
黏虫CYP9A113基因的克隆及外源物质对基因表达的诱导效应
2019-09-04 09:34张雅男刘月庆JUNAIDSM王智琪樊东
植物保护 2019年4期
关键词:克隆
张雅男 刘月庆 JUNAID S M 王智琪 樊东


摘要 黏虫是我国作物上最重要的害虫之一。细胞色素P450能够参与昆虫外源物质代谢。本研究采用RACE技术克隆了一条编码黏虫P450基因的cDNA序列,并通过Real-time PCR技术,检测了4种外源物质对该基因表达的诱导效应。该基因被国际P450命名委员会命名为CYP9A113,GenBank登录号为KY436739。利用2.5%高效氯氟氰菊酯乳油的LD50处理黏虫3 h,LD10、LD30和LD50处理12 h和24 h,可诱导表达CYP9A113基因;20%氯虫苯甲酰胺悬浮剂的LD10处理黏虫12、24和48 h,LD30和LD50处理24 h,CYP9A113基因表达呈诱导效应;0.1和0.5 mg/mL香豆素处理6、12、24和48 h,CYP9A113基因表达均呈诱导效应;0.1和0.5 mg/mL吲哚-3-甲醇处理3、6、12、24和48 h,CYP9A113基因表达均呈诱导效应。
关键词 黏虫; CYP9A113基因; 克隆; 外源物质; 诱导效应
中图分类号: S 435.132
文献标识码: A
DOI: 10.16688/j.zwbh.2018358
细胞色素P450酶(P450s或CYPs)是一类亚铁血红素硫醇盐蛋白[1],在哺乳动物的肝微粒体中首次被发现[2],现已证明几乎在所有生物体内广泛存在。自1965年Ray在昆虫体内发现P450后[3],目前已经克隆得到分属于CYP2、CYP3、CYP4和线粒体P450s的多个昆虫细胞色素P450基因序列[4],其中CYP3分支又包括CYP6和CYP9家族[5],这些基因既能催化激素、信息素、脂肪酸等内源物质的合成,又能参与杀虫剂、诱变剂、植物次生代谢物质等外源物质的代谢,对昆虫的生长发育起着非常重要的作用[68]。采用98%氯氰菊酯(55 ng/μL)对野桑蚕Bombyx mandarina点滴1 μL处理后,脂肪体中CYP9A20和CYP9A21基因表达量均升高[9];甘蓝夜蛾Mamestra brassicae经0.5 μL 2.5%溴氰菊酯水乳剂(0.152 ng/g)处理后,CYP9A90基因表达量总体呈诱导趋势[10];甜菜夜蛾Spodoptera exigua经95%氯虫苯甲酰胺(0.01 mg/kg和0.02 mg/kg)处理后,CYP9A9基因相对表达量升高[11];香豆素可诱导斜纹夜蛾Spodoptera litura脂肪体中CYP4M14和CYP6AB14基因的表达[1213]。吲哚-3-甲醇处理可诱导飞蝗Locusta migratoria体内CYP6HL1和CYP6FE12基因的表达[14]。
黏虫Mythimna separata属于鳞翅目,夜蛾科,是粮食作物上的重要害虫之一[15],在多个国家广泛分布,我国除西藏地区未见报道外,其他各地均有报道[16]。本研究克隆黏虫CYP9家族的一条P450基因,选取生产上常用于防治黏虫的杀虫剂2.5%高效氯氟氰菊酯乳油、20%氯虫苯甲酰胺悬浮剂和黏虫豆科寄主植物中含有的次生代谢物质香豆素[17]、十字花科蔬菜活性成分吲哚-3-甲醇[18],对其进行诱导效应的研究,探究该基因在黏虫外源物质解毒和对自然环境的适应性方面的重要作用,为利用分子手段防治该害虫奠定基础。
1 材料与方法
1.1 供试昆虫
黏虫M.separata采自东北农业大学向阳实验基地,黑光灯诱集成虫后于实验室以5%的蜂蜜水饲养,幼虫用玉米叶饲养,均放入恒温培养箱中,温度为(25±1)℃,相对湿度为70%,光周期为L∥D=16 h∥8 h。在实验室饲养2代后用作试验材料。
1.2 供试试剂与药剂
TRIzol Reagent试剂购自Invitrogen公司;反转录试剂盒、低熔点琼脂糖购于Promega公司;3′RACE试剂盒、5′RACE试剂盒、DNA纯化试剂盒和Taq DNA聚合酶购于TaKaRa公司;ReverTra Ace qPCR RT Master Mix with gDNA Remover、THUNDERBIRD SYBR qPCR Mix購于TOYOBO公司;2.5%高效氯氟氰菊酯乳油(EC)购于青岛星牌作物科技有限公司,20%氯虫苯甲酰胺悬浮剂(SC)购于美国杜邦公司;香豆素(coumarin)和吲哚-3-甲醇(indole-3-carbinol)购于合肥博美生物科技有限公司;其余试剂为国产或进口的分析纯。由上海生工生物有限公司完成基因的测序和引物合成。
1.3 CYP9A113基因序列的克隆及分析
1.3.1 总RNA的提取以及cDNA的合成
选择健康的4龄黏虫幼虫为样本,采用TRIzol提取法提取总RNA,按照反转录试剂盒的说明书合成cDNA,并保存于-20℃冰箱,剩余的RNA保存于-80℃冰箱备用。
1.3.2 全长基因序列的克隆
将GenBank上已经登录的甘蓝夜蛾M.brassicae CYP9A90(KR676343)和棉铃虫Helicoverpa armigera CYP9A12(AY371318)基因的序列进行比对,设计黏虫保守区上下游特异性引物Ms9A-F和Ms9A-R(表1)。以1.3.1中合成的cDNA为模板进行PCR扩增,将得到的PCR产物在1%琼脂糖凝胶电泳上检测,目的片段胶回收测序。根据3′RACE和5′RACE试剂盒中自带的引物3′RACE-Ro、3′RACE-Ri和5′RACE-Ro、5′RACE-Ri,分别设计对应的嵌套引物Ms9A-3′RACE-Ro、Ms9A-3′RACE-Ri和Ms9A-5′RACE-Ro、Ms9A-5′RACE-Ri(表1)。按照试剂盒说明书,分别进行3′端序列和5′端序列的扩增,并进行胶回收测序。通过AlignX软件将保守区序列、3′端序列和5′端序列进行拼接,得到完整的基因序列。再设计全长引物Ms9A-Full-F和Ms9A-Full-R(表1)克隆全长序列进行验证,确定最终序列。
1.3.3 序列分析
该基因由国际P450命名委员会命名,利用NCBI中的Open Reading Frame Finder (ORF Finder)将基因开放阅读框翻译成氨基酸序列;利用在线網站ExPASy (https:∥www.expasy.org/)进行功能域的预测;利用BioXM 2.6软件进行蛋白质分子量和等电点的计算;利用MEGA 5.1软件采用邻接(neighbor-joining)法构建系统进化树。
1.4 杀虫剂的诱导效应
1.4.1 高效氯氟氰菊酯的诱导效应
选取4龄第1天黏虫幼虫,称量单个虫体的重量,计算平均值,将2.5%高效氯氟氰菊酯EC用丙酮稀释成5个不同浓度,分别为33.33、16.67、8.33、4.17、2.08 μg/mL,以丙酮为对照。每个浓度药剂和对照均点滴0.5 μL于虫体的第2~3腹节之间,每处理20头,3次重复,于培养箱中正常饲养条件培养。24 h后检查死亡虫数,求出2.5%高效氯氟氰菊酯EC对黏虫的LD10、LD30和LD50。
选择4龄第1天黏虫幼虫进行试验,分别点滴0.5 μL LD10、LD30和LD50的2.5%高效氯氟氰菊酯EC于黏虫幼虫2~3腹节之间,每处理和对照均点滴40头,共设3个重复,放在实验室饲养条件下培养,于3、6、12、24和48 h分别收集3头存活的幼虫,于液氮中速冻,放在-80℃冰箱中保存。每个重复的3头幼虫混合提取RNA后,根据荧光定量反转录试剂盒制备cDNA模板。采用SYBR Green染料3步法进行荧光定量,利用Primer 5.0软件设计荧光定量上下游引物Ms9A-RT-F和Ms9A-RT-R(表1),以黏虫β-actin作为内参基因,设计上下游引物β-actin-F和β-actin-R(表1)。在荧光定量PCR仪(BIO-RAD CFX Connect)中进行反应,反应结束记录熔解曲线和相关数据,检测LD10、LD30和LD50的2.5%高效氯氟氰菊酯EC处理不同时间CYP9A113基因的表达情况。
1.4.2 氯虫苯甲酰胺的诱导效应
选择4龄第1天黏虫幼虫进行试验,饥饿处理12 h后利用ddH2O将20%氯虫苯甲酰胺SC稀释成5个不同浓度,分别为5、2.5、1.25、0.625和0.312 5 μg/mL,以ddH2O为对照,每个浓度20头黏虫,3个重复。将玉米叶用打孔器打成直径为1 cm的叶碟,其上点10 μL药液晾干备用,每个培养皿中均放入1头黏虫和1片叶碟,叶碟吃光后用未处理正常叶片喂饲。24 h后检查死亡虫数,求出20%氯虫苯甲酰胺SC对黏虫幼虫的LD10、LD30和LD50。
供试虫体的选择和试验方法同上,分别用LD10、LD30和LD50进行处理,以ddH2O作为对照,每处理和对照均饲喂40头,设3个重复,于3、6、12、24和48 h分别收集3头存活的幼虫,于液氮中速冻,放在-80℃冰箱中保存。提取RNA后,通过Real-time PCR检测LD10、LD30和LD50的20%氯虫苯甲酰胺SC处理不同时间CYP9A113基因的表达情况,具体步骤参照1.4.1。
1.5 植物次生代谢物质的诱导效应
选取4龄第1天幼虫进行试验,香豆素和吲哚-3-甲醇均设两个浓度,分别为0.1和0.5 mg/mL,用加热的无菌ddH2O稀释到所需浓度,对照为加热的无菌ddH2O。待溶液温度降到室温时,开始试验,之后的试验方法均参照1.4.2。提取RNA后,通过Real-time PCR检测不同浓度的2种植物次生代谢物质处理不同时间CYP9A113基因的表达情况,具体步骤参照1.4.1。
1.6 数据处理与分析
利用DPS软件进行回归统计分析;采用2-ΔΔCt法计算荧光定量PCR相对定量数据结果;数据的多重比较和差异显著性分析利用SAS 9.4软件Duncan氏检验法进行(P < 0.05);数值均以平均值±标准误(SE)表示;图表的制作以及相关数据的计算均在Excel 2010中进行;将每一时间点CK均设为1,同一时间点进行差异显著性分析作图。
2 结果与分析
2.1 CYP9A113基因的鉴定及序列分析
通过克隆测序后拼接,进行序列全长的验证后,得到一条包含1 752个碱基的P450基因序列,在NCBI上比对发现其属于昆虫CYP9A亚家族,经国际P450命名委员会命名为CYP9A113(GenBank登录号:KY436739)。CYP9A113基因编码529个氨基酸,分子量约为60.4 kDa,等电点为8.81。在编码的氨基酸序列中,具有P450基因的保守序列,分别为血红素结合域(FGVGPRNCIG)、meander结合域(PEKFDPERF)、螺旋C(WKDMR)、螺旋I(AGFETV)和螺旋K(ELLR)(图1)。
翻译后的氨基酸在NCBI的BLAST上比对,找出与之相似的其他鳞翅目昆虫的氨基酸序列,进行系统进化树的构建。结果发现,克隆的黏虫M.separata CYP9A113(ARI68319)与甘蓝夜蛾M.brassicae CYP9A13(AAR26518)、棉铃虫H.armigera CYP9A17(AAY21809)和美洲棉铃虫H.zea CYP9A12 (ABH09252)首先聚类,然后与斜纹夜蛾S.litura CYP9A39(ACV88722)和甜菜夜蛾S.exigua CYP9A9(BAG71410)聚类,再与草地贪夜蛾Spodoptera frugiperda CYP9A58(AID55429)聚类,与六星灯蛾Zygaena filipendulae CYP9A36(ACZ97417)和家蚕Bombyx mori CYP9A19(BAM73827)等其他鳞翅目非夜蛾科昆虫CYP9家族基因遗传距离较远,聚类结果基本符合形态分类特征(图2)。
2.2 杀虫剂对CYP9A113基因表达的诱导效应
2.2.1 高效氯氟氰菊酯对CYP9A113基因表达的诱导效应
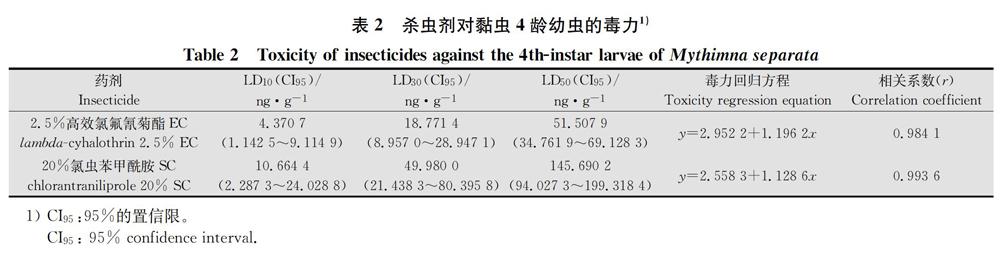
经过室内毒力测定,2.5%高效氯氟氰菊酯EC对黏虫4龄幼虫的LD10、LD30和LD50分別为4.370 7(1.142 5~9.114 9)、18.771 4(8.957 0~28.947 1)和51.507 9(34.761 9~69.128 3)ng/g(表2)。
黏虫幼虫经LD10和LD30的2.5%高效氯氟氰菊酯EC处理3 h,CYP9A113基因表达均被抑制,经LD50处理3 h,CYP9A113基因相对表达量升高,呈现诱导效应;LD10、LD30和LD50处理6 h,CYP9A113基因表达均被抑制;LD10、LD30和LD50处理12 h,CYP9A113基因表达均呈诱导效应,相对表达量分别为对照组的2.5、2.0和2.1倍;LD10、LD30和LD50处理24 h,CYP9A113基因表达均呈诱导效应,相对表达量分别为对照组的2.6、1.9和2.3倍;LD10和LD50处理48 h,CYP9A113基因相对表达量无明显变化,LD30处理下,CYP9A113基因表达被抑制(图3)。结果说明,2.5%高效氯氟氰菊酯EC对CYP9A113基因表达产生诱导效应与时间和剂量有关。
2.2.2 氯虫苯甲酰胺对CYP9A113基因表达的诱导效应
经过室内毒力测定,20%氯虫苯甲酰胺SC对黏虫4龄幼虫的LD10、LD30和LD50分别为10.664 4(2.287 3~24.028 8)、49.980 0(21.438 3~80.395 8)和145.690 2(94.027 3~199.318 4)ng/g(表2)。
黏虫幼虫经LD10、LD30和LD50的20%氯虫苯甲酰胺SC处理3 h,CYP9A113基因相对表达量无明显变化;3个剂量处理6 h,CYP9A113基因相对表达量均明显降低;LD10处理12 h,CYP9A113基因表达呈诱导效应,为对照组的3.9倍,LD30和LD50处理下无明显变化;LD10、LD30和LD50处理24 h,均对CYP9A113基因表达有诱导效应,分别为对照组的11.1、5.1和3.0倍;LD10处理48 h,CYP9A113基因表达呈诱导效应,为对照组的2.1倍,LD30和LD50处理下与对照相比无显著差异(图4)。
2.3 植物次生代谢物质对CYP9A113基因表达的诱导效应
黏虫幼虫经0.1和0.5 mg/mL香豆素处理3 h,CYP9A113基因表达均被抑制;处理6、12、24和48 h,均对CYP9A113基因的表达有诱导效应,且均在处理6 h诱导作用最强,分别为对照组的2.7和2.8倍;高浓度处理12、24和48 h,CYP9A113基因表达量均显著高于低浓度处理(图5)。结果说明,不同浓度香豆素处理6 h后,可能对CYP9A113基因表达均产生诱导效应,并且在12 h后,高浓度处理诱导效应高于低浓度处理。
黏虫幼虫经0.1和0.5 mg/mL的吲哚-3-甲醇处理3、6、12、24和48 h,CYP9A113基因表达均呈诱导效应,低浓度处理48 h诱导作用最大,为对照组的2.0倍。高浓度处理12 h诱导作用最大,为对照组的2.4倍(图6)。结果说明,不同浓度吲哚-3-甲醇处理不同时间,可能对CYP9A113基因表达均产生诱导效应。
3 结论与讨论
细胞色素P450在合成具有关键生物学功能的内源物质和代谢天然或合成的外源化学物质方面起重要作用[6],长期以来一直是研究的热点。本研究克隆黏虫的一条P450基因CYP9A113,属于在异物代谢和杀虫剂抗性中起重要作用的CYP9家族基因,研究了其在合成杀虫剂和植物次生代谢物质诱导后的表达情况,结果发现外源物质对其有诱导效应,而且是在一定剂量下经过一定时间才产生,具有时间和剂量效应。
已有研究表明,家蚕B.mori经1 μL的98%氯氰菊酯(5 ng/μL)处理24 h,脂肪体中CYP9A22基因相对表达量升高[19]。飞蝗L.migratoria用3 μL溴氰菊酯(0.01、0.02、0.04、0.08和0.12 μg/mL)处理12 h,在3个较高浓度处理下CYP9AQ2基因表达量升高,存在剂量效应[20]。甜菜夜蛾S.exigua经0.01 mg/kg的95%氯虫苯甲酰胺原药处理36 h,CYP9A9基因表达诱导效应最高,0.02 mg/kg处理12 h诱导效应最高,存在时间和剂量效应[11]。不同药剂对不同昆虫的同一家族P450基因产生的效应也可能不同。在本研究中,黏虫经LD10和LD30的2.5%高效氯氟氰菊酯EC处理12 h和24 h,对CYP9A113基因表达有诱导作用,LD50处理3、12和24 h均可产生诱导效应,说明基因的诱导反应与时间和剂量有关。LD50处理6 h,基因表达量降低,可能是由于虫体特殊生理过程对其造成影响,3个剂量处理48 h均无诱导反应,可能因为细胞色素P450酶系的合成是个耗能过程,不会一直保持高含量,当需要时产生诱导作用,药剂毒性逐渐消失,诱导作用也会慢慢消失。黏虫经LD10的20%氯虫苯甲酰胺SC处理12、24和48 h,CYP9A113基因表达呈诱导效应,LD30和LD50处理24 h产生诱导效应,同样具有时间和剂量效应,在开始的时间点没有诱导效应产生,并且剂量高,产生诱导效应时间晚,可能是由于虫体产生中毒反应,无法及时进行调节。
植物产生有毒物质防止被侵害,昆虫体内物质也会相应调节进行解毒进而能够取食植物,二者相辅相成,共同进化[12]。已有研究表明香豆素和吲哚-3-甲醇可对昆虫P450基因产生诱导效应。吲哚-3-甲醇处理飞蝗L.migratoria 24 h,可诱导CYP6HL1和CYP6FE12基因的表达[14];斜纹夜蛾S.litura脂肪体中CYP4M14和CYP4S9基因表达均可被香豆素诱导[12],并且用高浓度香豆素处理48 h,CYP6AB14基因在中肠和脂肪体中的表达量均高于低浓度处理时的表达量,表现出剂量效应[13]。本试验将黏虫用0.1和0.5 mg/mL香豆素处理3 h,基因表达无诱导效应,可能是因为其对虫体产生较大刺激,未能及时做出反应。处理6、12、24和48 h,均有诱导效应,并且高浓度处理12、24和48 h基因相对表达量均显著高于低浓度处理,与之前研究结果相似,存在时间和剂量效应。0.1和0.5 mg/mL吲哚-3-甲醇处理每个时间点,基因表达均被诱导,说明可能在一定时间和剂量范围内,吲哚-3-甲醇对CYP9A113基因表达均产生诱导效应。
本试验克隆了黏虫的一条P450基因CYP9A113,并对外源物质对该基因表达的诱导效应进行了研究,初步探索了CYP9A113基因的功能及其与外源物质的相互作用,为今后该基因的系统研究及其应用于黏虫的防治提供基础依据。
参考文献
[1] NELSON D R, KAMATAKI T, WAXMAN D J, et al. The P450 superfamily: update on new sequences, gene mapping, accession numbers, early trivial names of enzymes and nomenclature[J]. DNA & Cell Biology, 1993, 12(1): 151.
[2] ASPERGER O, KLEBER H P. Distribution and diversity of bacterial cytochromes P450 [C]∥RUCKPAUL K.Microbial and Plant Cytochrome P-450. London: Taylor and Francis, 1991: 153.
[3] RAY J W. The epoxidation of aldrin by housefly microsomes and its inhibition by carbon monoxide [J]. Biochemical Pharmacology, 1967, 16: 99107.
[4] FEYEREISEN R. Arthropod CYPomes illustrate the tempo and mode in P450 evolution [J]. Biochimica et Biophysica Acta-Proteins and Proteomics, 2011, 1814(1): 1928.
[5] LI Xianchun, SCHULER M A, BERENBAUM M R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics [J]. Annual Review of Entomology,2007,52:231253.
[6] FEYEREISEN R. Insect P450 enzymes [J]. Annual Review of Entomology, 1999, 44: 507533.
[7] DING Tianbo, NIU Jinzhi, YANG Lihong, et al. Transcription profiling of two cytochrome P450 genes potentially involved in acaricide metabolism in citrus red mite Panonychus citri [J].Pesticide Biochemistry & Physiology,2013,106:2837.
[8] ANNE J, ANDERS M B G, LENE J R, et al. Biotransformation of polycyclic aromatic hydrocarbons in marine polychaetes [J]. Marine Environmental Research, 2008, 65(2): 171186.
[9] 趙华强,宋丽莉,李兵,等.野桑蚕细胞色素P450氧化酶活性测定及CYP9家族基因的诱导转录[J].浙江农业学报,2010,22(3):287291.
[10]刘月庆,樊星,周夏,等.甘蓝夜蛾CYP9A90基因的克隆及溴氰菊酯对其诱导表达[J].植物保护,2017,43(1):5460.
[11]王学贵,余慧灵,梁沛,等.氯虫苯甲酰胺诱导甜菜夜蛾细胞色素P450基因上调表达[J].昆虫学报,2012,58(3):281287.
[12]王瑞龙,孙玉林,梁笑婷,等.6种植物次生物质对斜纹夜蛾解毒酶活性的影响[J].生态学报,2012,32(16):51915198.
[13]WANG Ruilong, XIA Qingqing, BAERSON S R, et al. A novel cytochrome P450 CYP6AB14 gene in Spodoptera litura (Lepidoptera: Noctuidae) and its potential role in plant allelochemical detoxification[J]. Journal of Insect Physiology, 2015, 75: 5462.
[14]李亚红.杀虫剂及植物次生代谢物对飞蝗细胞色素P450基因的诱导表达[D].太原:山西大学,2015.
[15]江幸福,张蕾,程云霞,等.我国粘虫研究现状及发展趋势[J].应用昆虫学报,2014,51(4):881889.
[16]张智.北方地区重大迁飞性害虫的监测与种群动态分析[D].北京:中国农业科学院,2013.
[17]龚蕾,刘雁雨,焦必宁,等.植物中天然香豆素类化合物的提取纯化技术研究进展[J].食品工业科技,2015,36(20):377383.
[18]周颖,朱伟,杨杏芬.肿瘤化学预防剂吲哚-3-甲醇作用的靶点[J].热带医学杂志,2007(8):824827.
[19]赵华强,王东,李兵,等.家蚕细胞色素P450基因CYP9A22的克隆及在中肠和脂肪体的诱导转录[J].蚕业科学,2008,34(4):634641.
[20]GUO Yanqiong, ZHANG Xueyao, WU Haihua, et al. Identification and functional analysis of a cytochrome P450 gene CYP9AQ2 involved in deltamethrin detoxification from Locusta migratoria [J]. Pesticide Biochemistry and Physiology, 2015, 122: 17.
(责任编辑:田 喆)
猜你喜欢
小天使·二年级语数英综合(2020年5期)2020-12-23
电脑报(2020年28期)2020-07-31
学苑创造·A版(2016年10期)2016-11-19
计算机应用(2016年7期)2016-07-19
发明与创新·中学生(2016年3期)2016-03-29
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
时代英语·高二(2015年2期)2015-05-18
哈尔滨理工大学学报(2014年3期)2015-01-04
环球时报(2009-07-21)2009-07-21
