
不同间作模式玉米田天敌节肢动物群落特征
2019-09-10 07:22:44贾永超禹田张晓明李强
南方农业学报 2019年7期
贾永超 禹田 张晓明 李强




摘要:【目的】研究3种不同间作模式玉米田天敌节肢动物群落特征,为生态调控玉米田害虫提供理论基础。【方法】以目测法、网捕法和诱集法等对云南省昭通市鲁甸县3种间作模式玉米田(玉米单作、玉米马铃薯间作、玉米马铃薯向日葵间作)天敌节肢动物群落的组成和结构进行系统调查,采用群落特征指数和主分量分析法对不同间作模式玉米田天敌节肢动物群落特征及其相似性进行分析。【结果】调查区玉米田共发现天敌节肢动物2纲(昆虫纲和蛛形纲) 6目(鞘翅目、双翅目、膜翅目、半翅目、革翅目和蜘蛛目)22科92种2442头,其中玉米单作模式田有2纲4目(鞘翅目和双翅目、膜翅目和蜘蛛目)12科37种780头,玉米马铃薯间作模式田有2纲5目(鞘翅目、双翅目、膜翅目、革翅目和蜘蛛目)17科38种834头,玉米马铃薯向日葵间作模式田有2纲5目(鞘翅目、双翅目、膜翅目、半翅目和蜘蛛目)18科46种828头;天敌群落丰富度指数、多样性指数和均匀度指数均表现为玉米马铃薯向日葵间作田>玉米马铃薯间作田>玉米单作田,而优势度指数和优势集中性指数则表现为玉米单作田>玉米马铃薯间作田>玉米马铃薯向日葵间作田。玉米单作田在苗期与玉米马铃薯间作田在苗期的天敌群落相似度最高,为0.682;玉米单作田在花粒期与玉米马铃薯向日葵间作田在苗期的天敌群落相似度最低,为0.290。在玉米单作田中,瓢虫类、寄生蜂类和长足虻类个体数量共同主导天敌群落变化;在玉米马铃薯间作田中,长足虻类和蜘蛛类个体数量共同主导天敌群落变化;在玉米马铃薯向日葵间作田中,寄生蜂类、步甲类和长足虻类个体数量共同主导天敌群落变化。【结论】3种不同间作模式玉米田中以玉米马铃薯向日葵间作田天敌节肢动物群落种类最丰富、稳定性最强,可在云南昭通地区推广应用。
关键词: 玉米;天敌;节肢动物;群落特征;间作模式;云南
0 引言
【研究意义】玉米(Zea mays L.)是全球重要的粮食作物,同时是全球种植范围最广、产量最高的谷类作物(田聪颖,2014)。截至2017年,我国玉米种植面积达3546万ha,占全国粮食播种面积的31.7%;玉米总产量达2.02 亿t,占全国粮食总产量的32.7%(魏小林,2018)。云南省玉米种植面积约133万ha,在省内粮食作物中居第一位,产量居第二位,但每年因害虫危害造成较大的产量损失,成为部分地区农民脱贫的重要障碍(黄钰淼等,2015)。昭通是云南玉米主产区之一,近年来玉米与马铃薯间作的模式较普遍,部分种植区还出现玉米马铃薯向日葵的间作模式。本研究根据昭通市玉米种植的实际情况,通过研究玉米田不同间作模式对天敌节肢动物群落的影响,为科学制定玉米间作方案,更大程度发挥天敌对害虫的控制作用提供理论基础,对实现玉米田害虫综合防控,提升玉米田生态、经济效益,促进当地群众脱贫增收具有重要意义。【前人研究进展】国外有通过在玉米田中间作林木,利用落叶成功控制玉米田斜纹夜蛾(Spodoptera litura Fabricius)的先例(Tiwari et al.,2005);在南非洲玉米种植中,并改变种植模式,提升了玉米田天敌群落的丰富度、多样性,并分析不同种植模式中天敌控制作用的差异,发现玉米间作田天敌作用好于玉米单作田(Soufbaf et al.,2012;Midega et al.,2014)。田耀加等(2012)研究发现,甜玉米间作不同作物后蜘蛛群落较单作玉米田种类更丰富,更有利于发挥对害虫的控制作用。岳磊等(2014)通过对花椒与玉米、大豆等不同作物间作田天敌群落进行系统调查,结果发现无论是玉米大豆花椒间作模式还是花椒玉米、花椒大豆间作模式,天敌群落的豐富度、稳定性均高于花椒单作田。可胜杰(2015)研究发现,玉米马铃薯间作模式较玉米单作模式天敌群落更丰富,对害虫生态调控的效果更凸显。柴正群等(2016)研究不同复杂生境对玉米田天敌群落的影响,结果表明,在云南普洱地区作物丰富度的提高有助于天敌种群增加,能提升天敌的控害能力。【本研究切入点】目前已有玉米马铃薯间作对玉米田昆虫群落和天敌群落影响的文献报道,但未见玉米马铃薯向日葵间作对天敌群落影响的报道。【拟解决的关键问题】以云南省昭通市鲁甸县农业综合开发项目区玉米示范园为调查区,用目测法、网捕法和诱集法等对不同间作模式玉米田天敌节肢动物群落进行系统调查,掌握不同间作模式下玉米田天敌群落的组成及其变化规律,系统分析不同间作模式对天敌群落的影响,为生态调控玉米害虫提供理论基础。
1 材料与方法
1. 1 试验材料
1. 1. 1 调查材料 玉米品种为云瑞7号,由云南省农业科学院粮食作物研究所选育获得;马铃薯(Solanum tuberosum L.)品种为云薯902,由云南省农业科学院经济作物研究所选育获得;向日葵(Helianthus annuus L.)品种为晋葵9号,由山西金鼎生物种业股份有限公司选育获得。
1. 1. 2 试验区情况 调查区属滇东北冷凉玉米种植带,海拔2158 m,年均降水量960 mm,年均气温11 ℃,无霜期200 d。试验地位于云南省昭通市鲁甸县龙树镇农业综合开发项目区玉米示范园(东经103°47′,北纬27°32′)内,该示范园地势平坦,土质均为营养相对较丰富的岩性土壤。地表有少许禾本科杂草,周围部分为玉米马铃薯间作田,部分为玉米单作田。调查期间,玉米田按常规管理。所选择的间作模式样地类型有:(1)玉米单作田(MM):连片种植面积2.5 ha,播种时间为2016年4月25日,行间距45 cm左右,株间距30 cm左右,在连片种植的区域内共设3块样地,面积分别为0.089、0.096和0.098 ha,样地随机排列,间隔120~160 m。(2)玉米马铃薯间作田(MP):连片种植面积15.0 ha,马铃薯播种时间为2016年4月5日,玉米播种时间为2016年4月25日;2行玉米套种2行马铃薯,玉米行间距40 cm,株间距30 cm,玉米与马铃薯行间距30 cm,马铃薯行间距40 cm,株间距30 cm;在连片种植的区域内共设3块样地,面积分别为0.068、0.077和0.070 ha,样地间随机排列,间隔120~160 m。(3)玉米马铃薯向日葵间作田(MPF):连片种植面积3.2 ha,向日葵播种时间为2016年3月30日,马铃薯播种时间为2016年4月5日,玉米播种时间为2016年4月25日,自左向右依次为马铃薯带、玉米带和向日葵带,其中马铃薯带宽2 m,共6行;玉米带宽1.2 m,共4行;向日葵带宽30 cm,共2行;玉米株间距30 cm,玉米行间距40 cm,玉米与马铃薯行间距30 cm,玉米与向日葵行间距30 cm,马铃薯株间距40 cm,马铃薯行间距40 cm,马铃薯与向日葵行间距30 cm,向日葵株间距30 cm,向日葵行间距30 cm;在连片种植的区域内共设3块样地,面积分别为0.111、0.132和0.120 ha,样地间随机排列,间隔120~160 m。
玉米单作田边缘离玉米马铃薯间作田靠近的边缘直线距离2600 m左右,玉米马铃薯间作田处于玉米单作田和玉米马铃薯向日葵间作田的中间,其边缘距离玉米马铃薯向日葵间作田靠近的边缘直线距离1700 m左右。
1. 1. 3 试验用具 捕虫网:长1.5 m,直径36 cm,由河南云飞科技发展有限公司生产;黄板:25 cm×30 cm,双面特粘黄板,由山东腾盛商贸有限公司生产;黄盘规格:20 cm×17.5 cm×4 cm,PVC材质,由华南农业大学生产;糖醋液:按照酒水糖醋以质量比1∶2∶3∶4配制,酒为当地市场上购买的散装玉米酒,酒精浓度约50%,水为当地自来水,糖为当地购买的固体状红糖,蔗糖含量约90%,醋为海天牌老陈醋;奥林巴斯双目解剖镜:奥林巴斯解剖镜SZ51,普赫光电(上海)科技有限公司。
1. 2 调查方法
于2016年5月7日玉米出苗后开始首次定期定点系统调查,至2016年9月15日玉米收获完毕后调查结束,分别于玉米三叶期(5月7日)、拔节期(5月27日)、小喇叭口期(6月17日)、大喇叭口期(7月5日)、抽雄期(7月20日)、抽丝期(8月2日)、乳熟期(8月17日)、蜡熟期(9月1日)和完熟期(9月15日)各调查一次,周年每块样地共调查9次,每次调查时间为9:00~15:00,遇雨天顺延。根据玉米生育进程将三叶期至拔节期归为苗期,将拔节期至抽丝期归为穗期,将抽丝期至完熟期归为花粒期(马磊磊,2014)。每块样地均同期、同时采用植株观察法、网捕法、糖醋液诱集、黄盘诱集和黄板诱集5种调查方法。植株观察法:在每个样地以棋盘式10点取样进行调查,每个点调查2株玉米,每样地共调查20株,3种模式玉米田每次调查9块样地,共180株,准确记录玉米上天敌节肢动物的种类、数量。网捕法:在行走路线上边走边呈Z字形扫网,来回挥网一次记为1网,每样地调查一次扫网30次。黄板、黄盘和糖醋液诱集法:每块样地放置2块黄板,在玉米行间对角处,将黄板用竹竿固定并悬挂在竹竿上部,黄板的悬挂方向为所有样地均统一朝南北方向,随着玉米植株高度的变化不断调节黄板的高度,悬挂高度始终保持在超过玉米植株顶端5~10 cm的位置;在黄盘中倒入约50 mL立白洗洁精,加自来水至黄盘的2/3处,搅拌均匀,将2个黄盘分别放置在试验样地玉米行中对角、与黄板对称的玉米植株根部附近;在每块样地玉米行最中间位置,悬挂放置糖醋液诱集瓶1个,瓶内糖醋液约占容积的2/3(张晓明等,2015)。
1. 3 标本鉴定
观察植株时直接记录天敌节肢动物种类及数量,对黄板、网捕、黄盘和糖醋液中标本用奥林巴斯双目镜鉴定分类到种,对暂时不能鉴定的标本进行编号后请专家协助鉴定。
1. 4 分析方法
1. 4. 1 群落特征指数 采用下列公式对节肢动物群落特征指数进行分析。
式中,Dma表示丰富度指数,反映物种丰富度与个体总数间的关系;S表示群落中总物种数量;N表示物种总个体数量;pi表示第i种天敌的个体数量占群落总个体数量的百分数;Ni表示第i个种群的个体数量;Nmax表示群落中优势种群的个体数量(柴正群等,2016)。
1. 4. 2 相似性分析 根据杰卡特(Jaccad)相似性系数Cj=j(a+b-j)计算,其中,j为群落A与B的共有种数;a为群落A含有的全部种数,b为群落B含有的全部种数(于景金,2009)。
1. 4. 3 主分量分析 以调查数据为样本,以类群种类为划分依据,将划分后各类群天敌节肢动物个体数量设为变量,计算各因子特征向量与负荷量值(周玉锋,2009)。
1. 4. 4 统计分析 运用SPSS 21.0对试验数据进行统计分析,用Excel 2016制图。
2 结果与分析
2. 1 不同间作模式玉米田天敌节肢动物的组成
对不同间作模式玉米田调查共记录天敌节肢动物2纲(昆虫纲和蛛形纲)6目(鞘翅目、双翅目、膜翅目、半翅目、革翅目和蜘蛛目)22科92种2442头(表1)。其中,玉米单作模式田调查到天敌2纲4目(鞘翅目、双翅目、膜翅目和蜘蛛目)12科37种780头,优势类群依次为步甲类、瓢虫类、长足虻类和蜘蛛类,优势种为中华婪步甲(Harpalus sinicus)、异色瓢虫(Harmonia axyridis)、基黄长足虻(Dolichopus simulator)和白斑狼蛛(Striped soldier);玉米马铃薯间作模式田调查到天敌2纲5目(鞘翅目、双翅目、膜翅目、革翅目和蜘蛛目)17科38种834头,优势类群依次为长足虻类、步甲类和蜘蛛类,优势种为基黄长足虻、丽青步甲(Chlaenius pericallus)、黄腰长足虻(Diaphorus lividus)和白色蟹蛛(Thomisus albus);玉米马铃薯向日葵间作模式田调查到天敌2纲5目(鞘翅目、双翅目、膜翅目、半翅目和蜘蛛目)18科46种828头,优势类群依次为步甲类、瓢虫类、长足虻类和隐翅甲类,优势种为谷婪步甲(Harpalus calceatus)、异色瓢虫、丽长足虻(Sciapus sp.)和冠突眼隱翅虫(Stenus coronatus)。
2. 2 不同间作模式玉米田玉米不同生育进程天敌群落的相似性分析结果
由表2可看出,玉米单作模式田在苗期与玉米马铃薯间作模式田在苗期的天敌物种相似度最高,为0.682;其次为玉米马铃薯向日葵间作模式田玉米在穗期和花粒期,天敌物种相似度为0.643;天敌物种相似度最低的为玉米单作田的花粒期与玉米马铃薯向日葵间作模式田的苗期,相似度为0.290。说明3种不同种植模式玉米田在玉米不同生育进程中的天敌群落相似度存在一定差异。
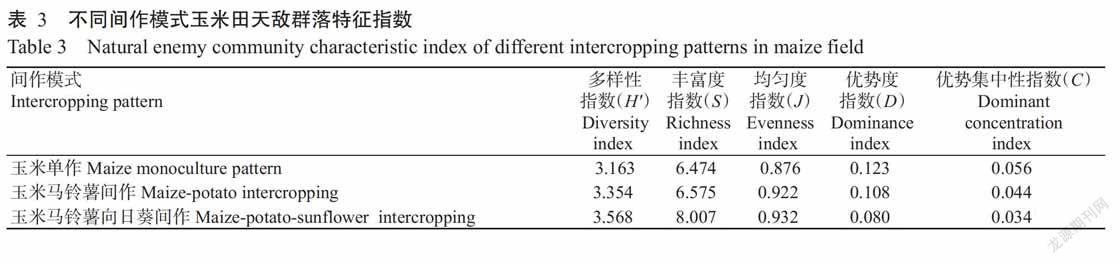
2. 3 不同间作模式玉米田天敌群落特征指数分析结果
由表3可知,3种间作模式中玉米田天敌群落的多样性指数、丰富度指数和均匀度指数均以玉米马铃薯向日葵间作田最高,分别为3.568、8.007和0.932,玉米单作田在3种间作模式中的优势度指数和优势集中性指数最高,分别为0.123和0.056。3种间作模式玉米田天敌群落多样性指数、丰富度指数和均匀度指数均表现为玉米马铃薯向日葵间作田>玉米马铃薯间作田>玉米单作田;而优势度指数和优势集中性指数则恰好相反,表现为玉米单作田>玉米马铃薯间作田>玉米马铃薯向日葵间作田。说明玉米马铃薯向日葵间作模式的天敌群落丰富度最高、稳定性最强。
2. 4 不同间作模式玉米田天敌群落特征的时序动态分析结果
玉米单作田、玉米马铃薯间作田和玉米马铃薯向日葵间作田的天敌多样性指数周年波动分别为1.95~3.14、2.68~3.37和2.94~3.24,天敌群落多样性指数周年平均值分别为2.72、2.95和3.02,玉米马铃薯向日葵间作田天敌群落多样性指数周年变化幅度最小、平均值最高,群落较稳定(图1-A)。玉米单作田、玉米马铃薯间作田和玉米马铃薯向日葵间作田天敌群落丰富度指数周年波动分别为3.46~6.55、4.85~6.35和5.72~7.14,平均值分别为4.99、5.63和6.22,3种间作模式中,玉米马铃薯向日葵间作田的天敌群落丰富度指数年平均值最高、周年波动幅度最小(图1-B)。由图1-C可知,在天敌个体数量上,玉米马铃薯向日葵间作模式田与玉米马铃薯间作模式田周年数量相差较小,均高于玉米单作模式田。由图1-D可知,3种间作模式田中天敌群落周年均匀度指数变化幅度表现为玉米单作田>玉米马铃薯间作田>玉米马铃薯向日葵间作田。从周年群落特征变化来看,玉米马铃薯向日葵间作模式田的稳定性、丰富度和均匀度均最高,与天敌群落特征指数分析结果一致。
2. 5 不同间作模式玉米田天敌群落的主分量分析结果
由表4可知,3种不同模式间作玉米田天敌群落中排在前3个主成分的累积贡献率均在80.00%以上。玉米单作田天敌群落第一主成分(PC1)代表寄生蜂类天敌个体数量、瓢虫类天敌个体数量和长足虻类天敌个体数量的综合因子;第二主成分(PC2)代表蜘蛛类天敌个体数量、寄生蜂类天敌个体数量和步甲类天敌个体数量的综合因子;第三主成分(PC3)代表其他类天敌个体数量和长足虻类天敌个体数量的综合因子。以上3个主成分的累积贡献率总计为83.36%,其中第一主成分累积贡献率最高,为45.61%,说明在昭通玉米单作模式田中,瓢虫类、寄生蜂类和长足虻类个体数量是主导天敌群落变化的主要因子。
玉米马铃薯间作模式田天敌群落的第一主成分代表长足虻类天敌个体数量和蜘蛛类天敌个体数量的综合因子;第二主成分代表其他类天敌和寄生蜂类天敌个体数量的综合因子;第三主成分代表步甲类天敌和瓢虫类天敌个体数量的综合因子。3个主成分的累积贡献率总计为80.87%,其中第一主成分的累积贡献率最高,为40.03%,说明在玉米马铃薯间作模式田中,长足虻类和蜘蛛类个体数量共同主导天敌群落的变化。
玉米马铃薯向日葵间作模式田天敌群落的第一主成分代表寄生蜂类天敌、步甲类和长足虻类天敌个体数量的综合因子;第二主成分代表其他类天敌和瓢虫类天敌个体数量的综合因子;第三主成分代表瓢虫类天敌和蜘蛛类天敌个体数量的综合因子。3个主成分累积贡献率总计为81.38%,其中第一主成分的累积贡献率最高,为38.10%,说明在玉米马铃薯向日葵间作模式田中,寄生蜂类、步甲类和长足虻类3种天敌个体数量共同主导该模式田天敌群落的变化。
综上所述,3种不同间作模式玉米田天敌群落变化的主要影响因子存在差异,玉米单作田中瓢虫类、寄生蜂类和长足虻类个体数量为主导天敌群落变化的主要因子;玉米马铃薯间作田中长足虻类和蜘蛛类个体数量主导天敌群落的变化;玉米马铃薯向日葵间作田中寄生蜂类、步甲类和长足虻类个体数量是主导天敌群落变化的主要因子。
3 讨论
本研究通过对3种不同间作模式玉米田玉米整个生育期的天敌种类及个体数量进行系统调查,发现在天敌物种数量方面表现为玉米马铃薯向日葵间作模式田>玉米马铃薯间作模式田>玉米单作模式田,与田耀加等(2012)不同间作甜玉米对蜘蛛群落的影响结果一致;在个体数量方面,玉米马铃薯向日葵间作模式田与玉米马铃薯间作模式田相差不明显,但明显多于玉米单作模式田。调查发现,玉米马铃薯间作模式田与玉米单作模式田天敌物种数量相近,可能与害虫种类有一定关系。时培建等(2014)研究发现,害虫物种丰富度受到作物物种丰富度的显著影响,但天敌物种丰富度受到的影响相对较小,而天敌物种丰富度受到害虫物种丰富度的影响较明显。说明相邻营养层间有较强的级联效应,而不相邻营养层间的作用关系由于受到中间营养层的影响而相对较弱。不同间作模式玉米田节肢动物群落其级联效应如何尚需进一步研究。
本研究结果表明,玉米单作模式田和玉米马铃薯间作模式田在玉米苗期的天敌群落物种相似度最高,玉米单作模式田在玉米花粒期与玉米马铃薯向日葵间作模式田在玉米苗期天敌群落的物种相似度最低。同一种模式玉米田玉米不同生育进程天敌节肢动物群落相似度也存在差异,可能与玉米不同生育期的环境因子、害虫因子等变化有关(Gao et al.,2008;Fornara and Tilman,2009;Soufbaf et al.,2012;Diehl et al.,2013)。节肢动物群落的多样性指数不仅体现了群落中物种的丰富程度、变异情况及均匀度情况,还是群落组织水平的重要体现,同时能在一定程度上体现当地气候、环境、地理状况及群落的发展情况(曹宁宁等,2019)。从天敌群落多样性指数、丰富度指数和均匀度指数来看,3种不同间作模式玉米田均表现为玉米马铃薯向日葵间作模式田>玉米马铃薯间作模式田>玉米单作模式田,而优势度指数和优势集中性指数均表现为玉米单作模式田>玉米马铃薯间作模式田>玉米马铃薯向日葵间作模式田,说明玉米马铃薯向日葵间作模式中天敌群落的稳定性最强、天敌种类最多,玉米马铃薯间作模式次之,玉米单作模式天敌群落稳定性较差,对3种玉米间作模式天敌群落动态周年变化的研究也证明了这一结果。从增加田间天敌类群及天敌群落稳定性的角度分析,玉米马铃薯向日葵间作模式优于玉米馬铃薯间作模式,两种间作模式均优于玉米单作模式。Wyckhuys和O’Neil(2007)通过增加玉米田种植的复杂程度提升蜘蛛和地甲虫的数量,改变了玉米田天敌群落的结构,本研究结果与其相近。也有学者得出不同的结论,如通过在玉米田中种植植物篱的方式改变玉米田种植模式,发现对玉米田蜘蛛群落的活动密度没有影响(Boutin et al.,2009),究其原因可能是植物篱的存在为蜘蛛提供了更好的休息环境和食物来源,导致蜘蛛不愿由植物篱中进入玉米田(Kromp and Steinberger,1992)。也有研究表明,通过在玉米田周围种植柳枝稷以增加植被种类却出现不利于瓢虫发生的现象(Gardiner et al.,2010)。对于玉米田间作具体的作物种类及布局能否起到对天敌更积极的影响,尚需进一步探究。
3种间作模式玉米田天敌群落变化均受到多个因子主导,且存在一定区别,玉米单作模式中主导因子为瓢虫类、寄生蜂类和长足虻类天敌节肢动物的个体数量;玉米马铃薯间作模式为长足虻类和蜘蛛类的个体数量;玉米马铃薯向日葵间作模式为寄生蜂类、步甲类和长足虻类天敌的个体数量。张晓明等(2009)、蔚立强等(2017)分别研究不同间作模式花椒园和不同杂草比例玉米园昆虫群落变化的主导因子,结果表明不同复杂程度的田间,影响昆虫群落变化的主导因子也有所不同,间作作物的复杂度和杂草比例的不同决定了天敌种类和数量的多少,表现出群落稳定性的变化。然而天敌个体数量、种类、多样性变化对害虫的抑制作用还需进一步研究,如有研究认为增加生境结构上的多样性,会降低狼蛛对猎物捕食的效果,出现天敌丰富度提升但控害能力下降的情况(张永强,1993)。从另一方面来看,也有可能是因为天敌内部产生共争猎物情况而出现种内残杀现象,降低了对害虫的控制作用,同时这些现象能影响天敌物种的分布(Davey et al.,2013; Sitvarin and Rypstra,2014)。对于不同间作模式玉米田中是否存在上述现象仍需深入研究。
4 结论
3种不同间作模式玉米田天敌节肢动物群落组成存在差异,在天敌群落丰富度、多样性和均匀度指数方面均表现为玉米马铃薯向日葵间作模式田最高,天敌种类更丰富,稳定性更强。玉米田间作马铃薯向日葵不仅提高了天敌群落的种群数量,增加了天敌群落的稳定性,同时通过间作能充分利用土地和自然资源,对玉米生产的可持续发展和提高玉米田经济、社会和生态效益及农民收益均具有一定的现实意义,可在云南昭通地区推广应用。
参考文献:
曹宁宁,葛文超,叶辰,武晟兴,杜广祖,陈国华,朱书生,陈斌. 2019. 思茅松林下三七上节肢动物群落组成结构及多样性分析[J]. 南方农业学报,50(2): 299-306. [Cao N N,Ge W C,Ye C,Wu S X,Du G Z,Chen G H,Zhu S S,Chen B. 2019. Composition structure and diversity of arthropod communities on understory planted Panax notoginseng in Pinus kesiya var. langbianensis forest[J]. Journal of Southern Agriculture,50(2): 299-306.]
柴正群,可胜杰,黄吉,黄钰淼,贾永超,陶玫,李强,陈国华. 2016. 不同种植环境夏玉米田节肢动物群落特征及稳定性[J]. 生态学杂志,35(12): 3306-3314.[Chai Z Q,Ke S J,Huang J,Huang Y M,Jia Y C,Tao M,Li Q,Chen G H. 2016. Arthropod community characteristics and stability in summer maize fields with different planting environments[J]. Chinese Journal of Ecology,35(12): 3306-3314.]
黄钰淼,董晨辉,柴正群,可胜杰,黄吉,陶玫,李强,杨雪超,陈国华,马丽. 2015. 云南两个不同生态区玉米蚜种群动态研究[J]. 环境昆虫学报,37(5): 946-954. [Huang Y M,Dong C H,Chai Z Q,Ke S J,Huang J,Tao M,Li Q,Yang X C,Chen G H,Ma L. 2015. The population dynamics of Rhopalosiphum maidis Fitch in two ecological systems of Yunnan Province[J]. Journal of Environmental Entomology,37(5): 946-954.]
可胜杰. 2015. 昭通市不同间作玉米田节肢动物群落特征对比研究[D]. 昆明:云南农业大学. [Ke S J. 2015. Comparative study of arthropod community characteristics in
different intercropped maize fields in Zhaotong[D]. Kun-
ming: Yunnan Agricultural University.]
馬磊磊. 2014. 玉米生育进程对播期及其光温因子的响应[D]. 太谷: 山西农业大学. [Ma L L. 2014. Responses of maize growth process to sowing date and the factors of light and temperature[D]. Taigu: Shanxi Agricultural University]
时培建,惠苍,门兴元,赵紫华,欧阳芳,戈峰,金显仕,曹海锋,Li B. Larry. 2014. 作物多样性对害虫及其天敌多样性的级联效应[J]. 中国科学:生命科学,44(1): 75-84.[Shi P J,Hui C,Men X Y,Zhao Z H,Ouyang F,Ge F,Jin X S,Cao H F,Larry L B. 2014. Cascade effects of crop species richness on the diversity of pest insects and their natural enemies[J]. Scientia Sinica(Vitae),44(1):75-84.]
田聪颖. 2014. 国际玉米市场价格对国内玉米市场价格的传递效应[J]. 农业展望,10(9):31-36. [Tian C Y. 2014. Price pass-through effect of maize from international market to domestic market[J]. Agricultural Outlook,10(9): 31-36.]
田耀加,梁广文,曾玲,陆永跃. 2012. 甜玉米间作不同作物对蜘蛛群落结构的影响[J]. 华南农业大学学报,33(3): 342-345. [Tian Y J,Liang G W,Zeng L,Lu Y Y. 2012. Influence of intercropping sweet maize with different crops on the structure of spider community[J]. Journal of South China Agricultural University,33(3): 342-345.]
蔚立强,陈国华,卢文琴,张晓明,吴智峰,吴俊荣,傅杨. 2017. 玉米与杂草互作田昆虫群落结构及稳定性研究[J]. 西南农业学报,30(5): 1057-1062. [Wei L Q,Chen G H,Lu W Q,Zhang X M,Wu Z F,Wu J R,Fu Y. 2017. Study on structure and stability of insect community in maize interaction with weed field[J]. Southwest China Journal of Agricultural Sciences,30(5): 1057-1062.]
魏小林. 2018. 中国玉米期货市场功能效率研究[D]. 沈阳:辽宁大学. [Wei X L. 2018. Research on the functional e-fficiency of China’s corn futures market[D]. Shenyang: Liaoning University.]
于景金. 2009. 塞罕坝华北落叶松人工林下植物多样性研究[D]. 保定:河北农业大学. [Yu J J. 2009. Study on plant diversity of Larix principis-rupprechii plantation in Saihanba[D]. Baoding: Hebei Agricultural University.]
岳磊,罗凯,马丽,宋家雄,石安宪,高熹,李强. 2014. 花椒马铃薯间作对花椒园节肢动物群落结构的影响[J]. 南方农业学报,45(4): 580-584. [Yue L,Luo K,Ma L,Song J X,Shi A X,Gao X,Li Q. 2014. Effects of Zanthoxylum bungeanum intercropped with potato on arthropod community structure in Zanthoxylum bungeanum garden[J]. Journal of Southern Agriculture,45(4): 580-584.]
张晓明,李强,陈国华,石安宪,宋家雄. 2015. 花椒园昆虫群落结构及时间动态的聚类分析[J]. 应用昆虫学报,52(1): 237-248. [Zhang X M,Li Q,Chen G H,Shi A X,Song J X. 2015. Fuzzy clustering analysis of insect community structure in a Zanthoxylum bungeanum garden[J]. Chinese Journal of Applied Entomology,52(1): 237-248.]
张晓明,李强,陈国华,杨洁,高熹,宋家雄. 2009. 不同种植模式花椒园昆虫群落的结构及稳定性[J]. 应用生态学报,20(8):1986-1991. [Zhang X M,Li Q,Chen G H,Yang J,Gao X,Song J X. 2009. The structure and stability of insect community in different planting patterns of Zanthoxylum bungeanum garden[J]. Chinese Journal of A-pplied Ecology,20(8):1986-1991.]
張永强. 1993. 农田狼蛛亚群落结构的研究[J]. 动物学报,39(2): 139-144. [Zhang Y Q. 1993. Research on the structure of the wolf spider subcommunity in cultivated land[J]. Acta Zoologyica Sinica,39(2): 139-144.]
周玉锋. 2009. 苇状羊茅人工草地节肢动物群落研究[D]. 贵阳: 贵州大学. [Zhou Y F. 2009. Research on arthropod community in tall fescue artificial grassland[J]. Guiyang: Guizhou University.]
Boutin C,Martin P A,Baril A. 2009. Arthropod diversity as affected by agricultural management(organic and conventional farming),plant species,and landscape context[J]. Ecoscience,16(4):492-501.
Davey J S,Vaughan I P,Andrew King R. 2013. Intraguild predation in winter wheat: Prey choice by a common epigeal carabid consuming spiders[J]. Journal of Applied Ecology,50(1): 271-279.
Diehl E,Mader V L,Wolters V,Birkhofer K. 2013. Management intensity and vegetation complexity affect webbuil-ding spiders and their prey[J]. Oecologia,13(2):579-589.
Fornara D A,Tilman D. 2009. Ecological mechanisms associa-ted with the positive diversity-productivity relationship in an N-limited grassland[J]. Ecology,90(2): 408-418.
Gao F,Ge F,Liu X H,Song Y. 2008. Impact of insecticides on the structure and productivity of insect pest and natural enemy communities associated with intercropping in cotton agroecosystems[J]. International Journal of Pest Management,54(2):103-114.
Gardiner M A,Tuell J K,Isaacs R. 2010. Implications of three biofuel crops for beneficial arthropods in agricultu-ral landscapes[J]. BioEnergy Research,3(1): 6-19.
Kromp B,Steinberger K. 1992. Grassy field margins and arthropod diversity:A case study on ground beetles and spiders in eastern Austria(Coleoptera:Carabidae;Arachnida:Aranei,Opiliones)[J]. Agriculture,Ecosystems & Environment,40(1):71-93.
Midega C A O,Jonsson M,Khan Z R,Ekbom B 2014. E-ffects of landscape complexity and habitat management on stemborer colonization,parasitism and damage to maize[J]. Agriculture,Ecosystems & Environment,188: 289-293.
Sitvarin M I,Rypstra A L. 2014. The importance of intraguild predation in predicting emergent multiple predator effects[J]. Ecology,95(10):2936-2945.
Soufbaf M,Fathipour Y,Zalucki M P,Hui C. 2012. Importance of primary metabolites in canola in mediating intera-ctions between a specialist leaf-feeding insect and its specialist solitary endoparasitoid[J]. Arthropod-Plant Interact,6(2): 241-250.
Tiwari M,Singh C P,Goel R. 2005. Effect of intercropping on the population,dynamics of insect pests and yield of mustard[J]. Shashpa,12:106-110.
Wyckhuys K A G,O'Neil R J. 2007. Influence of extra-field characteristics to abundance of key natural enemies of Spodoptera frugiperda Smith(Lepidoptera:Noctuidae) in subsistence maize production[J]. International Journal of Pest Management,53(2): 89-99.
(責任编辑 麻小燕)
猜你喜欢
大自然探索(2024年1期)2024-03-19 19:01:03
云南画报(2021年11期)2022-01-18 03:15:40
基层中医药(2021年4期)2021-07-22 07:15:32
科学大众(中学)(2021年3期)2021-05-17 17:07:01
云南画报(2020年12期)2021-01-18 07:19:20
云南画报(2020年9期)2020-10-27 02:03:16
——管氏肿腿蜂的应用技术
江苏林业科技(2020年3期)2020-08-31 01:00:08
非公有制企业党建(2019年7期)2019-07-19 03:31:24
课外生活(小学1-3年级)(2018年10期)2018-11-19 07:26:10
现代园艺(2017年21期)2018-01-03 06:42:15
