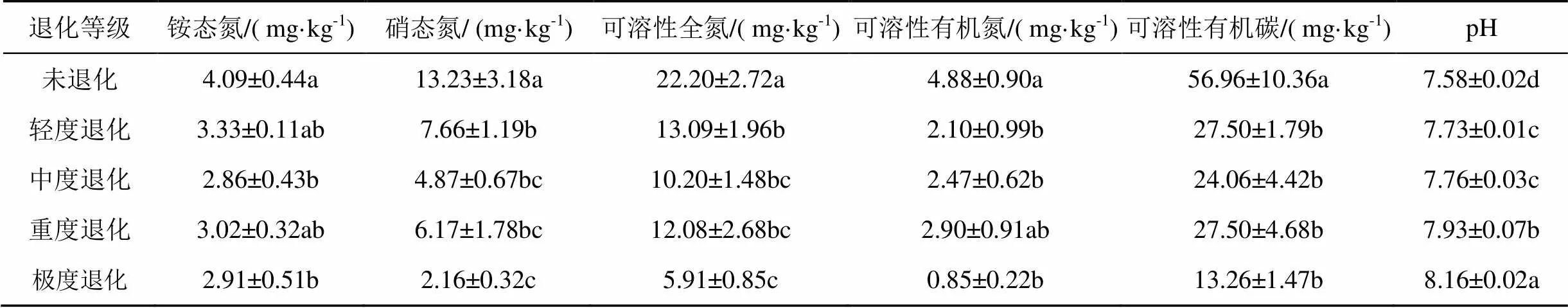
表1 不同退化沙化阶段土壤理化性质
注:不同小写字母表示各处理间在< 0.05水平上差异显著。

图1 不同退化沙化阶段土壤有机质(a)、全氮含量(b)
Figure 1 Soil organic matter (a) and total nitrogen content (b) under different degrees of degradation

图2 不同退化沙化阶段土壤微生物生物量碳(a)、氮(b)
Figure 2 Soil microbial biomass carbon (a) and nitrogen (b) under different vegetation coverage
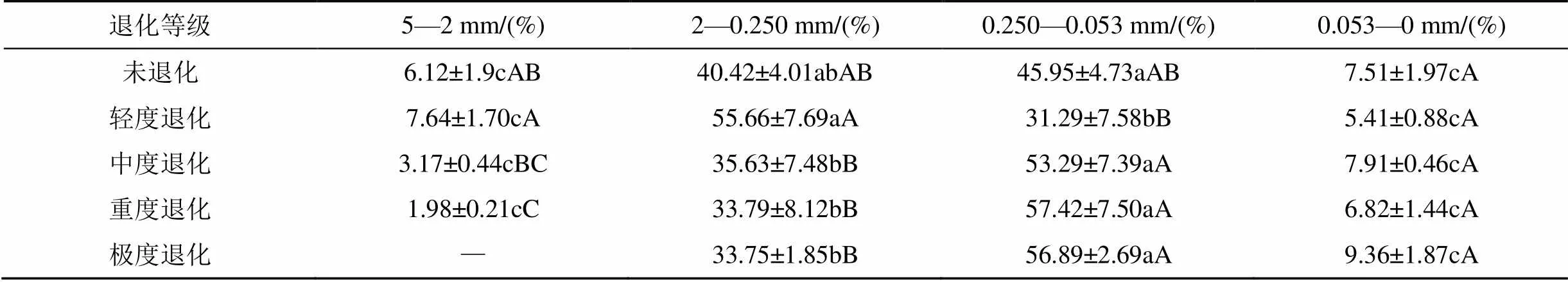
表2 不同退化沙化阶段土壤团聚体百分比
不同小写字母表示不同粒级在P < 0.05水平上差异显著, 不同大写字母表示不同沙化程度在P < 0.05水平上差异显著。

表3 不同退化沙化阶段土壤团聚体平均重量直径和几何平均直径
不同小写字母表示在P < 0.05水平上差异显著。
从表3中可以看出, MWD和GWD变化趋势相近, 随沙化程度加深均表现出显著下降的趋势(<0.05)。MWD在中度退化处理阶段开始显著降低(P<0.05), 相比较未退化阶段, 中度、重度和极度退化阶段MWD分别降低了31.53 %、38.09 %及46.14 %。GWD在重度退化阶段出现显著降低变化(<0.05)并在极度退化时达到最低值(0.254±0.012 mm), 相比未退化阶段, 重度、极度退化阶段GWD分别降低了32.18 %和41.2 %。
2.5 土壤微生物生物量碳氮、土壤理化及土壤团聚体指数的相关性分析
结果表明(表4), 土壤全氮除与pH呈极显著负相关(<0.05)外, 与其余指标均呈显著或极显著正相关; 有机质与MWD呈极显著正相关(<0.01), 相关系数为0.567。微生物生物量碳与铵态氮、可溶性有机碳、有机质、全氮均有极显著正相关(<0.01)相关系数分别为0.679、0.881、0.848、0.796; 微生物生物量氮除与pH呈极显著负相关(<0.01)外、与其他各养分指标间均呈显著(或极显著)正相关(<0.01); 从土壤结构相关性而言, 微生物碳、氮变化规律相近, 都与MWD呈显著正相关(<0.05)相关系数分别为0.488、0.538。土壤pH值与各指标间均呈显著或极显著负相关。MWD与GWD之间呈极显著正相关(<0.01), 相关系数为0.947。

表4 微生物生物量碳氮、土壤理化及土壤团聚体指数相关性
**<0.01; *<0.05
3 讨论与结论
3.1 退化沙化过程中土壤化学性质的变化
土壤化学性质反映土壤营养状况, 其变化将直接影响土壤微生物群落及植被的生长发育[21]。草地沙化将导致土壤养分含量的骤降, 引起各养分间的比例失调和土壤贫瘠化, 进而限制植物生长发育[22-23]。研究发现, 随着草地退化沙化程度的加剧, 硝态氮、可溶性全氮及可溶性有机碳均表现出显著下降变化(P<0.05), 这与陈东明等(2016)研究结果相似。这是因为草地退化程度的加剧, 地上生物量急剧减少, 地表植被覆盖率降低, 导致凋落物、地下植物根系及根系分泌物减少, 使得植物氮、碳矿化速率降低, 从而导致土壤中化学养分大量损失[24]。本研究表明, 随着草地退化沙化程度加深, 土壤碱性程度不断升高, 这与孙磊等(2016)研究结果相似, 这是因为草地不断退化沙化过程中, 植被减少, 覆盖率降低, 大量地表裸露, 土壤直接受到强烈的风蚀、太阳辐射等, 土壤含水量减少, 水分蒸发量加大, 土壤中的酸性溶液分解挥发, 少量碳酸钙集聚到土壤表层导致pH上升[3,25]。
有机质是草地土壤质量演变的主要标志和土壤结构的关键影响因子[26-27], 本研究中, 未退化阶段有机质含量显著高于其它4个退化阶段, 这与蔡晓布(2008)研究结果相近。未退化阶段地表植被盖度大, 且草毡层保存完整, 故土保水能力较强, 植被条件良好, 地上地下生物量增多并产生大量凋落物质, 将极有助于有机质转化与积累[8]。相关性结果表明(表4), 全氮与有机质、微生物生物量及土壤团聚体结构间存在极显著正相关关系(P<0.01), 这与王彩虹等(2008)、赵彤等(2013)研究结果相近。退化沙化过程中, 草地表层植被量下降, 地下根系减少, 土地裸露, 直接影响着土壤固氮能力, 导致土壤全氮含量降低; 有机质含量发生改变, 降低对土壤的稳定能力, 进而对土壤团聚体结构产生破坏, 土壤结构的破坏, 而土壤微生物是有机质转化过程中最重要的因素[28], 土壤结构稳定性的降低, 将导致微生物活力下降, 直接影响着有机质及全氮转化[9,29]。
3.2 退化沙化对草地土壤微生物生物量碳、氮含量的影响
土壤微生物量是高寒草地生态系统中一个重要的组成部分, 也是土壤有机质中最活跃、最易变化的部分[30]。其活动力强弱与草原植被类型、覆盖率等密切相关[31-33]。本研究表明, 退化沙化过程中土壤微生物生物量碳、氮均呈显著下降趋势, 这与魏卫东等(2014)的研究结果相似[32], 草地退化沙化过程中, 地表植被量不断减少, 覆盖率严重降低, 大量土壤裸露出现, 引起土壤退化, 使得输入土壤中的有机质减少, 微生物活动所需底物降低, 从而减少其生物量碳、氮含量[34]。研究发现, 土壤微生物生物量碳、氮不仅随着退化程度的加剧而降低, 且与土壤有机质和全氮含量变化呈极显著正相关(P<0.01)(表4), 这与林璐等(2013)研究结果相近[35], 这说明土壤中的养分物质, 将显著制约微生物生物量及活性。草地沙化, 土壤裸露将直接影响其有机质和全氮含量, 直接破坏、改变微生物生活环境[36-37]。高寒草地为土壤微生物提供了大量凋落物, 同时地下根系的发育程度以及不同植物根系的分泌物和衰亡都会直接影响到土壤微生物的能源物质数量和土壤环境[32,,38], 草地沙化程度加剧, 草地生态系统逆向演替, 植被覆盖率急剧下降, 植物种类、凋落物大量减少, 土壤有机质、全氮及有机碳等土壤养分大量损失, 土壤结构遭到严重破坏, 这系列过程使土壤微生物生长和繁殖的能源物质减少, 使微生物生育及活动受到抑制[39-40]。
3.3 退化沙化对土壤团聚体的影响
土壤团聚体是土壤的重要结构单元, 是土壤的重要组成部分, 是土壤中物质和能量转化及代谢的场所, 具有保证和协调土壤中的水、肥、气、热, 影响土壤酶的种类和活性, 维持和稳定土壤疏松熟层等3大重要作用[17,41]研究显示, 草地退化沙化不断加剧, 2—0.250 mm、0.250—0.053 mm粒级团聚体所占百分比始终高于5—2 mm、0.053—0 mm粒级团聚体, 这与U Mardhiah等(2014)研究结果相近[43], 这是由于草地退化沙化改变了地上植被和地下根系量, 引起土壤容重增加和水稳性团聚体数量下降, 这将直接或间接影响土壤颗粒组成, 从而改变土壤空隙的大小及分布[41]。退化沙化过程中仅5—2 mm粒级在重度退化阶段表现出显著下降变化(<0.05), 其余粒级并未发生显著变化, 而不同退化沙化阶段MWD、GMD呈现出显著下降趋势(<0.05), 这表明不同的退化程度对土壤团聚体大小分布状况无显著影响, 但对团聚体的稳定性有明显影响[9]。土壤团聚体MWD和GMD是反映土壤团聚体大小分布状况的常用指标, MWD 和GMD 值越大表示土壤团聚体聚合团聚程度越高, 结构稳定性越好[9]。本研究表明, 随着草地沙化程度的加剧, MWD、GWD均呈现出显著下降趋势(P<0.05), 这与(王洋, 2012)研究结果相似[41], 这是因为草地退化沙化致使土壤微生物数量、活力降低, 有机质及植物根系分泌物大量减少, 降低了土壤微小颗粒自身团聚, 直接影响大粒级团聚体形成, 并抑制了土壤结构发展, 降低了土壤的恢复能力[42-44]。同样, 相关性分析显示, 土壤团聚体MWD与微生物量碳、硝态氮、可溶性有机碳和全氮间呈显著或极显著正相关。试验表明团聚体MWD在中度退化阶段开始出现显著下降变化, 而微生物量碳、硝态氮、可溶性有机碳和全氮在轻度退化阶段就开始显著下降, 表明土壤相关碳氮养分的含量变化及其矿化速的改变将优先于土壤结构变化, 并影响着土壤团聚体结构质量的改变[43-44]。
3.4 结论
若尔盖高寒草地退化沙化过程中土壤化学、生物养分总体随草地沙化程度的加深而显著降低, 且在沙化前两阶段其下降百分比最为明显。草地沙化将改变土壤团聚结构, 而这一改变过程与微生物活性、养分转化速率及含量的变化显著相关。试验表明, 沙化初期将是对其进行恢复防治的关键时期, 草地管理过程中, 应加大对于这一阶段的预警工作。在治理与防治草地沙化过程中, 应兼顾土壤结构修复与养分输入, 处理好土壤结构稳定与养分转化间的交互关系。
[1] 税伟, 白剑平, 简小枚, 等. 若尔盖沙化草地恢复过程中土壤特性及水源涵养功能[J]. 生态学报, 2017, 37(1): 277–285.
[2] 邓东周, 杨执衡, 陈洪, 等. 青藏高原东南缘高寒区土地沙化现状及驱动因子分析[J]. 西南林业大学学报, 2011,31(5): 27–32.
[3] 孙磊, 刘玉, 武高林, 魏学红. 藏北退化草地群落生物量与土壤养分的关系[J]. 草业科学, 2016, 33(6): 1062– 1026.
[4] 张继义, 赵哈林. 退化沙质草地恢复过程土壤颗粒组成变化对土壤–植被系统稳定性的影响[J]. 生态环境学报, 2009, 18(4): 1395–1401.
[5] 干友民, 李志丹, 泽柏, 等. 川西北亚高山草地不同退化梯度草地土壤养分变化[J]. 草业学报, 2005, 14(2): 32–38.
[6] 舒向阳,, 胡玉福,, 蒋双龙, 等. 川西北沙化草地植被群落、土壤有机碳及微生物特征[J]. 草业学报, 2016, 25(4): 45–54.
[7] 仲波, 孙庚, 陈冬明, 等. 不同恢复措施对若尔盖沙化退化草地恢复过程中土壤微生物生物量碳氮及土壤酶的影响[J]. 生态环境学报, 2017, 26(3): 392–399.
[8] 蔡晓布, 张永青, 邵伟. 不同退化程度高寒草原土壤肥力变化特征[J]. 生态学报, 2008, 28(3): 1034–1044.
[9] 郑子成, 李廷轩, 张锡洲, 等. 不同土地利用方式下土壤团聚体的组成及稳定性研究[J]. 水土保持学报, 2009, 23(5): 229–236.
[10] 任继周. 草业科学研究方法[M]. 北京: 中国农业出版社, 1998.
[11] ZHENG Zhaopei, LIU Zuoxin. Soil quality and its evaluation[J]. Chinese Journal of Applied Ecology, 2003, 14(1): 131–134
[12] 冯瑞章, 周万海, 龙瑞军, 等. 江河源区不同退化程度高寒草地土壤物理、化学及生物学特征研究[J]. 土壤通报, 2010, 41(2): 263–269.
[13] 梁存柱, 祝延绒, 王德利. 21世纪我国草地生态学研究展望[J]. 应用生态学报, 2002, 13(6): 743 – 746.
[14] 张韫, 郭亚芬, 崔晓阳. 土壤学实验实习指导[M]. 哈尔滨: 东北林业大学出版社, 2011.
[15] PANG Xueyong, BAO Weikai. Effect of Substituting Plantation Species for Native Shrubs on the Water–Holding Characteristics of the Forest Floor on the Eastern Tibetan Plateau[J]. Journal of Resources and Ecology, 2011, 2(3): 217–224.
[16] 吴才武, 夏建新, 段峥嵘. 土壤有机质测定方法述评与展望[J]. 土壤, 2015, 47(3): 453–460.
[17] 王清奎, 汪思龙. 土壤团聚体形成与稳定机制及影响因素[J]. 土壤通报, 2005 , 36(3): 415–421.
[18] 中国科学院南京土壤研究所. 土壤理化分析[M] . 上海: 上海科技出版社, 1978.
[19] SMITH A P, MARIN E, MAD G, et al. Microbial community structure varies across soil organic matter aggregate pools during tropical land cover change[J]. Soil Biology & Biochemistry, 2014, 77(7): 292–303.
[20] 邱莉萍, 张兴昌, 张晋爱. 黄土高原长期培肥土壤团聚体中养分和酶的分布[J]. 生态学报, 2006, 26(2): 364–372.
[21] 杨梅焕, 曹明明, 朱志梅, 等. 毛乌素沙地东南缘沙漠化过程中土壤理化性质分析[J]. 水土保持通报, 2010, 30(2): 169–172.
[22] 聂浩刚, 岳乐平, 杨文, 等. 呼伦贝尔草原沙漠化现状、发展态势与成因分析[J]. 中国沙漠, 2005, 25(5): 635– 639.
[23] ZHAO Halin, YI Xiaoyong, ZHOU Ruilian, et al. Wind erosion and sand accumulation effects on soil properties in Horqin Sandy Farmland, Inner Mongolia[J]. Catena, 2006, 65(1): 71–79.
[24] 陈冬明, 张楠楠, 刘琳, 等. 不同恢复措施对若尔盖沙化草地的恢复效果比较[J]. 应用与环境生物学报, 2016(4): 573–578..
[25] 裴海昆. 不同放牧强度对土壤养分及质地的影响[J]. 青海大学学报, 2004, 22(4): 29–31.
[26] 高英志, 韩兴国, 汪诗平. 放牧对草原土壤的影响[J]. 生态学报, 2004, 24(4): 790–797.
[27] CIHACEK L J, SWAN J B. Effects of erosion on soil chemical properties in the north central region of the United States[J]. Journal of Soil & Water Conservation, 1994, 49(3): 259–265.
[28] 王彩虹, 朱进忠, 范燕敏, 等. 不同退化阶段伊犁绢蒿荒漠草地土壤养分动态[J]. 草业科学, 2008, 25(2): 16–20.
[29] 赵彤, 闫浩, 蒋跃利, 等. 黄土丘陵区植被类型对土壤微生物量碳氮磷的影响[J]. 生态学报, 2013, 33(18): 5615– 5622.
[30] 李浩. 内蒙古荒漠草原土壤微生物季节动态变化的研究[D]. 呼和浩特: 内蒙古师范大学, 2008.
[31] 曾智科. 三江源区高寒草甸土壤微生物季节动态及对草地退化的响应[D]. 西宁: 青海师范大学, 2009.
[32] 魏卫东, 刘育红. 不同退化程度高寒草地土壤微生物量碳特征分析[J]. 西北农业学报, 2014, 23(2): 205–210.
[33] JAV VEEN, LADD JN, FRISSEL M J. Modelling C and N turnover through the microbial biomass in soil[M]. Biological Processes and Soil Fertility. Springer Netherlands, 1984: 257–274.
[34] 林璐, 乌云娜, 田村宪司, 等. 呼伦贝尔典型退化草原土壤理化与微生物性状[J]. 应用生态学报, 2013, 24(12): 3407–3414.
[35] 彭岳林, 蔡晓布, 薛会英. 退化高寒草原土壤微生物变化特性研究[J]. 西北农业学报, 2007, 16(4): 112–115.
[36] 黄德清, 于兰, 张耀生, 等. 祁连山北坡天然草地地下生物量及其与环境因子的关系[J]. 草业学报, 2011, 20(5): 1–10.
[37] 卢虎, 姚拓, 李建宏,等. 高寒地区不同退化草地植被和土壤微生物特性及其相关性研究[J]. 草业学报, 2015, 24(5): 34–43.
[38] 温军, 周华坤, 姚步青, 等. 三江源区不同退化程度高寒草原土壤呼吸特征[J]. 植物生态学报, 2014, 38(2): 209–218.
[39] 蒋永梅, 师尚礼, 田永亮, 等. 高寒草地不同退化程度下土壤微生物及土壤酶活性变化特征[J]. 水土保持学报, 2017, 31(3): 244–249.
[40] SIX J, ELLIOTT E T, PAUSTIAN K. Soil structure and soil organic matter: II. A normalized stability index and the effect of mineralogy[J]. Soil Science Society of America Journal, 2000, 64(3): 1042–1049
[41] 王洋. 不同退化程度下高寒草甸土壤有机碳及团聚体特征研究[D]. 南京: 南京农业大学, 2012.
[42] 陈小红, 段争虎, 谭明亮, 等. 沙漠化逆转过程中土壤颗粒分布及其养分含量的变化特征–以宁夏盐池县为例[J]. 土壤通报, 2010, 27(6): 1412–1417.
[43] MARDHIAH U, CARUSO T, GURMELL A ,et al . Just a matter of time: Fungi and roots significantly and rapidly aggregate soil over four decades along the Tagliamento River, NE Italy[J]. Soil Biology & Biochemistry, 2014, 75: 133–142.
[44] 陈帅, 孙涛. 松嫩草地不同退化阶段的土壤团聚体稳定性[J]. 草业科学, 2017, 34(2): 217–223.
Change of soil nutrient and aggregate structure during the desertification process of grassland in Zoige
CHEN Qiujie1,2, ZHANG Nannan2, ZHONG Bo2, CHEN Dongming4, SUN Geng2,*, LIU Lin,*
1. Department of Grassland Science, College of Animal Science and Technology, Sichuan Agricultural University, Chengdu 611130, China 2. Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization, Chinese Academy of Sciences, Key Laboratory of Ecological Restoration and Biodiversity Conservation, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China 3. Sichuan Ganzi Tibeta Autonomous Prefecture Institute of Animal Husbandry Science, Kangding, Sichuan 626000, China
Zoige is an important water source for the upper reaches of the Yellow River which locates on the eastern edge of the Qinghai-Tibet Plateau. In recent years, due to overgrazing, rodents and pests and other reasons, the Zoige alpine grassland has been seriously degraded, and some grasslands have completely desertified. Grassland desertification has become a major problem imperiling the ecological security of the Zoige alpine grassland. This study was conducted at the Maixi Township, the main desertification area of Zoige. According to the vegetation coverage rate of the desertified grassland, we divided the grassland to five different desertification stages: non-desertification, mildly desertification, moderately desertification, severely desertification and extremely desertification. Soil nutrient and aggregate structure were measured in these desertified grassland. The results showed that: (1) In the process of desertification, the soil nutrient content decreased significantly (<0.05). (2) The structure of soil aggregates changed significantly during desertification. The average weight diameter (MWD) and geometric mean diameter (GWD) of aggregates decreased significantly as the degree of desertification deepened (<0.05). In terms of the percentages of the aggregate fractions of different particle sizes, only the percentage of 5-2 mm particle size aggregate decreased significantly at the stage of severe desertification. (3) The aggregates MWD had significantly positive relationship with microbial biomass, organic matter, total nitrogen, nitrate nitrogen and organic carbon respectively, but had significantly negative relationship with pH (<0.01). During the desertification process of grassland, the soil nutrient content decreased significantly and the soil aggregate structure was seriously damaged. Therefore, in the process of controlling and recovering desertified grassland, it is necessary to take into account the soil structure restoration and nutrient input and improve the interaction between soil aggregate structural stability and nutrient conversion.
Zoige; grassland desertification; soil nutrients; soil aggrega
10.14108/j.cnki.1008-8873.2019.04.003
S155.7+7
A
1008-8873(2019)04-013-08
2018-07-30;
2018-09-16
国家科技部重点研发项目(2016YFC0501803), 国家自然科学基金项目(31570517, 31500346, 31350110328) , 中国科学院STS(KFJ-STS-ZDTP- 022-4)、西部之光项目, 四川省科技厅国际合作项目(2016HH0087), 四川省科技厅项目(2015HH0025, 2016HH0082, 2017SZ0080, 2017HH0084, 2017NFP0223, 2018NFP0107, 2018HH0008, 2018HH0021, 2018SZ0329); 中国科学院西部之光项目; 中国科学院成都生物研究所青年研究员项目资助
陈秋捷(1993—),男,四川宜宾人,硕士研究生, E-mail:979424456@qq.com
( 孙庚:E-mail:sungeng@cib.ac.cn; 刘琳:liulinsky@126.com)
陈秋捷, 张楠楠, 仲波, 等. 若尔盖高寒草地退化沙化过程中土壤养分与团聚体结构的变化特征[J]. 生态科学, 2019, 38(4): 13-20.
CHEN Qiujie, ZHANG Nannan, ZHONG Bo, et al. Change of soil nutrient and aggregate structure during the desertification process of grassland in Zoige[J]. Ecological Science, 2019, 38(4): 13-20.
猜你喜欢
矿冶工程(2022年6期)2023-01-12 02:15:10
科学技术创新(2021年22期)2021-08-16 11:17:48
成都信息工程大学学报(2021年1期)2021-07-22 07:21:50
绿色中国(2017年7期)2017-08-02 17:47:11
青年文学家(2016年34期)2017-03-31 18:32:36
环境保护与循环经济(2017年11期)2017-03-16 03:17:07
中国诗歌(2016年7期)2016-12-05 20:43:22
绿色中国(2016年1期)2016-06-05 09:03:00
绿色中国(2016年1期)2016-06-05 09:03:00
高原山地气象研究(2016年3期)2016-02-28 13:53:19
