
不同刺激方式下油松针叶蛋白质含量的变化
2019-09-11 08:47谭红岩陈利英赵茹霞周国娜
安徽农学通报 2019年15期
谭红岩 陈利英 赵茹霞 周国娜


摘 要:针对不同林分类型(油松纯林和油松-山杏混交林)和不同刺激(取食和剪叶)下油松针叶蛋白质含量的变化进行研究,结果表明:混交林相比于油松纯林,油松针叶蛋白质含量高;10头油松毛虫处理的油松针叶比对照的蛋白质含量高;30头油松毛虫处理的油松针叶比对照的蛋白质含量低;而剪叶刺激下的油松针叶與对照的蛋白质含量差异不明显。该研究揭示了不同条件对油松针叶蛋白质含量的影响,为进一步探讨油松的防御机制和对油松毛虫的生态防治、有效调控提供理论依据。
关键词:林分类型;取食;剪叶;油松;蛋白质;含量
中图分类号 S79文献标识码 A文章编号 1007-7731(2019)15-0020-04
Changes of Protein Content in Pinus tabuliformis Needles under Different Stimulation Methods
Tan Hongyan1 et al.
(1Pingquan Huangtuliangzi Forestry Adiministration,Pingquan 067500,China)
Abstract:In this paper,the changes in protein content of Pinus tabuliformis needles under different forest types(pure forest of Pinus tabuliformis and Pinus tabuliformis and apricot mixed forest) and different stimulates(feeding and cutting leaves) were studied.The results show that the protein content of Pinus tabuliformis needles in mixed forest was higher than that in pure forest;the protein content of Pinus tabuliformis needles treated with 10 Dendrolimus tabulaeformis was higher than that of contrast;the protein content of Pinus tabuliformis needles treated with 30 Dendrolimus tabulaeformis was lower than that of contrast;however,the protein content of Pinus tabuliformis needles stimulated by leaf cutting was not significantly different from that of contrast.The effects of different conditions on the protein content in Pinus tabuliformis needles were revealed.It provides theoretical basis for further discussion on the defense mechanism of Pinus tabuliformis and ecological control and effective regulation of Dendrolimus tabulaeformis.
Key words:Forest type;Feeding;Leaf cutting;Pinus tabuliformis;Protein;Content
油松是我国特有的树种之一,也是我国北方干旱地区的主要造林树种,生于海拔100~2600m地带,多为纯林。油松根系发达,耐干旱、耐寒冷,具有很强的保持水土和涵养水源的能力[1],其不仅具有重要的生态价值,还有着重要的工业价值和观赏价值。
油松毛虫属鳞翅目枯叶蛾科松毛虫属,发生量大、为害严重,轻则使针叶枯萎、发黄,重则导致油松整株或成片死亡[2-3],对河北省的油松林造成了极大的影响。为了更好地对油松毛虫进行生态防治,有必要研究油松毛虫诱导油松的抗性机制。研究发现,植物在面对化学刺激、机械损伤、昆虫取食、逆境胁迫等不利的生存环境时,为了自我保护,会刺激其自身防御机制的形成。一般来说,诱导防御的过程有以下3个阶段:系统监测、信号识别、防御物质的产生[4]。对于防御物质,有学者发现,在被油松毛虫取食后,油松体内各种营养物质均发生了一定的变化,说明营养物质也可能参与了防御过程[5-6]。植物对昆虫产生防御机制时,最主要的物质是化学物质,如某些次生代谢物、抗性相关蛋白质酶、蛋白质酶抑制剂等[7-8]。取食和剪叶刺激也可以诱导蛋白质酶抑制剂的变化[9]。总的来说,植物激活自身的防御系统主要是通过形成与抗性相关的物质,进而进行防御[10-11]。
本研究通过比较不同林分类型和不同刺激方式下油松针叶内蛋白质含量的差异,为进一步了解油松在不同条件下的抗性机制以及油松和油松毛虫之间的相互作用提供理论基础,对油松毛虫的生态防治、有效调控具有一定的实践指导意义。
1 试验地概况
试验地为油松毛虫的多发区,位于承德市平泉县黄土梁子镇的油松林区。海拔550~950m,属于燕山山脉的东北方向山地的中心地带,是河北省油松适生区的代表地段。
2 材料与方法
2.1 材料采集 分别在油松-山杏混交林和油松纯林2种类型的林地内设置一个50m×50m的标准样地,在标准样地内选择无松毛虫取食、树龄为25a生的油松各20株(注意:每株树的各项条件都保持一致)。分别设置4个处理:对照、10头油松毛虫取食(简称:10头处理,下同)、30头油松毛虫取食(简称:30头处理,下同)、剪叶处理(注意:要保证每天剪掉的量与30头油松毛虫每天的取食量相同)。每处理设置5次重复,选择4~7龄的油松毛虫进行实验,采用野外套笼放养的方法,注意定期更换套笼枝条,经过为期60d的实验后结束处理。分别在各种处理的每株油松大致相同位置的树冠中剪取未被油松毛虫取食过的2a生新鲜的油松针叶,迅速放入液氮中暂时保存,将其带回实验室后放入-80℃超低温冰箱中保存。
2.2 试验方法
2.2.1 仪器设备 分析天平、称量纸、研钵、药匙、水浴锅、移液枪、无菌枪头、10mL离心管、离心机、300mL锥形瓶、烧杯、HY-5A回旋式振荡器(金坛市杰瑞尔电器有限公司)、Eppendorf5430R小型台式高速冷冻离心机、冰袋、试管架。
2.2.2 主要試剂及用量 详见表1。
2.2.3 蛋白质提取 (1)每组样品各1g,放于事先预冷的研钵中研磨,为方便研磨,研磨过程中可加入液氮。将样品研磨成粉末,然后每个研钵中加入3mL预冷的蛋白质提取缓冲液,继续研磨至匀浆状。(2)将研磨好的匀浆转移至10mL离心管中,涡旋30s后,于4℃下静置4h。(3)在离心管中加入等体积的Tris-饱和酚(pH7.5),混合物涡旋30s,于-20℃静置1h。(4)取出并于4℃下10000r/min离心30min,然后用移液器吸取上层酚层,导入10mL离心管中(导入之前称管重),剩余的水相用4mL提取液和4mL Tris-饱和酚(PH7.5)再次提取。(5)离心后再次吸取酚层导入之前的离心管中,将2次提取的酚层合并混匀,再加入5倍体积预冷的0.1M乙酸铵/甲醇使蛋白质沉淀,涡旋后置于-20℃过夜。(6)次日早晨取出混合物,在4℃下10000r/min离心30min,将沉淀取出,用预冷的0.1M乙酸铵/甲醇、甲醇、丙酮、80%丙酮依次清洗后真空干燥。(7)称重,减去之前的空管重,计算所提取蛋白质的含量。
2.3 数据的统计分析 用Excel 2010软件进行数据计算及绘制图表,用DPS软件,LSD法进行多重比较。
3 结果与分析
3.1 不同处理对油松针叶蛋白质含量的影响 由表2可以看出:混交林的不同处理之间,蛋白质含量表现为:10头处理>对照>剪叶处理>30头处理;与对照相比,10头处理的油松针叶内蛋白质含量较多,30头处理的油松针叶内蛋白质含量较少,剪叶处理则无明显差别;经方差分析发现(F=230.38,P=0.0001),各种处理方式均可对油松针叶蛋白质含量产生影响,且达到显著水平,但各处理影响的程度存在差异。不同处理方式之间,昆虫取食对油松针叶蛋白质含量的影响程度更明显,而剪叶刺激对油松针叶蛋白质含量的影响程度则不明显。10头处理比30头处理的油松针叶蛋白质含量明显增多,且达到极显著。纯林的不同处理之间表现出与混交林相同的规律。
以上数据表明:昆虫取食和剪叶刺激均可诱导油松防御机制的产生,并刺激相关蛋白质的合成,但昆虫取食诱导油松产生防御机制的程度更明显,且随着取食程度的增加,相关蛋白质的含量减少。可能是因为油松毛虫的取食刺激是物理机械损伤以及昆虫口腔分泌物的化学刺激的双重诱导,而剪叶处理主要是单纯的机械损伤,前者所造成的影响更加繁琐而复杂[12]。当外界刺激到达一定程度时,才能刺激油松体内的防御机制,产生防御蛋白质。但当刺激超出了某一范围时,比如油松毛虫数量过多,会摧毁油松的防御机制,导致油松针叶内蛋白质减少[13-14]。
3.2 不同林分类型对油松针叶蛋白质含量的影响 通过了解2种不同林分类型的油松针叶蛋白质含量的变化,进一步分析林分类型对油松针叶蛋白质含量变化的影响。
3.2.1 自然条件下(对照处理)不同林分类型油松针叶蛋白质含量的变化 从图1可以看出:混交林的油松针叶蛋白质含量比纯林的高(混交林为0.017,纯林为0.0154);从显著性水平来看(F=73.52,P=0.001),混交林和纯林自然状态下的油松针叶蛋白质含量差异性极其显著。说明自然条件下,不同林分类型的油松针叶蛋白质含量不同,且混交林大于纯林。而造成这种现象的原因可能与不同林分类型下阳光、水分、热量、养分、生物多样性等方面存在的差异有关。另外,有关学者发现同一种植物在不同的林分类型下,植物体内的生化物质发生变化,使蛋白质含量发生变化[15]。
3.2.2 10头取食处理油松针叶蛋白质含量的变化 从图2可以看出:混交林10头取食的油松针叶蛋白质含量比纯林的高(混交林为0.0206,纯林为0.0185);从显著性水平来看(F=48.65,P=0.0022),两者之间的差异达到极显著水平。说明10头油松毛虫取食刺激下,不同林分类型的油松表现的反应不同,且混交林的油松明显高于纯林。可能是因为混交林中群落结构较复杂,物种间竞争激烈,为适应这种较为复杂的环境,植物的防御能力比纯林中植物的防御能力较强,导致防御蛋白质过量表达。
3.2.3 30头取食处理油松针叶蛋白质含量的变化 从图3可以看出:混交林30头取食的油松针叶蛋白质含量比纯林的高(混交林为0.0144,纯林为0.0133);从显著性水平来看(F=54.45,P=0.0018),两者之间的差异达到极显著水平。说明经过30头取食的不同林分类型的油松针叶蛋白质含量存在明显差异,变化规律与10头相一致。可见,在一定油松毛虫取食为害下,混交林与纯林相比,表现出较高的防御能力。
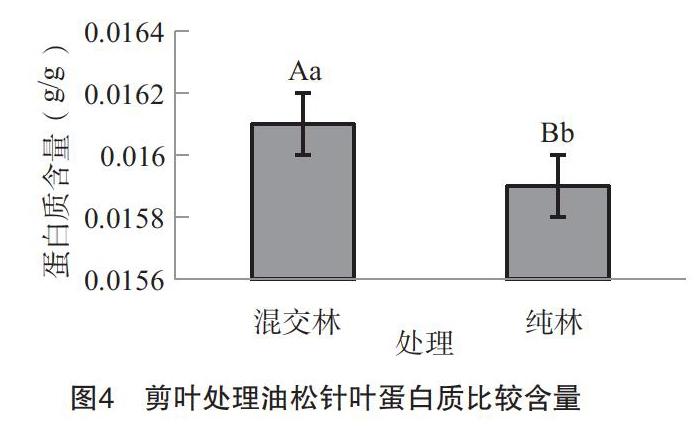
3.2.4 剪叶处理油松针叶蛋白质含量的变化 由图4可以看出:混交林剪叶处理的油松针叶蛋白质含量比纯林处理的稍高(混交林为0.0161,纯林为0.0159),但从显著性水平分析来看(F=7.00,P=0.0572),两者之间的差异并未达到显著性水平。说明剪叶刺激下能引起不同林分类型的油松针叶蛋白质含量的差异,但差异不显著。研究表明,植物抗性不仅表现在植物体内营养物质和某些次生物质的差异上,还表现在植物体内防御蛋白质的变化上。而剪叶处理可能只影响植物体内某些特定的物质变化,而对防御蛋白质方面的影响较小,另外也说明不同的刺激方式下可能诱导的植物体内的抗性机制存在一定的差异。
4 结论与讨论
4.1 讨论 (1)本实验研究发现,油松毛虫取食对油松针叶蛋白质含量存在一定的影响,且10头取食比对照蛋白质含量高,30头取食比对照油松针叶蛋白质含量低,说明油松毛虫数量超过一定限度时,会使体内蛋白质含量减少。也有研究发现,油松体内蛋白质含量与油松的树龄存在一定的关系。至于蛋白含量与油松抗性的关系,还有待于进一步研究。(2)研究表明,植物的抗性随着环境条件和植物体内营养成分而变化,使昆虫的生长发育和繁殖受到影响[15-16],植物体内防御蛋白也随着油松毛虫数量而发生变化。植物体内营养成分以及防御蛋白的变化与昆虫生长繁殖之间的关系,还有待今后作进一步研究。也有研究表明,油松和油松毛虫之间的防御和反防御交替进行。对油松和油松毛虫之间的相互作用进行研究,可以为油松毛虫的生态防治提供理论依据。
4.2 结论 (1)昆虫取食和剪叶刺激均可诱导油松防御机制的产生,并刺激相关蛋白质的合成,但昆虫取食诱导油松合成相关蛋白质的程度更明显。(2)取食程度影響诱导油松产生抗性的程度,在一定的程度内,随着油松毛虫取食程度的增加,油松针叶内相关蛋白质的含量减少。(3)不同林分类型下油松的抗性机制不同,油松针叶内蛋白质含量表现为混交林高于纯林。
参考文献
[1]赵素风.浅议油松的育苗技术[J].应用技术,2012,225:51-52.
[2]刘素华.油松毛虫的危害及防治方法[J].科技情报开发与经济,2006,16(21):285-286.
[3]王世忠,林阳,王鹏,等.朝阳地区油松毛虫的重度危害林分近自然恢复关键技术研究[J].林业科技,2017,42(2):42-46.
[4]陈明顺,仵均祥,张国辉.植物诱导性直接防御[J].应用昆虫学报,2009,46(2):175-186.
[5]王银翠,周国娜,张斌,等.油松毛虫取食和剪叶刺激胁迫下油松的蛋白质表达差异分析[J].林业科学,2016,52(8):1001-7488.
[6]朱麟,杨振德,赵博光,等.植食性昆虫诱导的植物抗性最新研究进展[J].林业科学,2005,41(1):165-173.
[7]李镇宇,王燕,陈华盛,等.油松对赤松毛虫的诱导化学防御及滞后诱导抗性.林业科学[J].2000,36(1):66-70.
[8]戈峰,李典谟,邱业先,等.松树受害后一些化学物质含量的变化及其对马尾松毛虫种群参数的影响[J].昆虫学报,1997,40(4):337-342.
[9]李水清,张钟宁.松墨天牛取食和人为损伤对马尾松针叶部分化学物质含量的影响[J].昆虫学报,2007,50(2):95-100.
[10]王小菲,高文强,刘建锋,等.植物防御策略及其环境驱动机制[J].生态学杂志,2015,34(12):3542-3552.
[11]王琪,严善春.剪叶及昆虫取食对兴安落叶松蛋白质酶抑制剂的影响[J].昆虫学报,2008,51(8):798-803.
[12]殷海娣,黄翠虹,薛堃,等.昆虫唾液成分在昆虫与植物关系中的作用[J].昆虫学报,2006,49(5):843-849.
[13]娄永根,程家安.植物的诱导抗虫性[J].昆虫学报,1997,40(3):320-331.
[14]Wang Y,Ge F,Li ZY.Spatial-tempetial trends of in-duced chemical change in pine pinus massoniana[J].Acta Ecologica Sinica,2001,21(8):1256-1261.
[15]阎凤鸣.化学生态学[M].北京:科学出版社,2003.
[16]理永霞,吕全,梁军,等.杨树接种溃疡病菌后蛋白质表达差异分析[J].南京林业大学学报,2011,35(4):1-6.
(责编:张宏民)
猜你喜欢
肝博士(2022年3期)2022-06-30
氯碱工业(2021年6期)2021-12-25
海外星云(2021年9期)2021-10-14
山西林业(2021年2期)2021-07-21
天津医科大学学报(2021年1期)2021-01-26
世界科学技术-中医药现代化(2020年2期)2020-07-25
现代园艺(2017年22期)2018-01-19
大型铸锻件(2015年5期)2015-12-16
山西农经(2015年7期)2015-07-10
水土保持通报(2014年5期)2014-06-09
