
镉胁迫对万寿菊属植物幼苗生理及富集的影响
2019-09-10 07:22:44刘翰升赵春莉刘玥国伟强
福建农业学报 2019年10期
关键词:生理指标
刘翰升 赵春莉 刘玥 国伟强


摘要:【目的】研究不同质量浓度(0、0.3、3、20、60、120、180、240mg.L)镉胁迫下,万寿菊属植物幼苗(万寿菊、孔雀草)的生理指标与富集指标,比较可溶性蛋白、丙二醛、过氧化物酶、超氧化物歧化酶与镉吸收量,明确不同质量浓度镉胁迫下万寿菊属植物幼苗生理调节机制与富集能力。【方法】通过水培试验,采用镉胁迫处理幼苗,测定可溶性蛋白、丙二醛、过氧化物酶、超氧化物歧化酶与镉吸收量,研究镉胁迫对万寿菊、孔雀草幼苗的生理指标与幼苗富集效果的影响。【结果】随着Cd胁迫质量浓度的增加,万寿菊幼苗的可溶性蛋白质含量逐渐降低,丙二醛含量随Cd质量浓度增加先降低后升高再降低,过氧化物酶活性逐渐升高,超氧化物歧化酶活性先降低后升高再降低;孔雀草幼苗可溶性蛋白质含量逐渐降低,丙二醛含量先下降后上升再下降后又上升,过氧化物酶先上升后下降,超氧化物歧化酶含量先上升后下降再上升后下降;万寿菊、孔雀草幼苗对镉的吸收量随Cd胁迫质量浓度的增加积累能力逐渐增强,万寿菊幼苗与孔雀草幼苗在180mg·LCd胁迫时,幼苗内Cd含量分别达到440.40、478.23mg.kg。【结论】中、低量的Cd胁迫(0.3-120mg.L)对万寿菊、孔雀草幼苗生理影响小,自身可以调节,高量的Cd胁迫(180、240mg.L)产生破坏作用,影响较大;万寿菊、孔雀草幼苗富集量随Cd胁迫质量浓度升高而升高,直到幼苗不萌发;万寿菊幼苗抵抗Cd胁迫生理机制过氧化物酶起主导作用,孔雀草幼苗抵抗Cd胁迫生理机制超氧化物歧化酶起主导作用;采用万寿菊、孔雀草种子进行植物修复有较大应用前景。
关键词:万寿菊属;镉胁迫;生理指标;镉富集
中图分类号:S681.9
文献标志码:A 文章编号:1008-0384(2019)10-1221-07
0引言
【研究意义】伴随着我国经济持续增长,工业“三废”:农业化肥与农药的不合理施用,致使我国环境日渐恶劣、土壤污染日趋严重,使重金属污染成为当今面积最广、危害性最大的环境问题之一,由于重金属的不可降解性,在环境中只能物质之间的相互迁移转换,所以重金属污染的消除往往更为困难,对生物引起的影响和危害使人类更为关注。重金属镉污染对植物具有毒害作用,对我国作物产量、环境安全及植物生长等方面造成了巨大影响,万寿菊属植物对重金属镉胁迫有较强的抗性,为我国植物修复技术领域提供新的思路。【前人研究進展】目前,关于镉胁迫植物生理的研究主要集中于低浓度镉胁迫下植物的可溶性蛋白、MDA、抗氧化酶系统等指标的变化。镉富集植物研究多集中于超富集植物,但因其生物量小、生长缓慢、地域局限导致难以大规模应用,万寿菊与孔雀草全国各地均可种植,具有生长速度快、生物量大、观赏性强等特点,已有研究表明孔雀草是新型的耐镉植物。研究万寿菊及孔雀草在不同质量浓度镉胁迫下的幼苗生理响应与富集指标,对万寿菊色素提取产业、美化环境、植物修复技术发展等有较大意义。【本研究切入点】目前植物修复技术存在超富集植物生长缓慢、生物量小等问题,适用于高浓度重金属污染地区植物修复的研究未见报道,针对重金属污染,高效、实用的修复技术并不多。万寿菊属植物对重金属镉胁迫的耐性较强,且万寿菊相比较于一般超富集植物生物量大,栽培密度高,采用种子的方式进行播种节约成本,发芽初期种子生长迅速,富集效率相对高。【拟解决的关键问题】本试验对万寿菊属植物种子在不同质量浓度镉胁迫下对植物种子幼苗生理生化指标与镉吸收量方面进行研究,比较不同质量浓度镉胁迫对可溶性蛋白、丙二醛、过氧化物酶、超氧化物歧化酶等生理指标与镉吸收能力,以期为万寿菊属植物种子在镉胁迫下的生理生化响应机制提供更为详细的数据模型,为以直接播种的方式进行植物修复提供理论依据,为植物修复技术的发展提供理论基础。对我国减轻重金属污染,进行生态修复具有一定的指导意义。
1材料与方法
1.1试验材料
供试材料为万寿菊(Tagetes erecta L.)、孔雀草(Tagetes patula L.)种子。试验于2018年9月到2018年10月在吉林农业大学园艺学院园林专业实验室进行。
1.2试验方法
1.2.1发芽试验 依据中华人民共和国国家土壤环境质量标准(GB36600-2018和GBl5618-2018)以及预试验所得结果,设置Cd质量浓度为0(CK)、0.3、3、20、60、120、180、240mg.L,设置万寿菊、孔雀草各处理分别为A1、A2、A3、A4、A5、A6、A7、A8,Bl、B2、B3、B4、B5、B6、B7、B8。cd供源由分析纯CdCl2·2.5H2O配制所得,将cd以溶液的形式加入到PC培养皿中,模拟土壤受到不同程度的重金属污染。在超净工作台内,每份试材选取50粒籽粒饱满、大小均一的种子,整齐摆放在垫有双层滤纸已灭菌的PC培养皿中,每个培养皿中添加8mL不同浓度梯度的Cd溶液,以添加8mL无菌水为对照,进行单因素试验。试验在模拟最优自然生长环境的人工气候箱中进行,温度设置为(19±4)℃,相对湿度设置为50%-90%,光照设置为0-3级,24h暗处理后,培养7d,3次重复,以此获得幼苗进行生理与富集试验。
1.2.2生理试验万寿菊、孔雀草幼苗中可溶性蛋白质含量采用考马斯亮蓝G-250染色法测定;MDA含量采用硫代巴比妥酸法;POD活性用愈创木酚法测定;SOD活性的测定采用NBT光还原法。
1.2.3富集试验将植株种皮摘掉后,冲洗3次,后用干燥滤纸吸掉植株表面的水分,每处理使用分析天平取10g,剪碎后装在聚四氟乙烯消解管中,编号。加入5mL硝酸浸泡过夜,第二天消解前加入0.5mL高氯酸,然后先用石墨消解仪180%消解45min,再将温度升高到220%继续消解到黄棕色烟雾完全冒尽,加入2mL过氧化氢,继续加热到白烟冒尽,溶液透明,消解结束后冷却到室温,定容至25mL,摇匀。使用原子吸收分光光度计测定溶液浓度。
1.3数据处理
采用SPSS20.0、Word2013对数据进行Duncan新复极差法显著性分析、数据整理与制作图表。
2结果与分析
2.1镉对万寿菊幼苗生理指标的影响
2.1.1镉对万寿菊幼苗可溶性蛋白含量的影响
植物体内的可溶性蛋白质大多数是参与各种代谢的酶类,测其含量是了解植物体总代谢的一个重要指标,是衡量植物抗逆性的重要指标。由表1可知,随着镉胁迫质量浓度的增加,万寿菊幼苗可溶性蛋白含量随之降低,A2处理与A1相比差异不显著,A3处理与A1处理相比极显著下降,下降6.82%,A3、A4、A5处理差异均不显著,A6、A7处理与A1相比极显著降低,分别下降19.55%和30.91%。从万寿菊幼苗可溶性蛋白的质量分数表明,低浓度镉胁迫对万寿菊幼苗可溶性蛋白的形成几乎无抑制,中低浓度镉胁迫对万寿菊幼苗可溶性蛋白的形成抑制作用较弱且抑制效果稳定,但随着镉胁迫质量浓度的增加,对万寿菊幼苗可溶性蛋白形成的抑制作用逐渐增强。
2.1.2镉胁迫对万寿菊幼苗丙二醛含量的影响丙二醛含量是植物细胞膜质过氧化程度的体现,丙二醛含量高,说明植物细胞膜质过氧化程度高,细胞膜受到的伤害严重。由表1可知,A,处理与A,处理相比,镉胁迫对万寿菊幼苗丙二醛含量几乎生成无促进,A3处理丙二醛含量相较于A1处理表现为极显著下降,下降48.99%。A4、A5处理丙二醛的含量均显著高于A1处理,上升40.47%和39.22%,此时万寿菊幼苗内丙二醛大幅度生成;A6、A7处理与A】处理相比极显著降低,分别下降6.82%和8.22%,此时万寿菊幼苗丙二醛生成再次被抑制。万寿菊幼苗丙二醛质量分数变化表明,低浓度镉胁迫对万寿菊幼苗丙二醛含量影响不大,中低浓度的镉胁迫对万寿菊幼苗丙二醛含量产生抑制,对万寿菊幼苗的细胞膜伤害性逐渐增加。
2.1.3镉对万寿菊幼苗抗氧化酶含量的影响抗氧化酶具有将体内形成过氧化物转换为毒害较低或无害的物质的功能,抗氧化酶的活性是植物能否对逆境适应的重要指标之一。由表1可知,随着Cd胁迫质量浓度的增加,过氧化物酶活性极显著增加,A7处理时达到最大值,处理A2、A3、A4、A5、A6、A7分别达到处理A1的1.08、1.60、1.88、1.97、2.69、4.24倍。超氧化物歧化酶活性与过氧化物酶活性呈相反状态,随着Cd胁迫质量浓度的增加,SOD的活性先降低后升高再降低,A7处理时达到最小值,与A1处理存在极显著性差异,下降58.19%。随着镉胁迫质量浓度的增加,对万寿菊幼苗过氧化物酶活性的促进作用逐渐增强;低浓度镉胁迫对万寿菊幼苗超氧化物歧化酶活性抑制明顯,中低浓度镉胁迫对万寿菊幼苗超氧化物歧化酶活性有一定促进作用,但随着镉胁迫质量浓度的增加,万寿菊幼苗超氧化物歧化酶活性的抑制作用再次逐渐增强。
2.2镉对孔雀草幼苗生理指标的影响
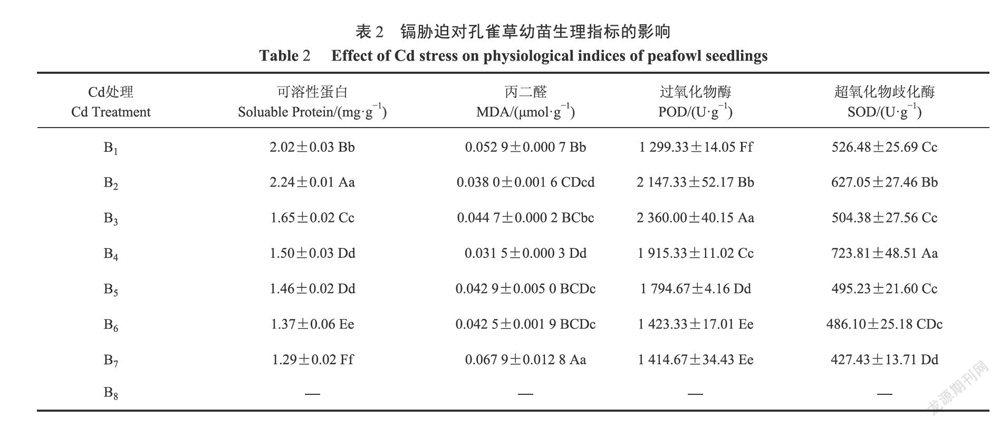
2.2.1镉对孔雀草幼苗可溶性蛋白含量的影响由表2可知,随着镉胁迫质量浓度的增加,孔雀草的可溶性蛋白质含量呈现先升高再下降的态势,B2处理相较于B,处理可溶性蛋白含量极显著增加,极显著促进可溶性蛋白的生成,与B1相比上升了10.89%;B3、B4、B5、B6、B7处理与B1处理相比均极显著下降,分别下降了18.32%、25.74%、27.72%、32.18%和36.14%,B4、B5处理无显著差异。低浓度镉胁迫对孔雀草可溶性蛋白的形成促进作用较强,但随着镉胁迫质量浓度的增加,孔雀草幼苗可溶性蛋白形成的促进作用转为抑制作用,并逐渐增强。
2.2.2镉对孔雀草幼苗丙二醛含量的影响
由表2显示,随着镉胁迫质量浓度的增加,孔雀草幼苗丙二醛的含量呈现先减后增的趋势,B2处理丙二醛的含量与B1处理相较极显著下降,分别降低了28.17%,B3处理丙二醛的含量随后上升,与B1处理相较差异不显著,B4处理丙二醛的含量再次极显著降低,与B1处理相较下降40.45%,Bs、B6处理较B4处理显著增高,但B5、B6处理本身差异不显著,B7处理丙二醛含量极显著增高,达到峰值。低中浓度镉胁迫对孔雀草幼苗丙二醛生成抑制效果显著,随着镉胁迫质量浓度的增加,丙二醛生成抑制效果逐渐消失转为促进。
2.2.3镉对孔雀草幼苗抗氧化酶活性的影响表2可知,在不同质量浓度镉处理下,对孔雀草幼苗过氧化物酶、超氧化物歧化酶活性的影响。随着镉胁迫质量浓度的增加,过氧化物酶的活性先升高再降低,B2、B1处理过氧化物酶的活性呈上升趋势,与B1处理相比极显著升高,分别上升了65.26%、81.63%;B4、Bs、B6、B7处理与B1处理相比极显著上升,但上升趋势减小,分别上升了47.41%、38.12%、9.54%和8.87%。超氧化物歧化酶活性先升高后降低再升高后降低,B2处理与B1处理相比极显著升高,上升了19.10%,B3处理与B1处理无显著差异,B4处理与B,处理相比极显著升高,上升了137.48%,B5、B6处理与B1处理无显著差异,B7处理与B1处理相比表现为极显著降低,降低了18.81%。低浓度镉胁迫,对孔雀草幼苗过氧化物酶活性的促进作用较强,随着镉胁迫质量浓度的增加,中高浓度镉胁迫对孔雀草幼苗过氧化物酶活性的促进作用逐渐减弱;低浓度镉胁迫对孔雀草幼苗超氧化物歧化酶活性促进明显,随后抑制,中高浓度镉胁迫对万寿菊幼苗超氧化物歧化酶活性有一定促进作用,但随着镉胁迫质量浓度的增加,对万寿菊幼苗超氧化物歧化酶活性的抑制作用再次逐渐增强。
2.3镉对万寿菊及孔雀草幼苗富集指标的影响
从图1可知,镉胁迫万寿菊的Cd含量随着Cd浓度的增加而逐渐增大,A2、A3处理与A1处理相比差异不显著,镉含量分别为0.37和1.57mg·kg;A4、A5、A6、A7处理与A1处理相比极显著升高,镉含量分别达到了22.9、87.27、128.30和440.40mg·kg;孔雀草幼苗镉含量随着镉胁迫质量浓度的增加而上升,B2、B3处理与B1处理相比差异不显著,镉含量分别达到了0.47和1.83mg.kg;B4、B5、B6、B7处理与B1处理相比极显著升高,镉含量分别达到了23.10、76.63、138.63和478.23mg·kg,通过柱形图对比,镉胁迫对万寿菊、孔雀草在幼苗体内的积累含量差异不大,基本相似。
3讨论与结论
可溶性蛋白作为渗透调节作用的重要物质,其含量在一定程度上可以自身调节。可溶性蛋白含量在万寿菊、孔雀草叶中均随cd浓度的增加表现为先增高后减少,可溶性蛋白含量越多,越能促进功能蛋白数量增加,对植物细胞维持正常代谢起促进作用,但过量镉胁迫对蛋白质合成起抑制作用,但过量的cd造成了万寿菊体内蛋白质的分解和运输受阻,促进蛋白水解酶活性,导致原有蛋白质分解加速,且cd对合成蛋白质的有关酶类、细胞器具有毒害性和破坏作用,使新蛋白合成受到了抑制,最终导致万寿菊及孔雀草体内的可溶性蛋白质含量大幅度下降。李建龙等提出随着重金属胁迫增强,油菜(Brassica napus L.)体内光合系统产生破坏,导致蛋白质分解及运输受阻,最终导致蛋白质含量下降,与本试验研究结果相似。
镉胁迫会诱变植物的质膜发生过氧化,细胞膜损伤,与外界的通透性增强,因自由基作用的脂质发生过氧化反应,最终生成具有细胞毒性的氧化终产物丙二醛。万寿菊幼苗丙二醛含量随着cd浓度增加,与对照比较表现先升高后降低的现象,但在3mg·L时表现下降趋势。彭昌琴等研究表明,在重金属镉的胁迫作用下,仅在一定范围内对植物的生长具有抗逆性,当超过这个极限浓度,植物自身的抗性就会消失,这与本试验中万寿菊幼苗丙二醛含量的先下降再上升后下降,万寿菊幼苗对低质量浓度镉胁迫伤害的抗逆性显著,后随着质量浓度升高,丙二醛含量突然剧增后逐渐下降,表明植物自身抗性逐渐减弱,生长发育受到严重影响的结果相类似。孔雀草幼苗丙二醛含量随cd浓度的逐渐增加表现先降低后升高。刘柿良等提出,不同浓度镉处理下龙葵(Solanum nigrum L.)幼苗丙二醛含量会随着cd质量浓度的升高而增加,这与本试验中孔雀草幼苗丙二醛含量的先下降后升高的结果不一致,应是孔雀草幼苗对镉拥有较强抗逆性强,直到180mg·L时,脂质过氧化反应才剧烈进行的原因。
过氧化物酶可作为组织老化的一种生理指标,与植物光合作用及呼吸作用、生长素养化均有关;超氧化物歧化酶对细胞免疫损伤起保护作用,在活性氧清除系统中扮演重要角色。万寿菊幼苗随cd浓度的增加,过氧化物酶活性呈现逐渐上升趋势,超氧化物歧化酶活性呈现逐渐下降趋势,徐旭等提出cd浓度超过一定范围后,导致过氧化物酶、超氧化物歧化酶活性降低甚至失活,这与本试验中万寿菊幼苗超氧化物歧化酶活性随着镉胁迫质量浓度的增加而降低相一致,但与本试验中万寿菊幼苗过氧化物酶活性随着镉胁迫质量浓度增加不同,这应是万寿菊幼苗过氧化物酶体系较一般植物抗逆性更强导致的。孔雀草幼苗过氧化物酶、超氧化物歧化酶活性均呈现先升高后下降的趋势,刘周莉等提出,低浓度镉胁迫对超氧化物歧化酶的产生有促進作用,活性增强对植物有保护作用,在25mg.LCd胁迫之后,超氧化物歧化酶活性降低,这与本试验中孔雀草幼苗超氧化物歧化酶活性随着镉胁迫质量浓度的增加先升高后降低相一致。
镉超富集植物的临界标准100mg·kg:本试验中,万寿菊幼苗与孔雀草幼苗在180mg·L镉胁迫时,幼苗内镉含量分别达到440.40、478.23mg·kg,120mg.LCd胁迫时128.30、138.63mg-kg,与张银秋研究的植物对镉的吸收、转运及积累随着植物种类的不同而有所差异结果相似。修复浓度在100mg·kg以上的土壤,镉吸收效率高,每7d生物量为1kg的富集量可以达到镉超富集植物一个生长周期的富集量。刘威等提出宝山堇菜Viola baoshan-ensis 150d富集量为233-1846mg.kg,这与本文中采用万寿菊与孔雀草种子以相同方法按150d计算,富集量分别可以达到9437.14mg·kg与10247.79mg.kg,采用万寿菊属植物种子进行植物修复镉胁迫有更大的应用范围与空间。
植物在镉胁迫的逆境条件下,镉会对植物造成严重损害,最直接的表现是植物的生理生化的反应。万寿菊、孔雀草体内的可溶性蛋白质含量、丙二醛含量、过氧化物酶、超氧化物歧化酶的活性与cd的浓度有关,cd浓度越高,对植物的生理影响越大,镉对万寿菊属植物幼苗的富集能力随浓度增大而增强。镉对万寿菊和孔雀草具有多方面的伤害,包括镉胁迫减少植物可溶性蛋白质的合成,改变其膜的通透性,降低酶活性等生理生化过程。万寿菊属植物在相同浓度镉胁迫下,生理抵抗机制表现为不同的反应,万寿菊幼苗抵抗镉胁迫生理机制中过氧化物酶不断提高,起主导作用,孔雀草幼苗抵抗镉胁迫生理机制中超氧化物歧化酶起主导作用,与丙二醛完全相反,但富集指标影响大体一致,具体机制还有待进一步研究。
猜你喜欢
现代农业科技(2017年1期)2017-03-06 01:26:03
中西医结合心血管病电子杂志(2016年22期)2017-03-03 00:42:57
江苏农业科学(2017年1期)2017-02-27 11:11:38
南方农业·下旬(2016年8期)2017-02-23 19:53:16
吉林农业·下半月(2017年1期)2017-02-16 11:31:56
江苏农业科学(2016年8期)2017-02-15 20:29:25
江苏农业科学(2016年8期)2017-02-15 20:14:21
安徽农学通报(2017年1期)2017-02-15 18:23:48
物联网技术(2016年12期)2017-01-21 22:27:52
中国科技博览(2016年1期)2016-04-25 07:47:28
