
贵州特有植物红花瘤果茶种群空间格局及数量动态研究
2019-09-10 07:22郭米香杨乃坤刘海燕汤升虎范志伟邹天才
广西植物 2019年10期
郭米香 杨乃坤 刘海燕 汤升虎 范志伟 邹天才

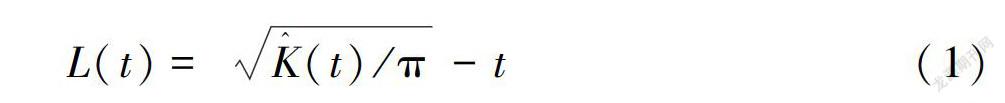





摘 要:紅花瘤果茶(Camellia rubituberculata)为地理分布狭窄的贵州特有植物,颇具经济价值。为查清其分布特征和评估其濒危状况,该文在红花瘤果茶的集中分布区设置三个典型样地进行调查,采用空间点格局及多元点格局分析法分析种群空间分布格局,利用种群动态量化指数分析预测种群数量动态规律。结果表明:(1)兴仁县新寨村大皮坡1号样地幼龄树的小尺度呈集群分布、大尺度呈随机分布,中龄树和老龄树均呈随机分布;兴仁县新寨村槽子土2号样地的幼龄树均呈集群分布,中龄树和老龄树均呈随机分布;晴隆县紫马乡龙头村上捧碧林区3号样地幼龄树、中龄树和老龄树均呈随机分布。(2)不同发育阶段的空间关联主要表现为1号样地幼龄树和中龄树均为负相关,幼龄树和老龄树在中小尺度上为负相关,在大尺度上为无相关,中龄树和老龄树均为无相关。2号样地幼龄树和中龄树在小尺度上为负相关,中大尺度上为无相关;幼龄树和老龄树在中小尺度上为负相关,中大尺度上为无相关;中龄树和老龄树均为无相关。3号样地的幼龄树和中龄树均为正相关;幼龄树与老龄树在小尺度上为负相关,中尺度上为无相关,大尺度上为正相关;中龄树和老龄树均为无相关。(3)种群数量动态分析表明3个样地中幼苗补充充裕,种群结构为增长型,但易受外界环境干扰、生态脆弱。(4)红花瘤果茶的生境异质性及其种内种间关系相互作用的结果是影响种群数量扩大和物种散布的主要因素。
关键词: 红花瘤果茶, 地理分布, 空间分布格局, 数量动态, 贵州特有
Spatial distribution pattern and quantitative dynamics of the endemic plant Camellia rubituberculata in Guizhou Province
GUO Mixiang1, YANG Naikun2, LIU Haiyan3, TANG Shenghu3, FAN Zhiwei3, ZOU Tiancai4*
( 1. College of Forestry, Guizhou University, Guiyang 550025, China; 2. Guiyang Zhongxiong Forestry Ecological Engineering Investigationn and Design Co. Ltd., Guiyang 55004, China; 3. Guizhou Botanical Garden, Guiyang 550004, China; 4. Guizhou Academy of Sciences, Guiyang 550001, China )
Abstract:Camellia rubituberculata is endemic to Guizhou Province with narrow geographical distribution and has economic value. In order to find out its distribution characteristics and evaluate its endangered status, three typical sample plots were set up in concentrated areas, and the spatial distribution pattern of population was analyzed by spatial dot pattern and multivariate point pattern analysis, the quantitative index analysis of population dynamics was used to predict the dynamic law of population quantity. The results were as follows: (1) The young trees in Dapipo sample plot 1 showed cluster distribution in small-scale and random distribution in large-scale, and the distribution of middle-aged trees and old-aged trees in all scales were random. The young trees in Caozitu sample plot 2 were clustered at all scales, the middle-aged trees and the old-aged trees were random. The young trees, middle-aged trees and old-aged trees were randomly distributed in Shangpengbi sample plot 3. (2) The spatial correlations of different developmental stages mainly manifested that the young and middle-aged trees in sample plot 1 were negatively correlated at all scales, the young trees and old-aged trees were negatively correlated at medium and small scale, and there were no correlation in the large scale. The middle age trees and the old-aged trees were not related at all scales. In sample plot 2, there was negative correlation between young trees and middle-aged trees at small scale and no correlation at medium and large scales, young trees and old-aged trees were negatively correlated at medium and small scales, but not at the medium and large scales, there was no correlation between middle-aged trees and old-aged trees at all scales. The young trees and middle-aged trees in sample plot 3 were positively correlated at all scales; there was a negative correlation between young trees and old-aged trees on the small scale, but no correlation on medium scale, and positive correlation on the large scale; the middle-aged trees and old-aged trees were irrelevant at all scales. (3) The population quantitative dynamic analysis indicated that the seedlings in the three sample plots were well supplemented, and the population structure was growth type, but they were vulnerable to environmental disturbance and ecological fragility. (4) The results of habitat heterogeneity and interspecific interaction of C. rubituberculata are the main factors affecting population expansion and species dispersal.
Key words: Camellia rubituberculata, geographical distribution, spatial distribution pattern, quantity dynamic, Guizhou endemic
红花瘤果茶又名红花三江瘤果茶(Camellia pyxidiacea var. rubituberculata)(Ming & Bartholomew,2008)、厚壳红瘤果茶(Camellia rubituberculata )(张宏达和任善湘,1998),常绿小乔木,高可达4 m,花大且花期长,果实与种子表面具有瘤状突起,是山茶属植物在系统发育上保持了原始性状的特化物种。主要分布在贵州省黔西南州兴仁县和晴隆县的喀斯特山地,出现在群落中亚乔木层或灌木层,喜光而耐阴,树形优美,为园林景观植物优选,多岩生,适用于石漠化地区造林(邓忠治等,2011)。其分布区极其狭窄,资源稀少,设立自然保护点,在保护的基础上加以研究和开发应用是当务之急(安明态,2005)。当前,已经开展了资源调查(杨冰等,2016)、生物学特性和种子萌发研究(周丽等,2015;郭米香,2018)、生命表及生存力初步研究(袁从军等,2016),但其物种资源的濒危状态评估、种群动态特征、资源保护与利用的科学问题尚需研究。
种群是形成群落的基本单元,也是研究生态学的最小单位(刘贵峰等,2011),种群空间分布格局能反映种群利用环境资源的基本情况,是研究种群个体间相互作用、种群特征及种群与环境关系的重要方法(于倩,2006;钟兆全,2015;许梅等,2017)。研究种群的数量动态及空间分布格局可揭示种群对环境资源的利用与适应,有助于认识种内、种间相互关系及物种的繁殖演替规律(康佳鹏等,2019)。本文以紅花瘤果茶集中分布区的天然群落为研究对象,对其种群空间分布格局和种群数量动态进行实地调查研究,以揭示红花瘤果茶种群空间分布特征及数量动态变化规律,为红花瘤果茶种质资源的有效保护和合理开发利用提供科学依据,也为石漠化地区植被恢复和生态环境建设中特色优势植物的开发利用提供参考。
1 研究区概况
研究区位于贵州省晴隆县紫马乡龙头村和兴仁县田湾乡新寨村,地处喀斯特沙页岩镶嵌的高原峡谷,海拔在1 000~1 500 m之间,岩石裸露达70%~90%,为典型的高原亚热带湿润季风气候,降水丰富,雨热同期(袁从军等,2016)。主要土壤为黑色石灰土,土层厚度差异大(枯落物层约7 cm、A层约12 cm、B层12~27 cm),土被分布不连续,有机质含量高,土质较为疏松(邓忠治等,2011)。实查晴隆县和兴仁县两个分布区的生境概况如表1所示,红花瘤果茶分布为单株或丛生,群落均为常绿落叶阔叶混交林,伴生植物较多,其中乔木层主要有青冈栎(Cyclobalanopsis glauca)、白栎(Quercus fabri)、毛石栎(Lithocarpus petelotii)、云南樟(Cinnamomum glanduliferum)、川黔润楠(Machilus chuanchienensis)等,红花瘤果茶主要位于亚乔木层;灌木层主要有火棘(Pyracantha fortuneana)、清香木(Pistacia weinmanifolia)、小叶女贞(Ligustrum quihoui)等,由于人类从事农林业生产活动的影响,红花瘤果茶也常出现在灌木层,主要伴生植物有大叶酸藤子(Embelia subcoriacea)、杠柳(Periploca sepium)等,草本植物和蕨类植物种类较为稀少。
2 研究方法
2.1 野外样地调查
2018年9月,对红花瘤果茶天然种群分布区及其所处群落设置样地进行了实地调查,在兴仁县新寨村石鹅场组设置2个样地,分别为1号样地大皮坡、2号样地槽子土,在晴隆县紫马乡龙头村上捧碧林区设置3号样地,样地总面积为2 700 m2。采用方精云等(2009)植物群落清查方法分别调查记录每个样地的生境因子(表2)及乔、灌、草各层植物物种组成和每株红花瘤果茶的相对坐标。
2.2 种群的龄级划分
种群的龄级和径级存在一定的差别,但是在相同的环境条件下,同一物种的龄级和径级对环境的响应呈现一致性规律(王磊等,2010;张金屯等,2004)。遵循生物多样性保护原则采用“空间代替时间”的方法(张兴旺等,2013;杨乃坤等;2015),以径级结构代替年龄结构分析其种群动态,根据红花瘤果茶的生长规律及生物学特性,将该种胸径<1 cm划为第一径级,胸径每增加1 cm则增加一个径级, 共划分为11个龄级, 统计各龄级的株数,依次划分为Ⅰ级(DBH<1.0 cm)、Ⅱ级(1.0 cm≤DBH<2.0 cm)、Ⅲ级(2.0 cm≤DBH<3.0cm)、Ⅳ级(3.0 cm≤DBH<4.0 cm)、Ⅴ级(4.0 cm≤DBH<5.0 cm)、Ⅵ级(5.0 cm≤DBH<6.0 cm)、Ⅶ级(6.0 cm≤DBH<7.0 cm)、Ⅷ级(7.0 cm≤DBH<8.0 cm)、Ⅸ级(8.0 cm≤DBH<9.0 cm)、Ⅹ级(9.0 cm≤DBH<10.0 cm)、Ⅺ级(DBH≥10 cm)。根据野外调查情况及红花瘤果茶植株生长特点及袁从军等(2016)对厚壳瘤果茶的生命表分析结果,将Ⅰ龄级的个体(DBH<1.0 cm)划分为幼龄个体,Ⅱ~Ⅴ龄级及胸径为5 cm的个体(1 cm≤DBH<5 cm)划分为中龄个体,Ⅵ~Ⅺ龄级中(DBH≥5 cm)的个体划分为老龄个体,根据以上划分标准统计各龄级的红花瘤果茶植株数量。
2.3 种群空间分布格局分析
采用空间点格局分析法(SPPA,亦称为Ripley’s K系数法)的修正函数L(t)分析红花瘤果茶种群的空间分布格局。
L(t)=K^(t)/π-t(1)
利用Monte-Carlo方法进行一千次随机空间模拟计算出L(t)99%的置信区间。实测L(t)值大于置信区间上限为集群分布,介于置信区间为随机分布,小于置信区间下限时则为均匀分布(张金屯,1998)。
2.4 种群不同发育阶段的空间关联分析
分析两个发育阶段的关系即对其进行多元点格局分析,实际上是两个不同发育阶段的点格局分析(张金屯和孟东平,2004)。计算公式如下:
L12(t)=K^12(t)/π-t(2)
用Monte-Carlo方法检验拟合L12(t),以检验两种间的关联是否显著,调查数据采用生态学软件ADE-4进行分析(李帅峰等,2013)。
2.5 种群动态量化分析
采用种群动态量化方法,推导出红花瘤果茶种群年龄结构的动态指数并描述红花瘤果茶的种群动态特征(陈晓德等,1998;刘海燕等,2016;黄冬柳等,2018)。
3 结果与分析
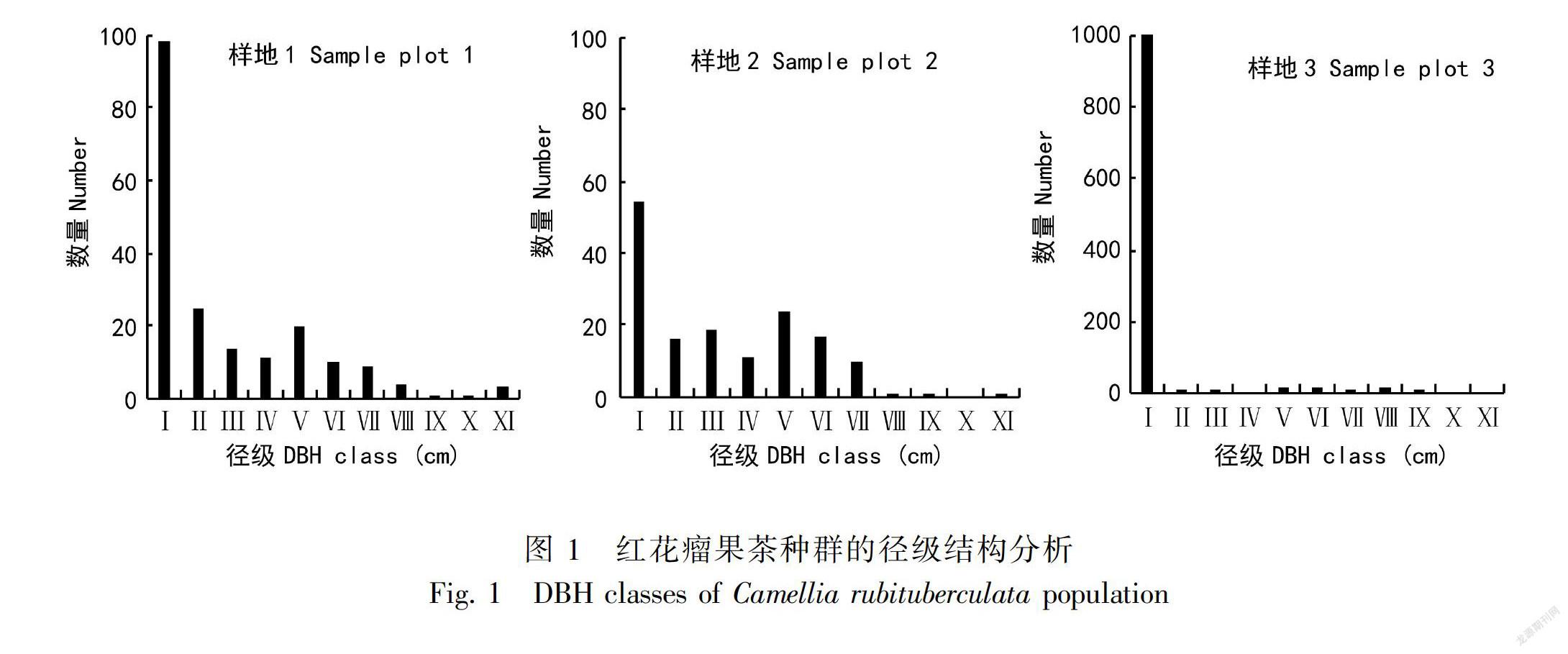
3.1 种群径级分布及年龄结构
调查统计红花瘤果茶种群径级结构如图1所示,3个样地幼龄树最多,中龄和老龄树相对较少。1号样地共有197株红花瘤果茶,幼龄树(DBH<1 cm)98株,占总数的49.75%,中龄树(1 cm≤DBH<5 cm)72株,占总数的36.55%,老龄树(DBH≥5 cm)共27株,占总株数的13.71%。2号样地共154株,幼龄树54株,占35.06%,中龄树72株,占总数的46.75%,老龄树28株,占总数的18.18%。3号样地,存在大量的幼苗个体(约1 000株),占总个体数的93%,中龄树52株,占3.13%,老龄树60株,占3.68%,该种群有大量幼苗储备,具备自然更新能力,属于增长型的种群。3号样地老龄树个体数量约是1号样地和2号样地的两倍,特定时间和空间从母株上散落的种子量丰富,独特的生境导致种子密集分布萌发产生大量的幼苗。
3.2 不同发育阶段的空间点格局分析
红花瘤果茶不同发育阶段个体在种群中分布的调查结果如图2所示,各年龄阶段植株的分布密度呈显著差异。在1号样地中,3个发育阶段的植株均显示出一定程度的集群分布,幼龄树最多,老龄树最少。2号样地中龄树最多,老龄树最少,3个发育阶段均呈不同程度的集群分布,3号样地幼龄树特多,分布最为密集,中龄树和老龄树分布相对均匀。图2虽然在一定程度上呈现了种群的分布类型,但并不能具体反应出该分布类型与样地尺度之间的关系,因此还需对该种群进行空间点格局分析。
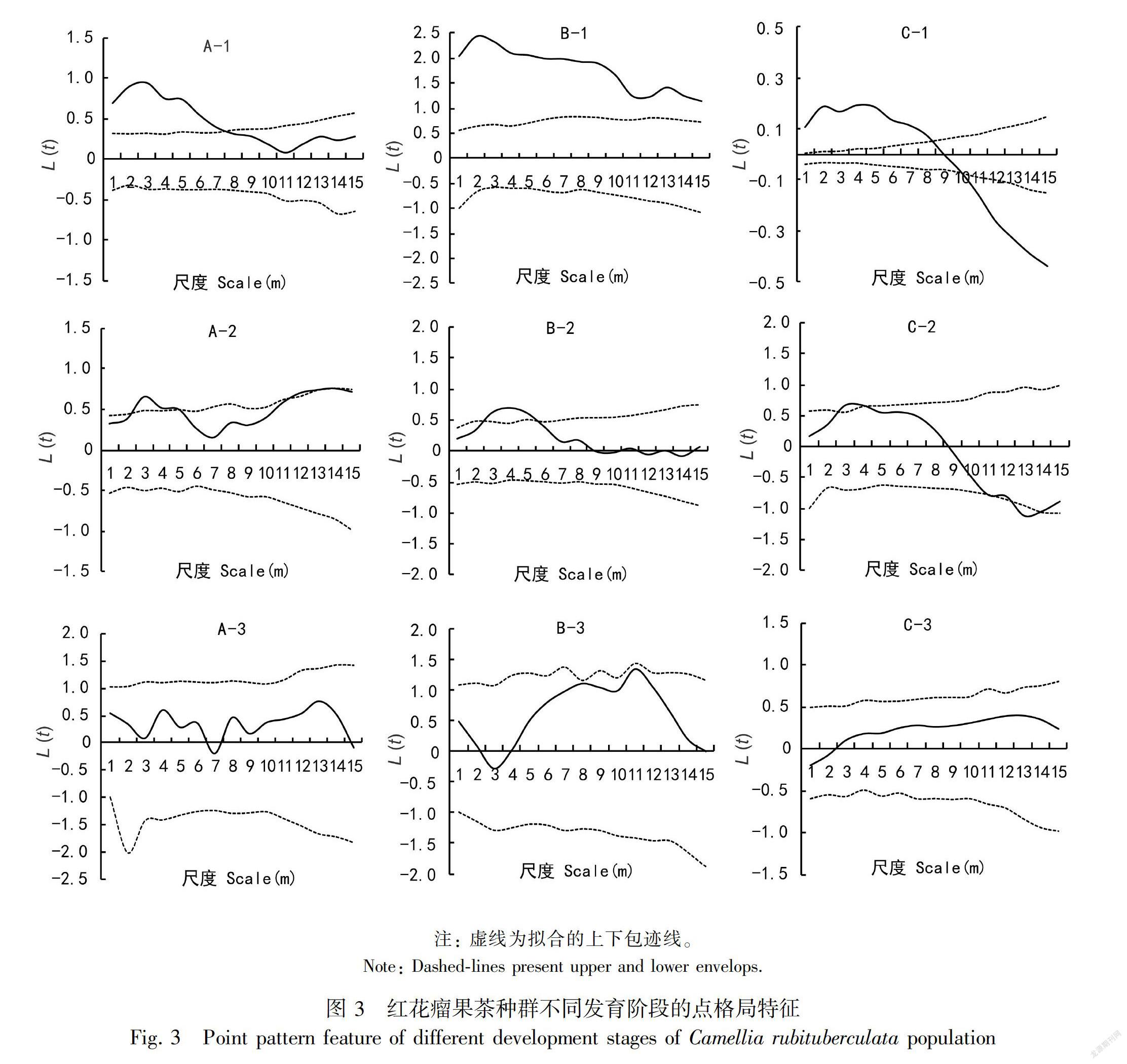
对三个调查样地红花瘤果茶种群不同发育阶段的空间点格局进行了分析,结果如图3所示,实线为实际数据计算的L(t)曲线,虚线为拟合的上下包迹线(置信水平),取t间隔为1 m,t的最大值为样方边长的一半(15 m),x坐标t用空间距离(m)表示。这表明随着樣地尺度的变化,红花瘤果茶不同发育阶段的分布格局也呈现着相应的变化规律。1号样地中幼龄树(A-1)在尺度0~7 m呈现为集群分布,在尺度8~15 m呈现随机分布;中龄树(B-1)所有尺度均呈现为集群分布;老龄树(C-1)在尺度0~8 m为集群分布,在尺度8~9 m为随机分布,在尺度9~15 m为均匀分布。样地2中幼龄树(A-2)在样地尺度0~2 m、尺度4~5 m和尺度10~15 m接近集群分布,但集群分布特征不显著,尺度2~4 m为集群分布,尺度5~10 m为随机分布;中龄树(B-2)在尺度0~2 m和5~15 m为随机分布,在尺度2~5 m为集群分布;老龄树(C-2)在0~2 m、4~10 m、14~15 m尺度上为随机分布,在2~4 m的尺度上呈集群分布,11~14 m的尺度上为均匀分布。3号样地中幼龄树(A-3)在所有尺度均表现为随机分布;中龄树(B-3)在尺度0~7 m、尺度8~10 m、尺度11~15 m均呈现随机分布,在尺度7~8 m和尺度10~11 m接近集群分布;老龄树(C-3)在所有尺度上均为随机分布。
3.3不同发育阶段种群的空间关联分析
对红花瘤果茶种群在不同发育阶段的空间关联性分析结果如图4所示,1号样地中幼龄树与中龄树(A-1)在所有尺度内呈现负相关关系,幼龄树与老龄树(B-1)在尺度0~13 m呈现显著负相关关系,在尺度13~15 m内无相关性,中龄树与老龄树(C-1)在所有尺度内均无相关性。2号样地中幼龄树与中龄树(A-2)在尺度0~4 m呈现负相关关系,在尺度4~15 m内无相关性,幼龄树与老龄树(B-2)在尺度0~9 m呈现负相关关系,在尺度9~15 m无相关性,中龄树与老龄树(C-2)在所有尺度内均无相关性。3号样地中幼龄树与中龄树(A-3)在所有尺度内呈现正相关关系,幼龄树与老龄树(B-3)在尺度0~8 m为显著负相关关系,在尺度8~12 m无相关性,在尺度12~15 m为显著正相关关系,中龄树与老龄树(C-3)在所有尺度内无相关性。
3.4 种群数量动态
红花瘤果茶种群各相邻龄级间的个体数量动态变化分析如表3所示。表3结果表明,3个样地的红花瘤果茶种群发育过程中各个龄级为下一龄级贡献潜力均呈波浪起伏式变化,1号样地Ⅰ~Ⅳ龄级贡献潜力下降趋势明显,Ⅳ龄级为下一级贡献潜力为负值,随后Ⅴ~Ⅷ龄级有所反弹,整体变化呈现“下降-上升-下降”的波浪式起伏,表明1号样地中幼苗充足,能够为种群各生长阶段提供足量的幼苗补给,种群结构为增长型。在2号、3号样地中,各龄级为下一龄级的贡献潜力整体呈“下降-上升-下降-上升-下降”的趋势,起伏频繁,但幼苗补充充足,两个种群结构均呈增长型。3个样地均在Ⅳ龄级时Vx同时出现负值,说明Ⅳ龄级个体死亡率急剧增加,大量个体在这个阶段发生死亡而不能成长为下一龄级个体。
红花瘤果茶种群年龄结构的动态指数(Vpi)的计算结果如表3所示,三个样地的动态指数均大于0,表明红花瘤果茶种群属于稳定增长型种群,能适应当地的气候条件并稳定增长。种群龄级结构的动态指数Vpi′均大于0,又远小于Vpi,表明了红花瘤果茶种群生态脆弱,受外界干扰敏感程度较高。因此,一旦外界环境压力变化加大,例如过度的人为干扰,喀斯特生境异质性的生态恶化,都会影响到红花瘤果茶种群的生长发育。
4 讨论与结论
4.1 红花瘤果茶属增长型种群,中龄树和老龄树大多呈现随机分布
分析种群的年龄结构和相关数量规律是揭示种群数量动态变化和预测种群发展趋势的重要手段(李文英等,2018)。本研究结果表明,3个调查样地的野生红花瘤果茶种群幼龄树、中龄树和老龄树个体数量依次减少,呈现出基部宽,顶部窄的种群金字塔结构,属于增长型种群,种群能进行天然更新并稳定增长,其中兴仁县新寨村调查样地以实生种群为主,晴隆县龙头村调查样地以萌生种群为主。种群的空间分布格局是种群个体在空间上配置与分布的特征表现(袁春明等,2012),红花瘤果茶种群的空间分布主要表现为小尺度的集群分布和大尺度的随机分布,该分布格局可能是由于种子散布受限、生境异质性、种内—种间竞争等相互作用的结果(He & Duncan, 2000;Lin et al., 2011)。本研究中,红花瘤果茶在幼龄、中龄、老龄3个发育阶段的种群分布格局呈现出一定差异性。
1号、3号样地的幼龄树在小尺度上均呈现集群分布,大尺度上呈随机分布或均匀分布,2号样地的幼龄树在所有尺度上均呈集群分布,3个样地中龄树和老龄树几乎在所有尺度上均呈随机分布。这种分布特征与生境特征及其自身生物学特性密切相关,该种群主要分布在喀斯特山区的岩石缝隙和岩坡地,坡度皆>30°,且岩石裸露皆>70%,土壤小块状不连续分布在岩缝中。红花瘤果茶果实一般含有种子3~12颗,种子数量较多,其种子自然脱落后,种子多聚集在适宜萌发的生境,因此幼苗多呈现显著的集群分布。随着幼苗成长,幼苗间种内竞争及与其他物种种间竞争加剧,导致幼苗死亡率升高,生长到中龄树和老龄树阶段的植株减少,因此3个样地中的中龄树和老龄树大多呈现随机分布。
4.2 红花瘤果茶不同发育阶段之间空间关联性的差异明显
种群不同发育阶段的空间关联分析是对一定时期内植物种群个体在空间相互关系的描述,包含空间分布关系、个体间的相互影响,关联分析可以揭示生物种群的现状、反应种群个体的相互作用(张健等,2007)。本研究中,红花瘤果茶幼龄、中龄、老龄不同发育阶段的空间格局关系主要表现为负相关、正相关和不相关。样地1幼龄树与中龄树在所有尺度上均表现为负相关,样地2幼龄树和中龄树在小尺度上为负相关,中大尺度上为不相关。主要是因为1号、2号样地为次生林地,受人为活动的干扰较为严重,中龄树和老龄树结实率较低,大部分幼苗是人为砍伐的中龄树和老龄树的树桩上萌生出的萌生苗,真正由红花瘤果茶种子萌发的实生苗数量较少。样地3幼龄树与中龄树呈现显著的正相关关系,主要是3号样地为准原生植被,植被群落和原生境保护完好,人为活动干扰强度极低,中龄树和老龄树结实率较高,果实和种子受重力脱落,就地萌生产生大量实生苗,加上3号样地种群幼苗绝对数量大,较多的幼龄树能发育成中龄树。1号、2号样地中幼龄树和老龄树均在中小尺度上呈显著负相关,在大尺度上无相关性,3号样地幼齡树和老龄树在小尺度上呈显著负相关,在中尺度上无相关性,大尺度呈显著正相关;3个样地中龄树和老龄树在所有尺度上均无相关性。这主要是由于随着红花瘤果茶自身成长,种内竞争加剧,导致大部分中、老龄个体周围的幼龄个体正常接受的营养和光照受到干扰影响,缺乏生存竞争力而不能保障正常发育,导致了个体的自然死亡。
4.3 种子生物学特性和自然环境因素是红花瘤果茶特有分布的主要原因
本研究从红花瘤果茶种群数量动态分析可以看出,3个样地所代表的种群幼苗补充充裕,种群结构可以定义为增长型,但2号、3号样地Ⅱ龄级个体、1号、2号、3号样地Ⅳ龄级死亡率较高,贡献潜力为负值,在一定程度上限制了种群数量的发展。通过实地调查发现1号、2号样地实生苗数量较少,萌生苗所占比重较大,可能在之前相当一段时间内受到外界人为活动干扰剧烈,如经济采伐、制造农具等致使大量Ⅳ龄级个体被砍伐。再加上喀斯特石灰岩地区基岩裸露率高,土层瘠薄,当幼苗生长到一定程度,瘠薄的土层不能满足其继续生长所需要的营养条件时,导致了大量个体自然死亡。自然因素和人为干扰是限制种群数量扩大的主要因素。另外,贵州高原亚地区喀斯特地貌沟壑纵横、坡陡谷深、地形切割破碎(吴征镒等,2010),而红花瘤果茶果实和种子的个大偏重,种子传播大部分靠重力自然坠落、滚动,纵横交错的山脊和深谷成了限制其种群扩散的主要因素,物种生物学特性和生境异质性是红花瘤果茶种群狭域分布的主要原因。红花瘤果茶种群空间分布格局及数量动态是由其种群自身的生态学特性、种内、种间关系以及原生生境异质性共同作用的结果。红花瘤果茶种群在原生生境能进行自然更新,但由于强烈的人为活动干扰、种内种间竞争及其原生境的特殊性,导致其种质资源难以扩散,种群分布面积狭窄。对小种群的特有珍稀植物保护来说,了解其野生种群生境和相关生态学特征是对其进行引种抚育、迁地保护和开发利用的前提条件,该研究结果可为稀有濒危的红花瘤果茶植物种质资源保护利用提供参考。
参考文献:
AN MT, 2005. Present status of the natural resource of Camellias in Guizhou Province [J]. Guizhou For Sci Tech-nol, 33(2): 26-29.[安明态, 2005. 贵州自然分布山茶属植物的资源现状 [J]. 贵州林业科技, 33(2):26-29.]
CHANG HD, REN SX, 1998. Flora Reipublicae Popularis Sinicae [M]. Beijing: Science Press, 49: 42-44.[张宏达, 任善湘, 1998. 中国植物志 [M]. 北京: 科学出版社, 49:42-44.]
CHEN XD, 1998. A study on the method of quantitative analysis for plant population and community structural dynamics [J]. Acta Ecol Sin, 18(2): 214-217.[陈晓德, 1998. 植物种群与群落结构动态量化分析方法研究 [J]. 生态学报, 18(2):104-107.]
DENG ZZ, PAN DQ, LIU ZB, et al., 2011. Preliminary study on Camellia rubituberculata Chang et Yu in Guizhou Province [J]. Guizhou For Sci Technol, 39(2): 39-42.[邓忠治, 潘德权, 刘正本,等, 2011. 贵州省厚壳红瘤果茶初步研究 [J]. 贵州林业科技, 39(2):39-42.]
FANG JY, WANG XP, SHEN ZH, et al., 2009. Methods and protocols for plant community inventory [J]. Biodivers Sci, 17(6): 533-548.[方精云, 王襄平, 沈泽昊, 等, 2009. 植物群落清查的主要内容、方法和技术规程 [J]. 生物多样性, 17(6):533-548.]
GUO MX, HUANG LH, HONG J, et al., 2018. Seed characte-ristics and biological significance of Camellia lapidea Wu and Camellia rubituberculata Chang ex Lin et Lu [J]. Seed, 37(9): 77-80.[郭米香, 黄丽华, 洪江, 等, 2018. 石果红山茶和红花瘤果茶的种子特性及生物学意义 [J]. 种子, 37 (9):77-80.]
HE FL, DUNCAN RP, 2000. Density-dependent effects on tree survival in an old-growth Douglas fir forest [J]. J Ecol, 88(4): 676-688.
HANG DL, LIANG SC, JIANG Y, 2019. Quantitative dynamics of Cyclobalanopsis glauca population in the Guilin karst mountainous areas [J]. Chin J Ecol, 38(3): 663-671.[黄冬柳, 梁士楚, 姜勇, 2019. 桂林岩溶石山青冈种群数量动态 [J]. 生态学杂志, 38(3):663-671.]
KANG JP, MA YY, MA SQ, et al., 2019. Dynamic changes of spatial pattern and structure of the Tamarix ramosissima population at the desert-oasis ecotone of the Tarim Basin [J]. Acta Ecol Sin, 39(1): 265-276.[康佳鵬, 马盈盈, 马淑琴, 等, 2019. 荒漠绿洲过渡带柽柳种群结构与空间格局动态 [J]. 生态学报, 39(1):265-276.]
LI SF, LIU WD, SU JR, et al., 2013. Age structure and spatial distribution patterns of Taxus yunnanensis population in Lanping County, Yunnan Province [J]. Acta Bot Boreal-Occident Sin, 33(4): 792-799.[李帅锋, 刘万德, 苏建荣, 等, 2013. 云南兰坪云南红豆杉种群年龄结构与空间分布格局分析 [J]. 西北植物学报, 33(4):792-799.]
LI WY, LI X, GAN XH, 2018. Population structure and dynamics of endangered plant Tetracentron sinense [J]. Subtrop Plant Sci, 47(3): 222-228.[李文英, 李欣, 甘小洪, 2018. 濒危植物水青树的种群结构与数量动态 [J]. 亚热带植物科学, 47(3):222-228.]
LIN YC, CHANG LW, YANG KC, et al., 2011. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation [J]. Oecologia, 165(1): 193-207.
LIU GF, DING Y,ZANG RG, et al., 2011. Distribution patterns of Picea schrenkiana var. tianschanica population in Tianshan [J]. Chin J Appl Ecol, 22(1): 9-13.[刘贵峰, 丁易, 臧润国, 等, 2011. 天山云杉种群分布格局 [J]. 应用生态学报, 22(1):9-13.]
LIU HY, YANG NK, ZOU TC, et al., 2016. Population structure and dynamics of the endemic plant Acre guizhouense in Guizhou Province [J]. Guihaia , 36(5): 548-556.[刘海燕, 杨乃坤, 邹天才, 等, 2016. 贵州特有植物贵州槭的种群结构及动态研究 [J]. 广西植物, 36(5):548-556.]
MIN TL, BARTHOLOMEW B, 2008. Flora of China[M]. Beijing: Science Press, 12: 400.[闵天禄, Bartholomew B, 2008. 中国植物志 [M]. 北京:科学出版社, 12: 400.]
WANG L, SUN QW, HAO CY, et al., 2010. Point pattern analysis of different age-class Taxus chinensis mairei individuals in mountainous area of southern Anhui Province [J]. Chin J Appl Ecol, 21(2): 272-278.[王磊, 孫启武, 郝朝运, 等, 2010. 皖南山区南方红豆杉种群不同龄级立木的点格局分析 [J]. 应用生态学报, 21(2):272-278.]
WU ZY, SUN H, ZHOU ZK, et al., 2010. Floristic geography of seed plants in China [M]. Beijing: Science Press: 71-83.[吴征镒, 孙航, 周浙昆, 等, 2010. 中国种子植物区系地理 [M]. 北京:科学出版社:71-83.]
XU M, DONG SB, ZHANG DH, et al., 2107. Study on spatial distribution pattern of Tilia amurensis population in Beijing [J]. J NW Sci Technol Univ Agric(Nat Sci Ed), 45(8): 81-88.[许梅,董树斌, 张德怀, 等,2017. 北京市紫椴种群空间分布格局研究 [J].西北农林科技大学学报(自然科学版), 45(8):81-88.]
YANG B, YUAN CJ, DAI XY, et al., 2016. Study on the flora of distribution areas of Camellia rubituberculata [J].Guizhou For Sci Technol, 44(3): 24-29.[杨冰, 袁丛军, 戴晓勇, 等, 2016. 厚壳红瘤果茶分布区植物区系研究 [J]. 贵州林业科技, 44(3):24-29.]
YANG NK, ZOU TC, LIU HY, et al., 2015. Age structure and spatial distribution pattern of endemic plant Camellia mairei var. lapidea population in Guizhou Province, China [J]. J Trop Subtrop Plant, 23(2): 205-210.[杨乃坤, 邹天才, 刘海燕, 等, 2015. 贵州特有植物长柱红山茶种群年龄结构及空间分布格局研究 [J]. 热带亚热带植物学报, 23(2):205-210.]
YU Q, 2006. Population ecology of Abies fargesii in Shenlongjia area [D]. Beijing: Graduate School of the Chinese Academy of Sciences (Institute of Plant Studies): 2-3.[于倩, 2006. 神农架巴山冷杉林种群生态学研究 [D]. 北京:中国科学院研究生院(植物研究所):2-3.]
YUAN CM, MENG GT, FANG XJ, et al., 2012. Age structure and spatial distribution of the rare and endangered plant Alcimandra cathcartii[J]. Acta Ecol Sin, 32(12): 3866-3872.[袁春明, 孟广涛,方向京, 等, 2012. 珍稀濒危植物长蕊木兰种群的年龄结构与空间分布 [J]. 生态学报, 32(12):3866-3872.]
YUAN CJ, YANG B, DAI XY, et al., 2016. Life table and survivability analysis of natural population of the endemic plant Camellia rubituberculata Chang ex Lin et Lu [J]. J Jiangsu Agric Sci, 44(5): 275-280.[袁丛军, 杨冰, 戴晓勇, 等, 2016. 特有植物厚壳红瘤果茶自然种群生命表及生存力分析 [J]. 江苏农业科学,44(5):275-280.]
ZHANG J, HAO ZQ, SONG B, et al., 2007. Spatial distribution patterns and associations of Pinus koraiensis and Tilia amurensis in broad-leaved Korean pinemixed forest in Changbai Mountains [J]. Chin J Appl Ecol, 18(8): 1681-1687.[张健, 郝占庆, 宋波, 等, 2007. 长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性 [J]. 应用生态学报, 18 (8):1681-1687.]
ZHANG JT, 1998. Analysis of spatial point pattern for plant species [J]. Chin J Plant Ecol, 22(4): 344-349.[张金屯, 1998. 植物种群空间分布的点格局分析 [J]. 植物生態学报, 22(4): 344-349.]
ZHANG JT, MENG DP, 2004. Spatial pattern analysis of individuals in different age-classes of Larix peincipis-rupprechtii in Luya mountain reserve, Shanxi, China [J]. Acta Ecol Sin, 24(1):35-40.[张金屯, 孟东平, 2004. 芦芽山华北落叶松林不同龄级立木的点格局分析 [J]. 生态学报, 24(1):35-40.]
ZHANG XW, ZHANG XP, GUO CY, et al., 2013. Point pattern analysis of Pteroceltis tatarionwii population at its different development stages in limestone mountain area of North Anhui, East China [J]. Chin J Ecol, 32 (3):542-550.[张兴旺, 张小平, 郭传友, 等, 2013. 皖北石灰岩山地青檀种群不同发育阶段的点格局分析 [J]. 生态学杂志, 32 (3):542-550.]
ZHONG ZQ, 2015. Study on the spatial distribution pattern of natural Castanopsis carlesii popolation in Northern Fujian [J]. For Eng, 31(3): 31-34.[钟兆全, 2015. 闽北天然米槠种群空间分布格局研究 [J]. 森林工程, 31(3): 31-34.]
ZHOU L, WANG Y, ZHANG YZ, et al., 2015. The fruit characteristics and seed germination characteristics of Camellia rubituberculata Chang ex Lin et Lu [J]. N Hortic, (2): 34-37.[周丽, 王苑,张应祝, 等, 2015. 厚壳红瘤果茶果实特征与种子萌发特性 [J]. 北方园艺,(2):34-37.]
猜你喜欢
世界热带农业信息(2022年8期)2022-07-19
种子科技(2021年8期)2021-07-11
长江文艺(2017年8期)2017-08-15
晚晴(2016年5期)2016-05-14
辽河(2015年4期)2015-09-15
安徽农学通报(2014年16期)2014-09-18
晚晴(2009年9期)2009-10-10
晚晴(2009年4期)2009-04-27
农村百事通(2009年4期)2009-03-02
雕塑(1997年4期)1997-06-28
