
模拟短期降雨量变化对准噶尔荒漠植物群落的影响
2019-09-05 08:57徐文轩杨维康
水土保持研究 2019年5期
吴 珂, 徐文轩, 杨维康
(1.中国科学院 新疆生态与地理研究所, 中国科学院干旱区生物地理与生物资源重点实验室, 乌鲁木齐 830011; 2.中国科学院大学, 北京 100049)
降雨是影响干旱、半干旱地区植被形成和发展的主要限制因子[1]。过去50 a里新疆各地降水量总体呈上升趋势,其中近10 a来新疆北部降水量变化幅度与增加趋势尤其明显[2]。有研究认为未来100年,新疆是中国干旱区降水量增加最快的地区[3],极端降水量也呈增加趋势[4-5]。亦有不同观点认为未来新疆北部将进入一个多年降水偏少的时期[6]。由此,新疆北部未来的降雨量变化特征,及其对荒漠植物群落的影响逐渐引起学者的广泛关注。植物生长季降雨量的变化是影响干旱、半干旱地区草地植被覆盖度和高度的主要驱动力[7-8],特别是在草地植被返青后迅速生长至成熟这段时间,植被对降雨量变化最为敏感[9-10]。因此无论未来降水增加还是减少,降雨量的改变都将对该区域的植物群落产生一系列影响。
降雨量变化可影响植物生长、改变物种间关系,进而影响植物群落的组成和结构,最终影响生态系统功能[11-12]。此外降雨量变化能够改变土壤水分含量、土壤微生物种类及组成等,从而对陆地生态系统生态过程产生间接影响[13]。近年来,我国关于草地生态系统如何响应全球变化的研究发展迅猛,取得了一系列的研究成果[14-15]。有学者认为降雨量变化对植物群落无显著影响。例如陈美玲等[16]在内蒙古图牧吉自然保护区对贝加尔针茅草甸的研究表明,增水处理虽然对土壤pH值、全磷含量和碳氮比有极显著的影响,但对植物群落关键指标无显著影响。而另一些学者则持相反观点,认为大量的降雨将增加氮流失、降低氮素滞留时间,从而加剧植物对土壤水分和养分的竞争,导致植物群落结构发生改变。例如在青海北部的高寒矮嵩草草甸和内蒙古短花针茅荒漠草原开展的模拟降雨研究均表明,随降雨增加不同植物类群的地上生物量、物种多样性指数和均匀性指数均有一定程度的提高[17-18];在黄土高原半干旱区的研究表明,陆地生态系统生产力的变化与降雨量的波动紧密相关,且降水越多,植物群落初级生产力越高;在加利福尼亚研究也发现,随降雨量增加一年生草地净初级生产力逐渐升高[19]。目前关于降雨量变化对草地植物群落影响的研究主要集中在草原和草甸地区,荒漠地区开展的相关研究少见报道。
新疆幅员辽阔,面积约为166万km2,占中国陆地面积的1/6,其中荒漠草地类占全新疆草地面积的46.93%,占新疆土地总面积的16.3%[20]。新疆有40%的荒漠可为放牧利用,是新疆游牧家畜的春秋季牧场及冬季牧场[21]。受自然和人为因素影响,新疆北部准噶尔荒漠草地大面积退化,严重威胁当地牧民的可持续牧业生产[22]。因此开展准噶尔荒漠植物群落对未来降雨量变化的响应机制,不仅有助于揭示气候变化对荒漠生态系统的影响,也有助于准确评价荒漠生态系统各项健康指标并预测其演变趋势,对制定合理可行的荒漠草地可持续管理和经营措施具有重要理论与实践意义。
1 材料与方法
1.1 研究区概况
研究区位于新疆准噶尔盆地东南缘、木垒县城东北部约80 km处的中国科学院木垒野生动物生态监测试验站(44.04°N,90.99°E)。试验站所在区域南邻天山,北接古尔班通古特沙漠,为开阔冲积平原,地势平坦,平均海拔800~1 000 m。土壤类型为壤质荒漠灰钙土,局部地区砾质性较强,北部边缘靠近沙漠为沙质土壤[23]。研究区属温带大陆性干旱气候区,春夏季多大风天气,年均温5.1℃,年平均降水量187.6 mm,主要集中在冬季和春季。无稳定地表径流,仅在大雨过后能在沟壑和洼地中形成临时性积水。
研究区植物群落物种组成相对简单,植被低矮稀疏,高约10~15 cm,盖度一般在20%以下。博洛塔绢蒿(Seriphidiumborotalense)群落是该区域的地带性植被类型,其他常见的灌木有梭梭(Haloxylonammodendron)、假木贼(Anabasissalsa)、刺木蓼(Atraphaxisspinosa)和琵琶柴(Reaumuriasongarica);多年生草本植物有绢蒿、沙生针茅(Stipaglareosa)等;一年生草本植物有四齿芥(Tetracmequadricornis)、小车前(Plantagominuta)、角果藜(Ceratocarpusarenarius)和叉毛蓬(Petrosimoniasibirica)等;此外黑鳞顶冰花(Gageanigra)、荒地阿魏(Ferulasyreitschikowii)、鸦葱(Scorzonerasp)、独行菜(Lepidiumsp)和郁金香(Tulipasp)等20余种短命和类短命植物分布于此。研究区为典型的春秋过渡牧场,每年3月初至6月上旬、9月初至11月中旬,畜群(主要是羊群)在冬夏牧场往返时停留于此,其余时间研究区仅散放有骆驼,且几乎无人类活动[24]。
1.2 研究方法
1.2.1 试验设计 试验于2016年4月—2017年9月进行。试验开始前,选择一处地势平坦、封育3 a的禁牧围栏作为研究样地。通过测定不同样地群落多样性、盖度和生物量指标(表1),发现研究区各指标在不同样地间无显著差异,说明试验区域选择合理,样地植被类型较为均一,土壤类型相同。试验共设置减雨60%,减雨30%,自然对照,增雨30%,增雨60%5种处理方式,每种处理的样地面积2 m×2 m。每个样方旁边设置至少3 m的缓冲带,消除处理样方间的互相干扰和试验过程中人为践踏的扰动。试验共设置5个重复,共计25个样方。分析近50 a来研究区降雨量数据,表明研究区降雨主要集中在春季(4—5月),且降雨量年际变化为±30%左右,因此本试验中将模拟降雨量变化梯度设置为增减30%。同时选取增减60%的降雨作为极端状况进行模拟[25]。
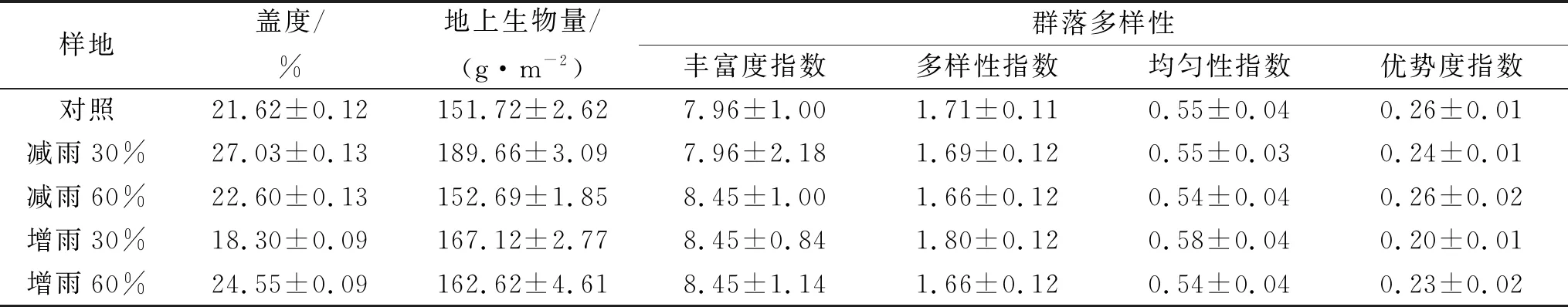
表1 试验前不同样地盖度、地上生物量和群落多样性
1.2.2 降雨处理 降雨减少参考采用固定式遮雨棚法[26]。遮雨棚的材料选用透光率超过90%的聚碳酸酯板。聚碳酸酯板被做成V字型板条,呈栅格状排列在铁制框架上,倾斜度为15°。聚酯碳酸板遮挡的面积比例即为降雨减少比例,通过控制V型板条的数量达到减雨30%和减雨60%的目的。遮雨棚的高度最低为50 cm,以保证近地表空气流通。为避免边缘效应,每个遮雨样地面积为3 m×3 m。每个样方旁挖取壕沟,以阻止坡面径流对截雨效果的影响。每次降雨结束,立即将减雨样地内收集到的雨水储存起来,并添加到增加降雨的样地内。添加降雨时,将所需添加的水分均匀的浇洒在试验小区表面(图1)。

图1 降雨量减少30%(A)和减少60%(B)
1.2.3 植物群落特征调查与生物量测定 植物群落特征调查:研究区降雨主要集中在春季的4月初-5月底,是短命植物生长最旺盛的时期,物种丰富度最高。于2017年5月在每个样地内调查植物种类、数量、盖度、高度等指标。在2017年8月中旬,植物地上生物量达到最高峰时采集植物地上部分,将样方内植物分种齐地面剪下后分装纸袋,即时称鲜重,带回实验室置入烘箱内,在80 ℃温度下烘12 h 后称干重以测定生物量。
1.2.4 数据分析计算 物种重要值采用如下公式:
IV=(RC+RH+RD)×100/3
RC=(某种植物的盖度/全部植物的盖度和)×100%
RH=((某种植物的高度/全部植物的高度和)×100%
RD=(某种植物的个体数/全部植物的个体数)×100%
式中:IV为重要值;RC为相对盖度;RH为相对高度;RD为相对密度[27]。
群落多样性的测度选用Simpson优势度指数、丰富度指数、Shannon-Wiener指数和Pielou均匀度指数。
物种丰富度指数(R):R=S
Shannon-Wiener指数(H):H=-∑(PilnPi)
Pielou均匀度指数(E):E=H/lnS
式中:S为样地植物种总数;i代表第i个物种;N为群落中所有物种的个体数之和;Pi为种i的重要值[28]。
1.2.5 统计分析 原始数据的整理采用Microsoft Excel 2013 来完成。数据分析利用SPSS 20.0软件。首先采用Kolmogorov-Smirnov检验数据是否符合正态分布。对于符合正态分布的数据采用单因素方差分析(One-way ANOVA)和多重比较分析比较不同降雨处理间α-多样性、盖度和地上生物量的差异显著性,用Pearson相关性分析比较不同处理与植物群落指标间的相关性。除特殊说明,所有统计检验的显著水平为p<0.05,数据均为平均值±标准误。
2 结果与分析
2.1 植物群落的物种组成与重要值
重要值是表示某物种在群落中地位和作用的综合数量指标。样地内共记录到10科19种植物,其中藜科植物7种,十字花科、菊科和禾本科植物各2 种,紫草科、车前科、柽柳科、白花丹科、旋花科和蓼科植物仅各有1种(表2)。依据生活型划分,样地中共有灌木6种,多年生草本植物3种,一年生草本植物10种。比较不同降雨处理梯度下植物的重要值,发现以假木贼(Anabasissalsa)、驼绒藜(Ceratoideslatens)和小蓬(Nanophytonerinaceum)为代表的藜科植物在植物群落中占优势地位(表2)。与对照样地相比,减雨样地物种数量较少,假木贼的重要值明显增加,在减雨60%时达到最高;优势度较高的其他植物分别是离子芥(Chorisporatenella)、驼绒藜和小车前(Plantagominuta);增雨样地物种数量高于对照样地,假木贼重要值虽有所增加,但仍显著低于减雨样地,多年生草本植物绢蒿(Seriphidiumborotalense)的优势度明显提高。
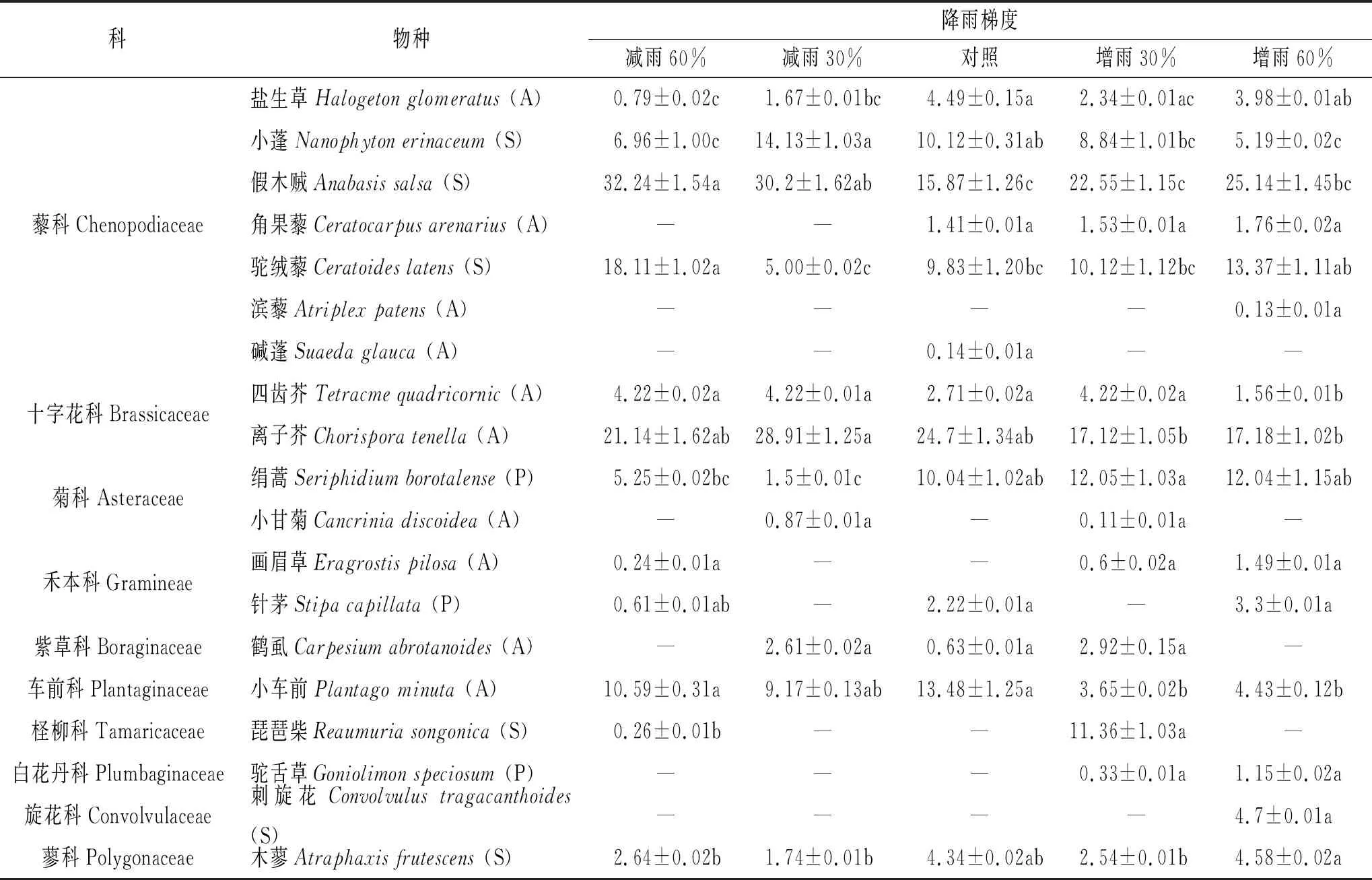
表2 不同降雨梯度下的植物群落的种类组成及其重要值 %
注:“—”表示未找到该物种;“A”表示一年生草本植物,“P”表示多年生草本植物,“S”表示灌木;同一列同一植物不同小写字母间差异显著,p<0.05。
2.2 植物群落物种多样性分析
试验开始前(2016年),不同样地植物物种丰富度无显著差异,不同处理样地的各指标较为均一;试验进行1 a后(2017年),植物物种丰富度在不同降雨增加处理间发生了显著变化(F=4.308,p<0.05)。随着降雨量增加呈一定的上升趋势(r=0.648,p>0.05)。与对照样地相比,降雨增加后,植物物种数增加,增雨60%的样地内植物物种丰富度显著高于对照样地和减雨样地,而增雨30%与增雨60%样地的丰富度指数无显著差异;而在降雨减少的样地内,植物物种丰富度指数略低于对照样地但无显著差异。无论降雨增加还是减少,群落的shannon-wiener多样性指数、Pielou均匀度指数和simpson优势度指数均无显著差异(p>0.05)(表3),主要原因是荒漠区植被种类稀少,且数量较少。

表3 不同降雨梯度样地植物群落α物种多样性的变化
注:数据后不同字母表示同行内样地间差异显著(p<0.05)。
2.3 降雨量变化对植物群落盖度和地上生物量的影响
试验开始前,不同生活型植物的盖度和生物量在不同样地间无明显差异。试验进行到第二年(2017年),不同处理样地间植物群落盖度与降雨量呈明显的正相关关系(r=0.993,p<0.01)。但仅增雨60%的样地与减雨60%样地间存在显著差异(F=0.835,p<0.05),其中灌木盖度和多年生草本植物的盖度与降雨量显著相关(灌木:r=0.869,p<0.05;多年生草本:r=0.949,p<0.05),一年生草本的盖度与降雨量变化无明显相关性(r=0.222,p>0.05),见图2。
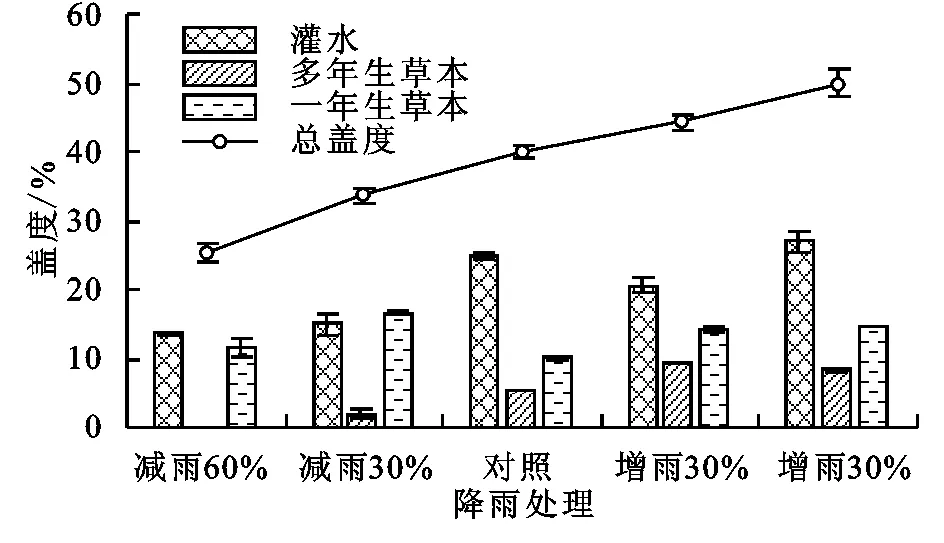
图2 不同降雨处理样地的植被盖度
随降雨量增加,不同处理样地植物群落地上生物量先增加后减小,增雨30%时明显增加且达到最大值(296.38 g/m2)。其中,多年生草本植物表现出与总地上生物量相同的变化趋势,在增雨30%时地上生物量达到峰值(113.17 g/m2),但不同降雨处理间无显著差异;灌木生物量随降雨量增加而增加,且在增雨60%时达到最大值;一年生草本植物的地上生物量没有随降雨量变化表现出明显的变化规律(图3)。灌木的地上生物量和总地上生物量与降雨量呈极显著的正相关关系(r=0.978,p<0.01;r=0.993,p<0.01),多年生草本植物的生物量与降雨量显著相关(r=0.598,p<0.05),而一年生草本植物的地上生物量与降雨量无相关性(r=0.109,p>0.05)。说明为期1 a的增减雨试验对荒漠植物群落的盖度和生产力具有显著影响,特别是对灌木和多年生草本植物影响显著,而对一年生植物的盖度和生产力影响较小。
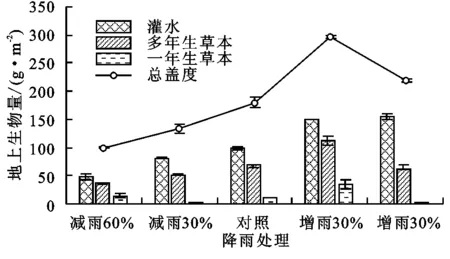
图3 不同降雨处理样地地上生物量的变化
3 讨论与结论
全球气候变化必然会导致区域降雨格局改变[29],准噶尔荒漠地处中亚内陆干旱区,多数学者认为该区域未来降水量将呈增加趋势[3-4,30],也有少数观点认为该区域存在降水偏少的可能[6]。因此,在该区域未来降水格局难以确定的前提下,通过控制试验,开展准噶尔荒漠区域未来降雨变化的生态效应研究尤为迫切。已有研究表明,人为追加和减少雨量可以使该地区荒漠植物个体、群落发生动态变化。例如降雨量增加能够促进荒漠草地植物生长,提高群落物种丰富度[13,31];降水减少会降低土壤含水量,进而抑制植物生长,导致植被盖度降低、物种丰富度下降等[31-32]。本研究表明,在植物生长季短期的降雨量变化也能够对荒漠草地植物群落结构、物种多样性和地上生物量等产生不同程度的影响,且不同生活型植物对生长季降雨量变化的响应各不相同。随降雨量增加,样地内主要优势物种重要值降低(表2),样地内物种数增加,且增加的物种主要是一年生草本植物。这符合西北干旱区植物的生长特性,降水丰富时短命植物开始增加,致使群落物种数量增加[33]。这是因为以短命植物为主的一年生草本植物对水分利用效率较高,对水分的供应极为敏感。可见,降雨量变化对荒漠植物群落物种丰富度起到至关重要的作用,特别是对降雨量变化更为敏感的一年生植物。这也与陶冶等[34]在新疆北部荒漠的研究结果一致。
物种多样性是物种丰富度和物种均匀度结合起来的一个统计量[35],Shannon-wiener 指数可以很好的反映出群落的物种多样性程度,优势度和均匀度与物种多样性密切相关[36]。均匀度指数与物种多样性指数呈正相关,与优势度指数呈负相关,优势度指数越高,说明群落内物种数量分布越不均匀,优势种的地位越突出。干旱区物种相对稀少,且物种个体大小和数量差异较大,通过多个指数的比较更能反映出群落的真实状况。多数研究表明降雨量增加能在一定程度上提高群落物种多样性,进而引起植物群落结构发生变化[37-38]。然而本研究发现,短期降雨量的改变并未显著影响荒漠植物群落物种多样性,但提高了荒漠植物群落物种丰富度,特别是一年生草本植物的丰富度。这可能是因为:(1) 荒漠植物群落种类和数量相对稀少、盖度低,群落物种组成相似程度较高,因此不同处理样方的多样性指数具有较强的一致性;(2) 荒漠植物群落中大多数物种的绝对数量较少,虽然对群落多样性起到了一定的作用,但对综合多样性指数贡献不大[35]。许多研究表明浅根系的一年生植物种子在降水丰富的年份能迅速萌发并生长[39-40]。然而,我们的研究表明一年生草本植物的盖度和生物量并未出现显著差异。Noy-Meir[8]认为在干旱、半干旱区,是“生物学上重要的”或“有效”的降水激活了植物的生长和繁育。在研究区,大多数一年生草本植物为短命植物,在夏季干热季节来临之前短短的2 个月左右时间即可迅速完成生活史周期[41-42]。这些物种由于生长期较早春季融雪能够有效促进其生长,在5月底快速完成生活史后枯萎或休眠,因此之后的降雨对其生物量积累不构成影响[43]。作者在8月测定盖度和生物量时,上述一年生短命植物早已经枯萎消失,因此生长季降雨量的变化虽然改变了以短命植物为主的一年生植物物种丰富度,但其对于盖度和生物量的影响并不显著。
植被盖度是衡量天然草地生产力高低、表征草地生长状况的一个重要参数[44-45]。本研究发现,随降雨量增加植物群落盖度逐渐增加,其中灌木盖度变化趋势与降雨量一致,这与多数研究结果吻合[46-47]。研究区属于春秋过渡牧场,植物群落物种数量少、结构简单,假木贼、绢蒿群系是该地区典型的地带性植被,研究其对降雨量变化的响应,有助于预测未来降雨量改变对该类型植被动态和生产力的潜在影响。研究发现,假木贼在群落中的优势度随降雨量减少而显著增加,绢蒿的优势度随降雨的增加而增加。在研究区,绢蒿春季刚萌发的嫩枝和秋季霜打后的植株为家畜所喜食,而假木贼因适口性差很少被家畜采食[48]。因此未来如果降水增加可能促进新疆北部荒漠草地的建群种优势度发生变化,提高荒漠草地作为春秋过渡牧场的利用价值,降水减少则可能降低荒漠草地的利用价值。
本研究中,增雨60%反而导致群落地上生物量下降,虽然仍然高于对照和减雨样地,但低于增雨30%的样地。作者分析认为这与研究区植被为典型的荒漠植被,适当的增雨能够提高荒漠植物的生产力,但60%的增雨对荒漠植物来说而言并不是可以利用的有效降雨量,特别是深根型的灌木[49]。其次,降雨量增加60%可能超过了多年生草本植物适宜的水分范围,导致适应干旱胁迫的多年生草本植物出现了不适应现象[50],从而导致群落总生物量的降低(图3)。此外,这也可能与试验开展的年限较短有关,这需要在今后的试验中进一步验证和分析。
在全球气候变化背景下,降水量也会发生不同程度的变化。荒漠植被对降水量变化的响应非常敏感[51]。降雨量增加会提高荒漠草地植物群落物种丰富度、植物的盖度以及灌木和多年生草本植物的地上生物量,但对一年生植物的盖度和地上生物量无显著影响。当然,荒漠草地内植被的生长除了受到降雨的影响外,还受到温度、氮沉降等其他因素的影响,这需要我们在今后的研究中同时予以关注和重视。
猜你喜欢
小哥白尼(趣味科学)(2022年5期)2022-08-15
治淮(2022年4期)2022-01-01
海河水利(2021年4期)2021-08-30
数学小灵通·3-4年级(2021年6期)2021-07-16
南方农业·中旬(2020年9期)2020-11-30
黑龙江工程学院学报(2020年5期)2020-10-21
绿色中国(2019年14期)2019-11-26
绿色科技(2017年15期)2017-09-01
儿童故事画报·智力大王(2016年7期)2017-02-08
文史春秋(2016年3期)2016-12-01
