
黑龙江省西部地区引进的饲用燕麦种质资源遗传多样性分析及综合评价
2019-09-10 16:25史京京薛盈文郭伟于崧陆旺于立河胡慧影郭灿侯楠
南方农业学报 2019年3期
史京京 薛盈文 郭伟 于崧 陆旺 于立河 胡慧影 郭灿 侯楠




摘要:【目的】分析黑龙江西部地区引进的饲用燕麦种质资源遗传多样性,为该地区饲用燕麦种质资源创新与新品种选育提供理论参考。【方法】以黑龙江西部地区引种的33份国内外饲用燕麦种质资源为材料,对其14个农艺性状和4个品质性状进行测定,计算遗传多样性指数,并采用主成分分析、相关性分析和聚类分析方法进行综合评价。【结果】33份饲用燕麦种质材料的18个性状存在较大变异,其中鲜草产量的变异系数最大(39.48%),其次为茎干重(35.28%)和叶干重(33.63%),叶可溶性糖含量的变异系数最小(10.97%)。各性状间表现出较好的遗传多样性,遗传多样性指数为1.55~2.02,平均为1.90,其中以籽粒淀粉含量最高(2.02),以千粒重最低(1.55)。各性状间存在不同程度的相关性:株高与穗长、鲜草产量、茎干重和叶干重呈极显著正相关(P<0.01,下同),与籽粒蛋白含量呈显著正相关(P<0.05,下同),与茎可溶性糖含量呈显著负相关;穗长与株高、单株穗铃数、茎干重和叶干重呈极显著正相关,与鲜草产量呈显著正相关。主成分分析發现,前6个主成分的累计贡献率为78.518%,其中,第一主成分(PC1)与饲草产量密切相关,第二主成分(PC2)与籽粒产量密切相关。T07和Ahzzewangc.i3821的综合得分较高,其次是T06、T13和Tihor oats,Heacharrelyngby、白燕7号和引3399的综合得分最低。33份饲用燕麦种质材料可划分为五大类群,其中,Ⅰ类群可作为黑龙江西部地区适宜种植的高产、优质型燕麦材料,Ⅱ类群可用于选育高秆型燕麦,Ⅲ类群可作为黑龙江西部地区选育低矮、抗倒伏、大粒型燕麦性状的良好亲本材料,Ⅳ类群可用作选育分蘖力强、高含糖量型燕麦的良好亲本。【结论】黑龙江西部地区引进的33份国内外饲用燕麦种质资源的遗传多样性丰富,T07、Ahzzewangc.i3821、T06、T13和Tihor oats为适宜黑龙江西部地区种植的饲用燕麦品种。
关键词: 饲用燕麦;种质资源;遗传多样性;综合评价;黑龙江西部地区
中图分类号: S512.602.4 文献标志码: A 文章编号:2095-1191(2019)03-0515-09
0 引言
【研究意义】燕麦(Avena nuda L.)是一种粮饲兼用型作物(Beche,2007;Ren et al.,2007;徐桂花等,2018)。在我国饲用燕麦主要为皮燕麦,具有高产、耐瘠、抗旱和适应性强等特性(周青平等,2015;慕平等,2015;王传凯和郭淼,2017;张志芬等,2017),其营养价值较高,是畜禽的优质食物来源(南铭等,2017)。黑龙江西部地区为半干旱气候区,草场退化、土壤盐渍化和沙化现象日趋加剧,为典型的生态环境脆弱区,也是主要的农牧交错区之一(张晓光等,2013;古丽内尔∙亚森等,2014;Wang et al.,2015),在该地区发展饲用燕麦,对于发展现代畜牧业,调整镰刀湾地区种植业结构具有积极的促进作用。【前人研究进展】我国对于饲用燕麦种质资源的研究多集中于农艺性状和分子标记的遗传多样性分析等。沈国伟等(2010)采用分子标记技术对加拿大和我国的64份燕麦种质资源遗传多样性和群体结构进行分析,结果表明燕麦种质间存在较小的遗传差异。相怀军等(2010)利用AFLP分子标记对国内外177份皮燕麦种质资源进行遗传多样性分析和聚类分析,却发现国内皮燕麦种质遗传多样性较低,与国外材料亲缘关系较远,应加强国外皮燕麦的引种工作。郭红媛等(2014)利用ISSR分子标记对47份燕麦种质资源进行遗传多样性分析,结果表明,燕麦属种间存在丰富的遗传多样性,且燕麦种质资源种内的遗传差异和遗传关系与地理来源密切相关。南铭等(2015)分析了国内外54份燕麦种质资源农艺性状的遗传多样性,结果表明供试种质资源农艺性状间存在较高的遗传多样性。王娟等(2017)对39份燕麦种质资源的15个性状进行形态多样性指数分析,结果表明供试燕麦推广品种的农艺性状间存在较高的形态多样性。武永祯等(2018)评价分析了华北区20个燕麦主栽品种的生产性能和遗传多样性,结果表明其17个质量性状的遗传多样性较低。【本研究切入点】黑龙江西部地区适宜饲用燕麦的开发利用较晚,其遗传多样性分析与综合评价的相关研究至今鲜见报道。【拟解决的关键问题】以黑龙江西部地区引种的33份国内外饲用燕麦种质资源为材料,采用聚类分析、变异性分析和主成分分析方法对其性状和遗传多样性进行分析评价,以期为该地区饲用燕麦的生产及育种实践提供参考。
1 材料与方法
1. 1 试验材料
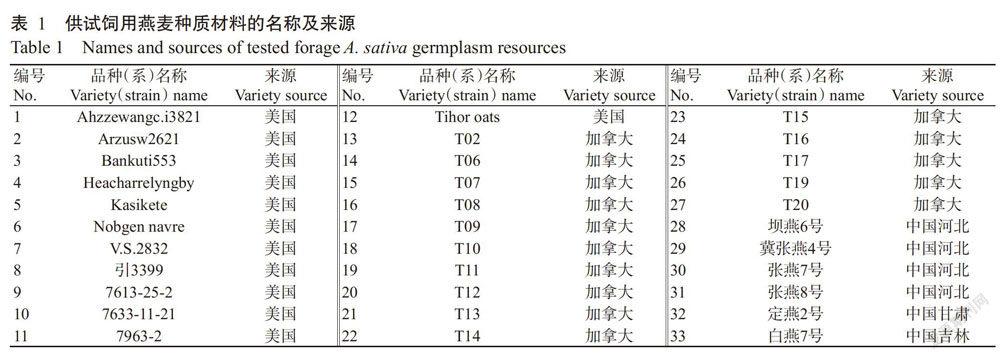
供试材料为黑龙江西部地区从国外引种的27个皮燕麦品种(系)(编号为1~27)和从我国其他省份引种的6个皮燕麦品种(编号为28~33),具体见表1。
1. 2 试验地概况
田间试验于2018年4─8月在黑龙江省大庆市黑龙江八一农垦大学试验实习基地(东经125°20′,北纬46°62′)进行。该区域地处北温带大陆性季风气候区,年平均气温4.2 ℃,年均无霜期143 d,年降水量427.5 mm,年蒸发量1635 mm。土壤类型为盐碱土(pH 8.10),前茬为玉米,土壤基础养分含量:碱解氮114.17 mg/kg、速效磷18.21 mg/kg、速效钾102.47 mg/kg、全氮1.16 g/kg、全磷0.15 g/kg和有机质28.12 g/kg。
1. 3 试验方法
2018年4月22日播种,采用随机区组设计,每品种设4个重复。小区面积为3.0 m×1.8 m,人工开沟条播,行距30 cm,种植密度67万株/ha,一次性施入基肥,施肥量为225 kg/ha(氮∶磷∶钾=1∶1.1∶0.5)。
1. 4 测定指标及方法
对供试材料的农艺性状和品质性状进行测定。其中农艺性状包括分蘖数、株高(基部第一节至顶端小穗长度)、茎粗(基部第一节外径)、收获穗数、穗长、单株穗铃数、单株粒数、单株粒重(带稃)、单株总重(去掉根系后单株地上部分的重量)、千粒重、鲜草产量(去掉根系后植株地上部分的重量)、茎干重、叶干重和籽粒产量。品质性状包括茎可溶性糖含量、叶可溶性糖含量、籽粒淀粉含量和籽粒蛋白含量。
测定方法:于成熟期从各小区随机选取10株进行考种测定,包括分蘖数、株高、茎粗、收获穗数、穗长、单株穗铃数、单株粒数、单株粒重、单株总重和千粒重。且从各小区选取长势均匀的1 m2植株并脱粒,籽粒风干后测定籽粒重量,换算成单位面积产量(kg/ha),即为籽粒产量。于开花期从各小区中齐地面刈割长势均匀的30 cm样段,称重,然后将茎、叶分开烘干再称重,即为鲜草产量、茎干重、叶干重。茎可溶性糖含量(开花期)、叶可溶性糖含量(开花期)和籽粒淀粉含量(成熟期)采用蒽酮比色法(徐仲等,2002)测定。籽粒蛋白含量采用凯氏定氮法(张宪政,1994)测定。
1. 5 统计分析
利用Excel 2003對测定获得的农艺性状数据进行整理分析,并计算各性状的总体平均数(X)、标准差(σ)及不同品种(系)性状的变异系数(CV)。根据计算结果将所有材料划分为10级,第1级[X1<(X-2.0σ)]、第2级[(X-2.0σ)≤X2<(X-1.5σ)]、第3级[(X-1.5σ)≤X3<(X-σ)]、第4级[(X-σ)≤X4<(X-0.5σ)]、第5级[(X-0.5σ)≤X5<X]、第6级[X≤X6<(X+0.5σ)]、第7级[(X+0.5σ)≤X7<(X+σ)]、第8级[(X+σ)≤X8<(X+1.5σ)]、第9级[(X+1.5σ)≤X9<(X+2.0σ)]、第10级[X10≥(X-2.0σ)],每0.5σ为一级。每一级的相对频率Pi用于计算遗传多样性指数(H´),H´=-∑Pi×lnPi,式中,Pi为某性状第i级别内材料份数占总份数的百分比。利用DPS 7.65进行性状相关性分析、主成分分析和聚类分析。
2 结果与分析
2. 1 遗传多样性分析结果
由表2可知,各饲用燕麦种质材料的农艺性状和品质性状存在较大变异,其中鲜草产量的变异系数最大,为39.48%,其次为茎干重和叶干重,分别为35.28%和33.63%,说明引进的饲用燕麦种质对提高、改良黑龙江西部地区的饲用燕麦产量性状具有重要作用;叶可溶性糖含量变异系数最小,为10.97%。33份饲用燕麦种质材料的农艺性状和品质性状遗传多样性指数为1.55~2.02,平均为1.90,其中以籽粒淀粉含量最高(2.02),以千粒重最低(1.55),说明引进的饲用燕麦种质对提高、改良本地区饲用燕麦籽粒品质性状具有重要作用。可见,供试的33份饲用燕麦种质材料的性状存在明显的遗传差异,表现出较好的遗传多样性。
2. 2 性状相关性分析结果
由表3可知,33份饲用燕麦种质材料的各性状间存在不同程度的相关性。株高与穗长、鲜草产量、茎干重和叶干重呈极显著正相关(P<0.01,下同),与籽粒蛋白含量呈显著正相关(P<0.05,下同),与茎可溶性糖含量呈显著负相关,说明通过增加株高可提高饲草产量,改良饲草品质可根据具体需求选择株高。穗长与株高、单株穗铃数、茎干重和叶干重呈极显著正相关,与鲜草产量呈显著正相关,说明增加穗长可提高饲草产量。可见,33份饲用燕麦种质材料的各性状间相互影响、相互制约,在种质创新利用时应综合考虑。
2. 3 主要性状的主成分分析结果
由表4可知,将供试种质材料的18个性状标准化后进行主成分分析,前6个主成分的累计贡献率为78.518%,表明这6个主成分代表了不同性状的大部分信息。第一主成分(PC1)贡献率为25.213%,其中鲜草产量、茎干重和叶干重的特征向量绝对值均大于其他性状,说明PC1主要由鲜草产量、茎干重和叶干重组成,与饲草产量密切相关,PC1的特征值越高,供试材料的饲草产量越高。第二主成分(PC2)贡献率为21.269%,单株穗铃数、单株粒数和单株粒重的特征向量绝对值大于其他性状,说明PC2主要由单株穗铃数、单株粒数和单株粒重组成,与籽粒产量密切相关,PC2的特征值越高,供试材料的籽粒产量越高。第三主成分(PC3)贡献率为9.902%,株高、穗长和千粒重的特征向量绝对值大于其他性状,说明PC3主要由株高、穗长和千粒重组成。第四主成分(PC4)贡献率为8.369%,分蘖数和单株总重的特征向量绝对值大于其他性状,说明PC4主要由分蘖数和单株总重组成。第五主成分(PC5)贡献率为7.933%,籽粒产量和籽粒淀粉含量的特征向量绝对值大于其他性状,说明PC5由籽粒产量和籽粒淀粉含量组成。第六主成分(PC6)贡献率为5.832%,叶可溶性糖含量的特征向量绝对值大于其他性状,说明PC6由叶可溶性糖含量组成。
综上所述,供试种质材料的18个性状中,鲜草产量、茎干重、叶干重、单株穗铃数、单株粒数、单株粒重、株高、穗长、千粒重、分蘖数、籽粒产量、籽粒淀粉含量、单株总重和叶可溶性糖含量为重要性状,其中,鲜草产量、茎干重、叶干重和籽粒产量为重要的构成因素指标,在综合考量品种表现时应着重考察这4个指标。
2. 4 综合评价结果
根据主成分分析结果得出相应的因子关系式:
综合评价函数F=λ1/(λ1+λ2+λ3+λ4+λ5+λ6)F1+λ2/(λ1+λ2+λ3+λ4+λ5+λ6)F2 +λ3/(λ1+λ2+λ3+λ4+ λ5+λ6)F3+λ4/(λ1+λ2+λ3+λ4+λ5+λ6)F4+λ5/(λ1+λ2+λ3+λ4+λ5+λ6)F5+λ6/(λ1+λ2+λ3+λ4+λ5+λ6)F6=0.321F1+0.271F2+0.126F3+0.107F4+0.101F5+0.074F6。其中,λ1、λ2、λ3、λ4、λ5和λ6分别为各主成分对应的特征值,根据综合评价函数,计算各品种的综合得分(表5),给予定量化的描述,得分越高,表明综合表现越好。由表5可知,T07和Ahzzewangc.i3821的综合得分较高,其次是T06、T13和Tihor oats,Heacharrelyngby、白燕7号和引3399的综合得分最低。白燕7号为黑龙江西部地区的主栽品种,引3399的综合得分低于白燕7号,说明引3399不适宜黑龙江西部地区引种。
2. 5 聚类分析结果
采用离差平方和法计算供试饲用燕麦种质材料的18个性状欧氏距离,并进行聚类分析。在欧氏距离为11.18时,33份饲用燕麦种质材料可划分为五大类群,各类群特征见表6。
Ⅰ类群包括12份材料。该类群的性状特征:植株分蘖力较弱,穗短;鲜草产量较高;茎和叶干重较高,籽粒产量最高;饲草品质较好(叶可溶性糖含量最高);籽粒品质最好(籽粒淀粉含量最高、蛋白含量较高),可作为黑龙江西部地区适宜种植的高产、优质型燕麦材料。
Ⅱ类群包括3份材料。该类群的性状特征:植株高大,分蘖力弱,单株总重最低,千粒重最重,其余性状处于中间水平,可用于选育高秆型燕麦。
Ⅲ类群包括3份材料。该类群的性状特征:植株低矮粗壮,分蘖力弱;穗较长,收获穗数、单株铃数、单株粒数和单株粒重最高,且变异系数较低,说明该类群具有较高的籽粒产量潜力;鲜草产量最高,且变异系数较低,具有较高的饲草产量潜力;籽粒蛋白含量最高。可见,该类群可作为黑龙江西部地区选育低矮、抗倒伏、大粒型燕麦性状的良好亲本材料。
Ⅳ类群包括6份材料。该类群的性状特征:植株高大粗壮,分蘖力较强;穗最长,收获穗数较大;茎可溶性糖含量最高,籽粒淀粉含量较高。该类群可用作选育分蘖力强、高含糖量型燕麦的良好亲本。
Ⅴ类群包括9份材料。该类群的性状特征:植株籽粒产量较高且变异系数最低,籽粒品质最差(籽粒淀粉含量和蛋白含量最低),其余性状处于中间水平。
3 讨论
3. 1 饲用燕麦种质资源的遗传多样性分析结果
本研究将变异系数与遗传多样性指数相结合分析国内外33种饲用燕麦种质资源的遗传多样性,结果显示,鲜草产量、茎干重和叶干重的变异系数较大,其次是穗粒数、穗粒重和单株总重的变异系数,株高和叶可溶性糖的变异系数较小,其余性状变异居中,该结果比经验性的直观分类结果更准确可靠。引进种质资源的利用应根据性状差异综合考虑,对于变异系数较大的性状,可通过亲本选配和改善栽培措施等方法使其获得较大程度的提高;对于变异系数中等的性状,通过該方法也可获得适度改善;但对于变异系数较小的性状,通过育种手段获得理想目标性状的难度较大(任欣欣等,2010)。尽管种质材料的有些性状变异可能并不符合目前生产的要求,但其所具有的基因资源可能成为特定育种目标的遗传基础,能为亲本选择提供更大的空间(王林海等,2008),因而有必要对其进行深入研究。
遗传多样性是育种工作的核心和基础,是物种遗传信息的总和,其决定了物种适应环境能力的强弱,同时决定了其利用潜力(卫泽等,2010;胡建斌等,2013;代攀虹等,2016)。饲用燕麦种质资源的遗传多样性研究既可为其科学分类提供理论参考,又为其育种合理选择亲本提供科学依据,还有助于充分了解国内外饲用燕麦种质资源的亲缘关系,对其资源管理、评价和利用具有重要意义(代攀虹等,2016)。本研究中33份饲用燕麦种质材料的18个性状遗传多样性指数为1.55~2.02,平均为1.90,说明这些种质资源的遗传差异程度较高,遗传多样性丰富,特别是籽粒产量、叶可溶性糖含量、株高和籽粒淀粉含量的遗传多样性指数较高,可利用引进的种质资源提高、改良黑龙江西部地区现有的饲用燕麦产量和品质。
3. 2 饲用燕麦种质资源的综合评价结果
本研究的相关性分析结果表明,供试燕麦种质材料的18种性状存在不同程度的相关性,其中单株粒重与收获穗数呈显著负相关,与南铭等(2018)的结论一致。株高与鲜草产量、茎干重和叶干重呈极显著正相关,穗长与株高、茎干重和叶干重呈极显著正相关,说明各性状间存在复杂的关联性。由于农艺性状和品质性状繁多,且彼此间存在不同程度的相关性,给国内外饲用燕麦种质资源的创新和利用造成极大阻碍,为此有必要筛选出重要性状并加以深入研究,以促进其在育种和生产上的应用。
主成分分析法是将作物多个性状转化为较少的几个主成分,可反映85%以上的性状信息,从而获得独立的综合指标,在评价和筛选种质资源时既可综合评价供试品种又能简化程序,具有科学性(武永祯等,2018;张露荷等,2018)。本研究利用主成分分析法将18个性状转化为6个主成分,能代表绝大部分的遗传信息,结果表明,鲜草产量、茎干重、叶干重和籽粒产量为重要的构成因素指标,在综合考量品种表现时应着重考察这4个指标。此外,本研究基于主成分分析结果构建了以6个主成分为参数的综合评价模型,计算各种质材料的综合得分,结果显示,T07、Ahzzewangc.i3821、T06、T13和Tihor oats的综合得分较高,可作为育种亲本用于杂交育种,或可通过单株选育形成新品种,具有很高的利用价值。
3. 3 饲用燕麦种质资源的聚类分析结果
聚类分析反映不同品种(系)的遗传差异,将性状相近的聚为一类。本研究的聚类分析结果表明,当欧氏距离为11.18时,33份供试材料可划分为五大类群,其中,Ⅰ类群可作为黑龙江西部地区适宜种植的高产、优质型燕麦材料;Ⅱ类群可用于选育高秆型燕麦;Ⅲ类群可作为黑龙江西部地区选育低矮、抗倒伏、大粒型燕麦性状的良好亲本材料;Ⅳ类群可用作选育分蘖力强、高含糖量型燕麦的良好亲本。归入同一类的材料间遗传差异较小,但不同类的材料间遗传差异较大,故杂交亲本选配不宜在类群内而宜在类群间选择。且我国育成的饲用燕麦品种并未聚成一类,说明材料间的遗传差异与地理来源无明显相关性,推测聚类分析结果是燕麦种质内部遗传因子和外部环境因子相互作用的结果。这与王鸣刚等(2004)、齐冰洁(2009)、张向前等(2010)的研究结果相似。
本研究性状测定的取样量较大,可提高结果分析的准确性,且供试的33个饲用燕麦种质资源均可在黑龙江西部地区正常生长发育,证明该地适宜种植饲用燕麦,表明本研究能获得较全面、稳定的性状数据。基于本研究结果,根据育种目标可选择性状适宜的品种(系),使黑龙江西部地区的燕麦育种更加科学,还可根据供试饲用燕麦种质材料的遗传多样性,利用分子标记分析基因间的遗传规律,从中发掘有利基因,并与常规育种相结合,为燕麦农艺性状的遗传改良提供更加全面的理论依据。
4 结论
黑龙江西部地区引进的33份国内外饲用燕麦种质资源的遗传多样性丰富,T07、Ahzzewangc.i3821、T06、T13和Tihor oats为适宜黑龙江西部地区种植的饲用燕麦品种。
参考文献:
代攀虹,孙君灵,何守朴,王立如,贾银华,潘兆娥,庞保印,杜雄明,王谧. 2016. 陆地棉核心种质表型性状遗传多样性分析及综合评价[J]. 中国农业科学,49(19):3694-3708. [Dai P H,Sun J L,He S P,Wang L R,Jia Y H,Pan Z E,Pang B Y,Du X M,Wang M. 2016. Comprehensive eva-luation and genetic diversity analysis of phenotypic traits of core collection in upland cotton[J]. Scientia Agricultura Sinica,49(19):3694-3708.]
古丽内尔·亚森,杨瑞瑞,曾幼玲. 2014. 混合盐碱胁迫对灰绿藜(Chenopodium glaucum L.)种子萌发的影响[J]. 生态学杂志,33(1):76-82. [Gulineier Y S,Yang R R,Zeng Y L. 2014. Interactive effects of salt-alkali mixed stresses on Chenopodium glaucum L. seed germination[J]. Chinese Journal of Ecology,33(1):76-82.]
郭红媛,贾举庆,吕晋慧,翟志文,靳艳婷,杨武德. 2014. 燕麦属种质资源遗传多样性及遗传演化关系ISSR分析[J]. 草地学报,22(2):344-351. [ Guo H Y,Jia J Q,Lü J H,Zhai Z W,Jin Y T,Yang W D. 2014. Analyses of genetic diversities and evolutions of Avena L. germplasm resour-ces by ISSR[J]. Acta Agrestia Sinica,22(2):344-351.]
胡建斌,馬双武,李建吾,王吉明,李琼,王林忠. 2013. 国外甜瓜种质资源形态性状遗传多样性分析[J]. 植物学报,48(1):42-51. [Hu J B,Ma S W,Li J W,Wang J M,Li Q,Wang L Z. 2013. Genetic diversity of foreign melon(Cucumis melo) germplasm resources by morphological cha-racters[J]. Chinese Bulletin of Botany,48(1):42-51.]
慕平,赵桂琴,柴继宽. 2015. 基于GGE-Biplot的甘肃省不同生态区燕麦生产性能及适应性分析[J]. 中国生态农业学报,23(6):705-712. [Mu P,Zhao G Q,Chai J K. 2015. Analysis of oats productivity and adaptability in different ecological regions of Gansu Province using GGE-Biplot[J]. Chinese Journal of Eco-Agriculture,23(6):705-712.]
南铭,马宁,刘彦明,任生兰,边芳. 2015. 燕麦种质资源农艺性状的遗传多样性分析[J]. 干旱地区农业研究,33(1):262-267. [Nan M,Ma N,Liu Y M,Ren S L,Bian F. 2015. Genetic diversity analysis on agronomic characteri-stics of oat germplasms[J]. Agricultural Research in the Arid Areas,33(1):262-267.]
南铭,赵桂琴,柴继宽. 2017. 黄土高原半干旱区饲用燕麦种质表型性状遗传多样性分析及综合评价[J]. 草地学报,25(6):1197-1205. [Nan M,Zhao G Q,Chai J K. 2017. Phenotypic diversity and comprehensive evaluations of Avena sativa L. germplasm in semi-arid area of the Loess Plateau[J]. Acta Agrestia Sinica,25(6):1197-1205.]
南铭,赵桂琴,李晶,柴继宽. 2018. 西北半干旱区引种燕麦品种产量与品质的关联分析及评价[J]. 草地学报,26(1):125-133. [Nan M,Zhao G Q,Li J,Chai J K. 2018. Co-rrelation analysis and synthesize evaluation of yield and quality introduced oat varieties in the semi-arid of northwest[J]. Acta Agrestia Sinica,26(1):125-133.]
齐冰洁. 2009. 燕麦种质资源遗传多样性研究[D]. 呼和浩特:内蒙古农业大学. [Qi B J. 2009. Study on genetic diversity of germplasm resources of oat(Avena sativa L.)[D]. Hohhot:Inner Mongolia Agricultural University.]
任欣欣,姚占军,岳艳丽,王莉. 2010. 黄淮海麦区四省份小麦品种的农艺性状及遗传多样性分析[J]. 华北农学报,25(1):94-98. [Ren X X,Yao Z J,Yue Y L,Wang L. 2010. Genetic diversity anaiysis of bread wheat(Triticum aestivum L.) cultivars in four provinces of Huang-huai-hai Area beased on agronmic traits[J]. Acta Agriculturae Boreali-Sinica,25(1):94-98.]
沈国伟,李建设,任长忠,胡银岗. 2010. 中加燕麦种质的遗传多样性和群体结构分析[J]. 麦类作物学报,30(4):617-624. [Shen G W,Li J S,Ren C Z,Hu Y G. 2010. Gene-tic diversity and population structure analysis of oat germplasm from China and Canada[J]. Journal of Triticeae Crops,30(4):617-624.]
王传凯,郭淼. 2017. 外源ALA 对常温和高温条件下裸燕麦生长发育及抗氧化酶活性的影响[J]. 河南农业科学,46(7):30-34. [Wang C K,Guo M. 2017. Effect of 5-aminolevulinic acid on growth and antioxidant enzyme acti-vities of Avena nuda under normal and high temperature conditions[J]. Journal of Henan Agricultural Sciences,46(7):30-34.]
王娟,李荫藩,梁秀芝,郑敏娜. 2017. 北方主栽燕麦品种种质资源形态多样性分析[J]. 作物杂志,(4):27-32. [Wang J,Li Y F,Liang X Z,Zheng M N. 2017. Morphological diversity of main oat germplasm resources in northern China[J]. Crops,(4):27-32.]
王林海,王曉伟,詹克慧,马东钦,许兰杰. 2008. 黄淮麦区部分小麦种质资源农艺性状的聚类分析[J]. 中国农学通报,24(4):186-191. [Wang L H,Wang X W,Zhan K H,Ma D Q,Xu L J. 2008. Cluster analysis of some wheat germplasms in Huang-huai area based on agronomic traits[J]. Chinese Agricultural Science Bulletin,24(4):186-191.]
王鸣刚,葛运生,陈亮,杨必键,王小如. 2004. 甘草亲缘关系的RAPD鉴定[J]. 武汉植物学研究,22(4):289-293. [Wang M G,Ge Y S,Chen L,Yang B J,Wang X R. 2004. Relationship analysis of Glycyrrhiza by RAPD[J]. Journal of Wuhan Botanical Research,22(4):289-293.]
卫泽,孙学振,柳宾,张兴居,王芳,马军,刘庆彩,郭海刚,宋宪亮. 2010. 国内外57份棉花种质资源的遗传多样性研究[J]. 山东农业科学,(6):13-18. [Wei Z,Sun X Z,Liu B,Zhang X J,Wang F,Ma J,Liu Q C,Guo H G,Song X L. 2010. Genetic diversity of 57 cotton germplasm resources from China and overseas[J]. Shandong Agricultural Sciences,(6):13-18.]
武永祯,张斌,王霞,葛军勇,李云霞,左文博,董占红,田长叶. 2018. 华北燕麦主栽品种生产性能评价与遗传多样性分析[J]. 西南农业学报,31(3):448-456. [Wu Y Z,Zhang B,Wang X,Ge J Y,Li Y X,Zuo W B,Dong Z H,Tian C Y. 2018. Evaluation on productivity performance and genetic diversity of 20 major oat cultivars in north China[J]. Southwest China Journal of Agricultural Scien-ces,31(3):448-456.]
相怀军,张宗文,吴斌. 2010. 利用AFLP标记分析皮燕麦种质资源遗传多样性[J]. 植物遗传资源学报,11(3):271-277. [Xiang H J,Zhang Z W,Wu B. 2010. Assessment of genetic diversity for hulled oat germplasm using AFLP markers[J]. Journal of Plant Genetic Resources,11(3):271-277.]
徐桂花,于徐根,黄栋,戴征煌,刘水华,胡文婷,甘兴华. 2018. 播种量对冬闲田种植燕麦和饲用豌豆草产量的影响[J]. 江西农业学报,30(9):39-42. [Xu G H,Yu X G,Huang D,Dai Z H,Liu S H,Hu W T,Gan X H. 2018. Effects of seeding rate on hay yield of Avena sativa and Pisum arvense planted in winter idle farmland[J]. Acta Agriculturae Jiangxi,30(9):39-42.]
徐仲,苍晶,郝再斌. 2002. 植物生理实验技术[M]. 哈尔滨:哈尔滨出版社. [Xu Z,Cang J,Hao Z B. 2002. Plant Physio-logical Experiment Technology[M]. Harbin:Harbin Press.]
张露荷,黄华梨,张广忠,季元租,戚建莉,赵晓芳,胡秉芬,朱红斌. 2018. 基于主成分分析法的鲜食枣品种综合评价[J]. 南方农业学报,49(4):727-734. [Zhang L H,Huang H L,Zhang G Z,Ji Y Z,Qi J L,Zhao X F,Hu B F,Zhu H B. 2018. Comprehensive evaluation for fresh-eating jujube varieties based on principal component analysis[J]. Journal of Southern Agriculture,49(4):727-734.]
张宪政. 1994. 作物生理研究法[M]. 北京:农业出版社:150-152. [Zhang X Z. 1994. Crop Physiology Research Method[M]. Beijing:Agricultural Press:150-152.]
张向前,刘景辉,齐冰洁,郭晓霞,焦伟红. 2010. 燕麦种质资源主要农艺性状的遗传多样性分析[J]. 植物遗传资源学报,11(2):168-174. [Zhang X Q,Liu J H,Qi B J,Guo X X,Jiao W H. 2010. Cluster diversity anaiysis of the main agronomic traits in oat germplasm[J]. Journal of Plant Genetic Resources,11(2):168-174.]
张晓光,黄标,梁正伟,赵永存,孙维侠,胡文友. 2013. 松嫩平原西部土壤盐碱化特征研究[J]. 土壤,45(2):1332-1338. [Zhang X G,Huang B,Liang Z W,Zhao Y C,Sun W X,Hu W Y. 2013. Study on salinization characteristics of surface soil in Western Songnen Plain[J]. Soils,45(2):1332-1338.]
张志芬,刘景辉,付晓峰,赵宝平,李立军,刘俊青,杨海顺. 2017. 干旱胁迫对燕麦叶片气孔和叶肉细胞超微结构的影响[J]. 麦类作物学报,37(9):1216-1223. [Zhang Z F,Liu J H,Fu X F,Zhao B P,Li L J,Liu J Q,Yang H S. 2017. Effect of drought stress on stomata and ultrastructure of mesophyll cells of oat leaf[J]. Journal of Triti-ceae Crops,37(9):1216-1223.]
周青平,颜红波,梁国玲,贾志峰,刘文辉,田莉华,陈有军,陈仕勇. 2015. 不同燕麦品种饲草和籽粒生产性能分析[J]. 草业学报,24(10):120-130. [Zhou Q P,Yan H B,Liang G L,Jia Z F,Liu W H,Tian L H,Chen Y J,Chen S Y. 2015. Analysis of the forage and grain productivity of oat cultivars[J]. Acta Prataculturae Sinica,24(10):120-130.]
Beche. 2007. EST-derived microsaltellites as a rich source of molecular markers for oats[J]. Plant Breeding,126(3):274-278.
Ren C Z,Ma B L,Burrows V,Zhou J,Hu Y G,Guo L,Wei L,Sha L,Deng L. 2007. Evaluation of early mature naked oat varieties as a summer-seeded crop in dryland nor-thern climate regions[J]. Field Crop Reseach,103(3):248-254.
Wang L X,Fang C,Wang K. 2015. Physiological responses of Leymus Chinensis to long-term salt,alkali and mixed salt-alkali stresses[J]. Journal of Plant Nutrition,38(4):526-540.
(責任编辑 陈 燕)
猜你喜欢
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
长江蔬菜·学术版(2016年12期)2017-01-12
吉林农业·下半月(2016年11期)2017-01-09
科学与财富(2016年26期)2016-12-01
医学信息(2016年29期)2016-11-28
大学教育(2016年11期)2016-11-16
企业导报(2016年9期)2016-05-26
