
三峡库区消落带立柳(Salix matsudana)生长及营养元素分配特征
2019-09-05 04:50吴科君马文超陈红纯何欣芮
生态学报 2019年14期
吴科君,马文超,李 瑞,陈红纯,黄 超,何欣芮,魏 虹,*
1 三峡库区生态环境教育部重点实验室,重庆市三峡库区植物生态与资源重点实验室,西南大学生命科学学院, 重庆 400715 2 攀枝花市旅游局, 攀枝花 617000
三峡水库自建成运行后,实行“冬蓄夏排”的反季节性水位调度模式,在每年的10月底到达最高水位175 m,次年6至9月维持在最低水位145 m[1],形成了库岸线长2000 km、消涨幅度为30 m、面积约350 km2的消落带[2]。库区消落带内大量原生植物因不能适应反季节长时间水淹而大面积消亡,造成消落带生物多样性降低,生态屏障功能减退以及景观恶化等环境问题[3]。为恢复库区消落带生态系统正常的结构和功能,利用适生物种进行植被重建被认为是最有效的手段之一[4- 6]。近年来,研究者逐渐将前期模拟研究筛选出来的耐水淹物种用于消落带植被重建示范工作中。立柳(Salixmatsudana)作为库区本土植物,因其耐水淹、生长迅速和成活率高等特性被用于库区165 m以上消落带植被重建[7- 8]。
前期模拟实验研究发现,在水淹胁迫下立柳可通过改变形态特征、光合效率以及提高抗氧化防御系统中酶的活性和增强次生代谢等来提升对水淹环境的耐受能力[7- 10]。随着库区消落带植被重建与恢复工作的展开,基于原位的试验研究逐渐增多,研究者通过对植物的生长情况、分配位置以及生理生态机制等方面的研究[11- 13],丰富了立柳等重建物种对库区“冬蓄夏排”的水文节律的响应机制。已有研究表明植物体内各器官中的营养元素对植物生长和各种生理功能起着重要的作用[14]。营养元素的不足会限制植物生长、发育和代谢,进而影响库区消落带植被修复与重建的可持续性[10,15- 16]。而高强度的水淹会显著影响植物各器官中营养元素的积累与分配方式,并影响到植株的生长[17- 21]。因此,通过原位试验探究在三峡库区消落带立柳的营养元素积累特征,对了解该物种对库区水位变化的适应及可持续性生长是必要的。
2012年3月,本课题组在位于重庆忠县石宝镇共和村汝溪河流域的三峡库区消落带植被修复示范基地165—175 m海拔区域内种植生长均匀的2年生立柳植株。经过3个水淹周期后,植株生长良好,为进一步探讨立柳对库区消落带冬季水淹的适应机理,于2015年5月中旬展开实地采样与研究,分析在经历了冬季水淹后的退水初期立柳各器官中营养元素的积累和分配特征,以期为消落带植被重建工作提供理论和实践指导。
1 材料与方法
1.1 研究材料
立柳(S.matsudana)是三峡库区原有的多年生库岸树种,又称龙爪柳,是旱柳的一个变种,属杨柳科,柳属落叶乔木。本研究选择三峡库区汝溪河流域消落带植被修复示范基地(107°32′—108°14′E,30°03′—30°35′N)作为试验样地,其中在基地内海拔为165—175 m之间以1 m×1 m的密度种植了立柳纯林,该林地面积约4 hm2。2012年3月种植时平均株高为1.613±0.13 m,基径0.288±0.045 cm,冠幅0.492±0.034 m2。试验样地3个采样带中土壤元素含量见表1。
表中数值为平均值±标准误(n=10);同列不同小写字母分别表示处理之间有显著差异(P<0.05)
1.2 试验设计及取样
于2015年5月中旬进行实地取样,取样时整个立柳纯林中林木保存率约为95%。根据每个淹水周期水淹时长和深度的差异将示范基地划分为3个采样带:175 m的对照组(CK)、170 m的中度水淹组(MS)以及165 m的深度水淹组(DS)。各采样带3个水淹周期内水淹情况见表2。
为尽可能减小阳光照射等因素对立柳的影响,分别在3个样带内随机选取长势均一的立柳样木,使用高枝剪于树冠中上层的东、南、西、北四个方位随机采集叶和枝条,然后将各方位的叶片、枝条单独混合后装入自封袋;用根钻以植株基部为圆心,0.5 m半径等距离钻取植株根样(直径2—5 mm),混合装于自封袋,每个采样带设置5个重复。样品带回实验室后,先用超纯水将其清洗干净,然后置于105℃的烘箱内杀青5分钟,再置于60℃的烘箱中烘干至恒重,最后将烘干后的样品放入球磨仪中粉碎后,封装待测。
在采样的同时,在3个样带随机选取立柳各10株,原位记录立柳的株高、基径和冠幅生长指标。使用卷尺测定每株立柳的株高和冠辐,用数显游标卡尺测量基径。
1.3 营养元素含量测定
将待测的植物样品粉碎、过筛后,称取0.005 g,采用Vario EL cube CHNOS元素分析仪(Elementar,德国)进行全N含量测定;另外称取0.05 g使用SpeedWaveMWS- 4微波消解仪(Berghof,德国)消解待测样后,采用ICAP 6000电感耦合等离子体发射光谱仪(Thermo,美国)测定立柳根、枝条及叶片P、K、Ca、Mg、Fe、Mn、Cu、Zn含量。
1.4 数据分析
使用统计分析软件SPSS 22.0进行数据处理以及统计分析,用配对样本t检验的方法(Paired sample T test)比较各处理在种植初期和种植3 a后的生长差异;采用单因素方差分析(One-way ANOVA)检验水淹处理对立柳生长和营养元素含量的影响,用Turkey法进行多重比较。使用Origin 8.5软件制图。
2 结果与分析
2.1 生长状况
图1所示为库区消落带示范基地内不同处理组立柳的生长状况。经过3 a的生长,与种植初期相比,3个采样带的立柳株高、基径和冠幅均有极显著增加(P<0.01)。随着海拔位的降低,水淹时长和深度增加,立柳的生长受到明显抑制。经过3个水淹周期后,与CK组相比,MS组和DS组植株株高分别降低了18.4%和48.7%,DS组立柳的基径和冠幅分别降低了50.3%和71.5%,但MS组的基径和冠幅没有显著差异。

图1 立柳植株生长状况Fig.1 The growth situations of S. matsudana图中数值为平均值±标准误 (n=5);3个采样带中的立柳种植时间和各初始生长指标一致: *表示各处理组在2个时期有极显著差异(P<0.01),相同字母表示2015年5月不同处理组间没有显著差异(P>0.05),不同小写字母表示各处理组之间有显著差异 (P<0.05)
2.2 水淹胁迫对立柳各器官中N、P、K含量的影响
N、P、K是植物生长发育所需要的大量元素。库区3个采样带的立柳对N、P、K元素含量在植株根、枝、叶中的分配表现出了类似的特征。从总体上来看,这3种大量元素在立柳叶片中的含量均显著高于在根和枝条中的含量(图2)。
在经历了不同强度的冬季水淹后,3个采样带内立柳的根、枝条和叶片中N、P、K元素的含量变化表现出不同的特征。水位变化对立柳叶和枝条中N、P元素含量无显著影响,但显著降低了水淹组根中N、P元素含量(P<0.05)。MS组和DS组N元素含量较CK组分别降低了28.2%和26.6%,P元素则分别降低了37.5%和15.2%。水位变化未对立柳叶和根中的K元素含量造成显著影响,但DS组枝条中钾含量显著低于MS组和CK组(图2)
不同处理组的立柳叶、枝条及根中N、P、K含量分别为5.2—23.7 g/kg、0.9—4.4 g/kg和5.3—13.6 g/kg,即为各器官干质量的0.52%—2.37%、0.09%—0.44%以及0.53%—1.37%,这些值均处于植物含量正常水平[22]。
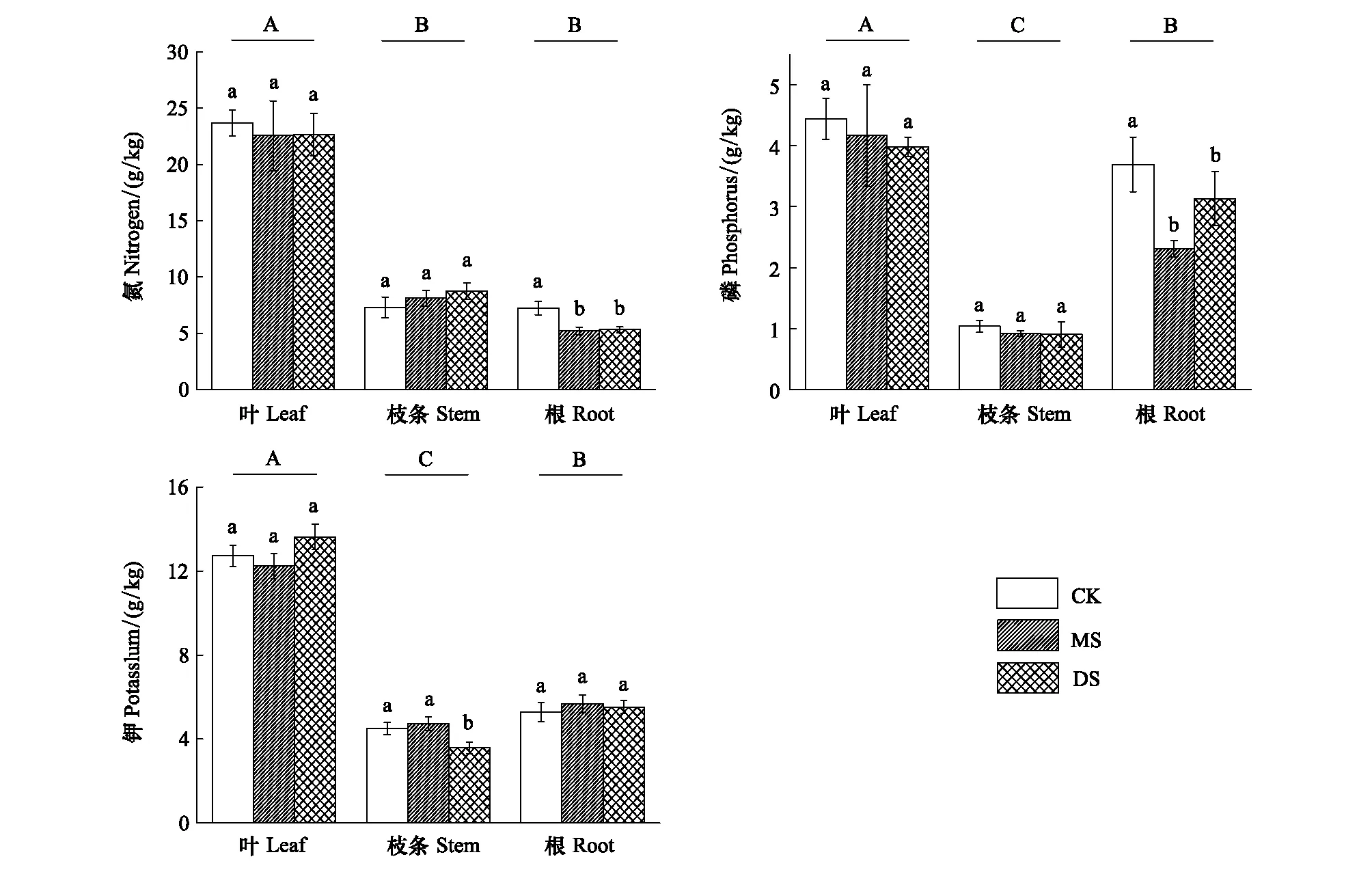
图2 不同水淹处理下立柳N、P、K元素在叶、枝条及根中的含量变化Fig.2 Changes of N, P and K contents in leaves, stems and roots of S. matsudana under different flooding treatments图中数值为平均值±标准误 (n=5);不同小写字母表示同一组织各处理之间有显著差异 (P<0.05);不同大写字母表示各器官间有显著差异(P<0.05)
2.3 水淹胁迫对立柳各器官中Ca、Mg含量的影响
Ca和Mg元素是植物体内重要的中量元素。从各器官中元素分配的总体情况来看,Ca元素表现为根中的含量明显低于叶和枝条,而Mg在各组织中基本均匀分布。
从图3可以看出,冬季水淹对3个采样带中立柳植株的Ca、Mg元素含量影响显著。在立柳的叶和枝条中,DS组中Ca元素含量受到水淹胁迫而显著降低(P<0.05),而CK组与MS组之间无显著差异;在立柳的根中则表现为MS组含量显著高于CK组和DS组,水淹抑制了DS组立柳叶片对Mg的吸收和积累,促进了立柳枝条对Mg的积累,而抑制了立柳根对Mg的吸收和积累各器官不同处理组中钙、镁的含量分别为7.9—31.3 g/kg和1.1—2.1 g/kg,仍处于植物正常元素含量范围[22]。
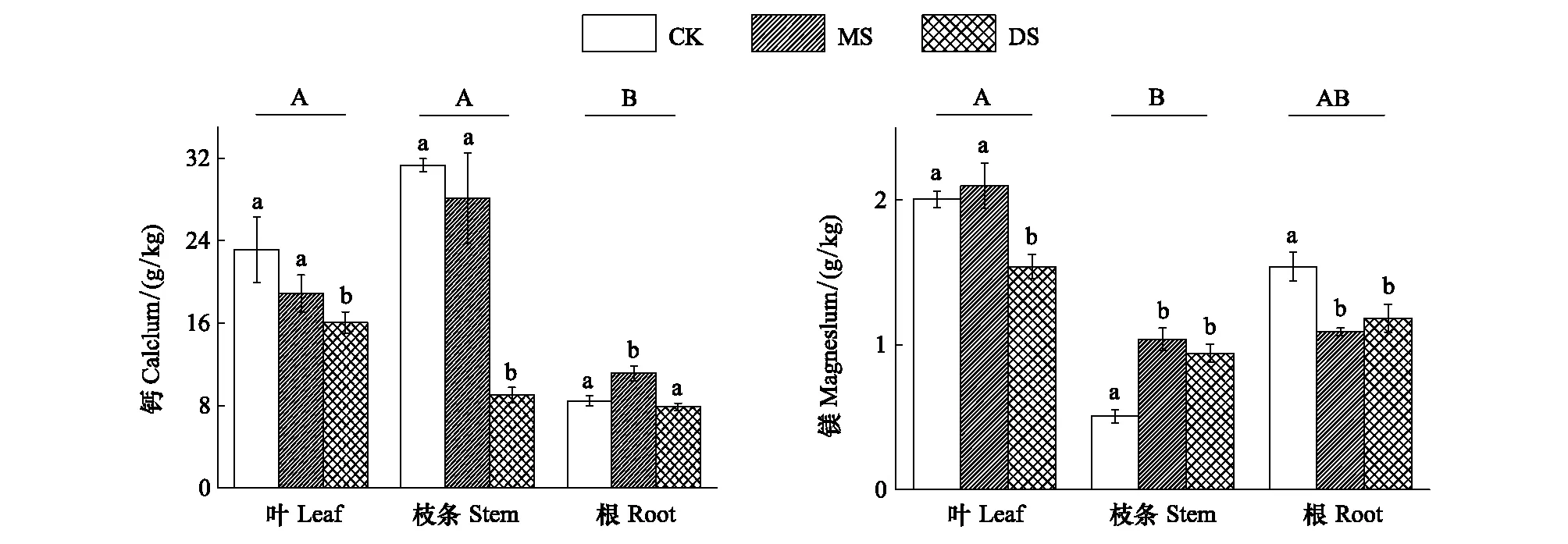
图3 不同水淹处理下立柳Ca、Mg元素在叶、枝条及根中的含量变化Fig.3 Changes of Ca and Mg contents in leaves, stems and roots of S. matsudana under different flooding treatments
2.4 水淹胁迫对立柳各器官中Fe、Mn、Cu、Zn含量的影响
Fe、Mn、Cu、Zn是植物体内重要的微量元素,其含量变化会直接影响植株的生理功能。这4种微量元素在立柳植株各器官中受水淹胁迫影响而表现出不同的分配特征。由图4可知,Fe元素在立柳叶、枝条和根中总体的表现为大量分配到立柳的根中;Mn元素分配情况总体表现为叶>根>枝条;Cu和Zn元素则主要分布在立柳枝条中。各处理组不同器官中营养元素Fe、Mn、Cu、Zn的含量均未低于植物体内的正常水平[22]。
由图4可知,水淹胁迫显著增加了根系中Fe元素(P<0.05),深度水淹(DS处理组)提高了立柳枝条中Fe元素含量,但水淹胁迫对叶片Fe含量的影响不显著。此外受水淹影响,MS组和DS组立柳叶中Mn含量较对照组显著降低(P<0.05),而在枝条和根中,水淹组含量均高于对照组。水淹处理极显著地降低了 DS组枝条中Cu和Zn的含量,同时降低了MS组和DS组根中的元素含量。

图4 不同水淹处理下立柳Fe、Mn、Cu和Zn元素在叶、枝条及根中的含量变化Fig.4 Changes of Fe、Mn、Cu and Zn contents in leaves, steams and roots of S. matsudana under different flooding treatments
3 讨论
在经历了3个水淹周期之后,各海拔高程的立柳生长状况和营养元素含量表现出显著的差异性。与种植初期相比,立柳株高、基径和冠幅均有显著增加,表明该物种对三峡库区消落带冬季水淹具有良好的适应性,并在退水后具有较好的恢复生长能力。良好的生长状态有利于植株争取更多的生长空间和资源,从而更好地应对三峡库区消落带周期性水淹[23]。但立柳的株高、基径和冠幅等生长指标均随着海拔高度的降低即水淹胁迫强度的增加而逐渐降低。与175 m海拔(无水淹胁迫)和170 m海拔(中度水淹胁迫)区域的植株相比,经过3 a的冬季水淹后,165 m海拔(深度水淹胁迫)处立柳的生长受到了明显抑制。生长于该海拔处的植株冬季最大水淹深度为10 m,水淹时间超过5个月,高强度的水淹导致植物光合产物的消耗明显增加,退水后恢复生长的难度也加大,植株光合产物的净积累量减少[24- 26]。
耐水淹植物能够通过生理生态的变化来适应淹水胁迫,其中通过改变营养元素积累与分配来适应水淹是重要的手段之一[10]。水淹导致植物因根系缺氧而引起根际功能紊乱甚至死亡,同时还会影响土壤中营养元素的含量以及有效性,最终显著影响植物根系对营养元素的吸收利用[27]。本研究结果显示,库区消落带水位周期性的变化导致水淹组立柳根中的N、P元素含量显著低于对照组,但各处理组叶和枝条中氮、磷元素的含量无显著差异。植物叶中N、P元素的含量在一定范围内与植物的光合速率成正比[28- 29],立柳叶片中较高的N、P元素含量,保证了植株正常高效的光合效率,从而保证了水淹组植株正常生长存活。K是维持类囊体膜上pH梯度的平衡离子[30],也是多种光合酶的活化剂,同时还参与了光合作用的CO2同化[31]; K还有助于植物组织的扩增生长,有利于植株生物量的积累。在本研究中,除165 m海拔处的立柳枝条中K元素含量有降低外,水淹未对立柳叶、枝条和根中K元素含量造成显著影响,说明库区内立柳在水淹胁迫条件下对该元素的积累和分配具有较好的适应,从而保证了植株株高、基径、冠幅等组织的扩增及正常的生理功能。
Ca和Mg元素是植物体内重要的中量元素。Ca能够感知和调节环境压力[14],通过维持细胞壁、细胞膜及膜蛋白的稳定性,以及通过参与信号传导来调节植物细胞对逆境反应和逆境适应性[32]。Mg是叶绿素的重要组成部分,在叶片中积累的最多,主要参与植物的光合作用[14],并涉及到大量光合酶的活化以及植物韧皮部形成[33]。本研究结果表明,165 m海拔处立柳叶和枝条中的Ca含量显著降低,但根中Ca元素的含量相对稳定。说明在库区消落带高强度的水淹胁迫降低了植株自身机能,导致对Ca离子的吸收能力下降,地上部分的积累量减少。立柳叶和枝条中Ca元素含量的降低可影响叶绿体双磷脂膜的形成,导致有机质积累量的持续减少,并影响到植物的正常生长,这也许是造成库区165 m海拔处立柳各项生长指标显著降低的原因之一。但立柳根中Ca元素含量的稳定,有助于对逆境胁迫信号的传递及并积极维持细胞膜形态,防止厌氧呼吸产生的有害物质对细胞膜的破坏。各处理组立柳Mg元素的含量和分配特征均表现为叶>根>枝条,但受到水淹影响,海拔165 m的立柳叶中的含量显著降低,这可能会影响到植物的正常光合作用,导致立柳生长所需的光合产物供应不足。因此165 m海拔的立柳需要追踪关注Mg含量降低对其生长造成的影响。
微量元素在植物体内的含量非常少,但对植物的生长却至关重要[34]。Fe和Mn是光合作用功能单元中的重要组分。本研究结果显示,水淹不同程度增加了立柳根部Fe和Mn的含量,但对立柳叶中的Fe和Mn的含量影响不明显,均处于正常范围内。立柳叶中Fe和Mn的含量的稳定是维持其正常的光合作用而采取的一种自我保护机制。但在水淹环境下,土壤氧化还原电位降低,Fe2+和Mn2+浓度增加,可溶性Fe2+和Mn2+很容易被植物根部过量吸收,导致立柳体内Fe、Mn的含量显著增加[35],尤其是165 m海拔处立柳的根和枝条。植物过量吸收Fe元素会对自身产生毒害作用,不仅会影响植物的光合作用,还会对植物呼吸酶结构造成破坏;同时立柳根部Fe元素的大量积累会在根际形成一层铁氧化合物保护膜[36],该保护膜会进一步限制立柳对其他营养元素的吸收利用,最终影响到植物的伸长生长,这可能是导致海拔165 m立柳各生长指标较170 m以上显著降低的又一个原因。
Cu和Zn是氧化还原酶的成分,在植物体内主要参与氧化还原反应。植物在水淹胁迫下会增加根部对Cu和Zn的吸收量,从而保证植物根际的强氧化还原能力,将低价阳离子氧化、阴离子还原,防止长期水淹导致低价阳离子产生的毒害[22]。本研究结果显示,水淹导致立柳根中Cu和Zn元素含量显著降低。立柳根中Cu和Zn元素含量降低会导致立柳Fe、Mn元素含量显著增加,而Fe元素含量增加又会限制植物对Cu和Zn的吸收。长期以往可能会影响到立柳正常的生理功能。本研究结果还表明,165 m海拔的立柳枝条中Cu、Zn含量显著低于170 m以上的立柳,这会导致165 m海拔处立柳的呼吸作用下降,同时由于生长激素的合成不足而影响植物的伸长生长[37],最终引起海拔165 m的立柳生长状况相交于其他两组差一些。
除此之外,库区冬季高强度水淹还会导致植物的生境发生改变,如土壤的酸碱度、水气交换情况、土壤微生物群和土壤元素含量等均会对植物的生长以及营养元素的吸收造成影响[11,38]。同时相关研究表明,在水淹胁迫下,植物根系代谢受阻会使根系对土壤中营养元素的主动吸收减弱,从而导致土壤中营养元素的可利用性与植物营养元素含量之间的关系变弱[39- 40]。目前一些学者对库区消落带内的立柳植株各营养元素与土壤中相对应的营养元素进行相关性分析后,发现这两者之间并无显著性相关关系[41],这说明了在水淹环境下,造成库区消落带内不同海拔高度的立柳生长及营养元素积累量之间存在差异更主要是由水淹胁迫导致的。
综上所述,在本研究中三个不同水淹强度下的立柳能主动通过调整其营养元素的积累和分配方式积极应对库区消落带周期性的水位变化。经过3个水淹周期后,165 m海拔处立柳营养元素的积累和分配方式与170 m海拔以上区域植株的差异较为明显,特别是Ca、Mg、Fe、Cu和Zn元素的差异对该海拔处立柳生长的影响最为显著。同时与170 m海拔以上区域的植株相比,经过3 a的冬季水淹后,165 m海拔处立柳的株高、基径和冠幅受到了明显抑制。所以持续的周期性水淹是否会进一步影响165 m海拔处植株叶、枝条和根中营养元素的含量与分配,进而影响立柳在该区域的生长乃至生存。
猜你喜欢
当代水产(2022年8期)2022-09-20
汽车实用技术(2022年16期)2022-09-03
湖南水利水电(2021年6期)2022-01-18
湖南水利水电(2021年6期)2022-01-18
农业工程技术(2021年28期)2021-12-30
小哥白尼(军事科学)(2018年3期)2018-06-15
现代园艺(2017年23期)2018-01-18
大众考古(2015年7期)2015-06-26
天然气勘探与开发(2012年2期)2012-03-25
植物营养与肥料学报(2011年3期)2011-10-24
