
河流鱼类分类群和功能群的纵向梯度格局
——以新安江流域为例
2019-09-04 09:30孟雪晨严云志
生态学报 2019年15期
陈 兵,孟雪晨,张 东,储 玲,严云志
安徽师范大学生命科学学院,安徽省高校生物环境与生态安全省级重点实验室, 芜湖 241000
河流生态系统具有极高的空间异质性,该异质性体现在栖息地斑块、河流纵向梯度、河流网络体等不同空间尺度[1- 2]。就河流纵向梯度尺度而言,从“上游-下游”,河流呈现为一个集连续性、等级性和异质性三重属性于一体的线性结构[3];沿着这一线性梯度,系列的非生物环境条件以及能量输入等都存在明显的空间异质性,进而影响了鱼类等水生生物群落的组成和数量分布[3]。已有不少研究表明,从河流的上游至下游,鱼类的物种数量逐渐增加,并往往在大型河流的中游河段(而非最下游河段)达到最大值[4- 6],该物种数的空间变化是由物种组成变化过程驱动的。从上游至下游,鱼类的物种组成变化可能表现为从上游至下游的物种递增(即物种获得)或从中游至下游的物种递减(即物种丢失),也可能同时表现为从一个群落到另一个群落的物种丢失和物种获得这2个过程[7- 8]。若仅含1个过程,则物种数量较少的群落是另一个群落的嵌套子集,其物种组成变化符合群落嵌套格局;若同时包含两个过程,则群落间的物种组成变化符合群落周转格局[9]。有研究表明,沿特定环境梯度下,若环境变化具有连续性,则群落的空间变化多呈嵌套格局[10- 13];相反,若环境条件呈跳跃式变化,则群落变化多为周转格局[14- 17]。
有关河流鱼类群落的纵向梯度变化的机制,目前已从栖息地多样性[18]、人类干扰强度[3]、中度干扰[19]、中域效应[20]等理论或假说中得到解释。其中,栖息地多样性假说强调的是局域栖息地多样性及其复杂性对鱼类群落的影响,其理论基础是基于生态位理论的环境过滤作用下的“确定模型”[18];中域效应假说强调的是鱼类自身的扩散过程及其潜在范围对鱼类群落的影响,其理论基础是基于中性理论的扩散过程作用下的“非确定模型”;此外,中度干扰假说强调的是环境的稳定性大小对鱼类群落的影响,其理论基础是中度干扰作用下的生物多样性最大化[19],而人类干扰强说强调的是人类活动的空间不均匀性及其对鱼类群落的负面影响[3]。有理由相信,河流鱼类群落沿“上游-下游”纵向梯度下的空间分布,可能是上述环境过滤、扩散限制、中度干扰、人为影响等多因素的联合作用结果[21]。但是,对于特定研究流域而言,其河流鱼类群落的纵向梯度变化,究竟是以哪类因素为主要驱动机制呢?遗憾的是,尽管目前有关河流鱼类群落的纵向梯度分布规律已得到很多报道,但据我们所知,目前尚未见有研究针对上述不同因素在驱动河流鱼类群落纵向梯度变化中的相对重要性进行报道。
此外,在已有的有关河流鱼类群落的纵向梯度格局研究中,绝大多数是针对基于物种组成的分类群(taxonomic organization)来开展研究的,而基于物种功能特征的功能群(functional organization)研究相对较少。需要指出的是,河流鱼类物种组成之所以随河流环境梯度而变化,主要是因为不同物种的生境选择、生理生态需求等存在差异,但是,环境压力的直接作用受体并不是物种本身,而是这些物种所呈现出来的食性、运动、繁殖等系列功能性状[22-24]。因此,在以往很多基于物种组成的鱼类分类群相关研究,生态学家们有时会观察到鱼类分类群组成及其多样性与环境条件之间的相关性不高这一现象,而开展鱼类的功能群及其多样性研究则能够很好的克服上述不足[25- 27]。已有研究表明,由于功能特征与物种组成的生态学属性不同,两者对环境变化的响应模式及其敏感程度也不尽相同[28-29],这意味着基于传统的分类群研究结果无法直接应用于功能群[5]。譬如,张东等研究发现,沿青弋江“上游-下游”纵向梯度,鱼类分类群空间变化主要以群落周转为主,而功能群变化则主要为群落嵌套[30]。然后,截止目前,有关河流鱼类功能特征的研究较为不足,河流鱼类功能群的纵向梯度格局及其形成机制是否、如何与分类群相区别尚不清楚。
基于上述认识,本研究以位于皖南山区的新安江流域为研究区域,沿“正源-河口”纵向梯度下连续设置调查样点,通过野外调查取样、鱼类功能性状测度等途径获得有关鱼类的物种组成、功能性状等因变量数据及局域栖息地、陆地景观和支流空间位置等自变量数据,在构建鱼类的分类群和功能群组成的基础上,比较研究了河流鱼类的分类群和功能群的纵向梯度分布特征及其群落变化规律,并解析了局域栖息地、景观条件和支流空间位置三类自变量对鱼类分类群和功能群的影响及其相对重要性。以期探明河流鱼类群落的纵向梯度格局及其形成机制。
1 材料与方法
1.1 研究区域
新安江流域位于安徽省和浙江省境内,发源于黄山市休宁县五股尖山,其海拔为1618.4 m,为怀玉山脉向西北延伸部分五龙山脉的主峰之一。新安江呈东北流向汇入千岛湖水库,属于钱塘江水系。流域面积约为11000 km2,流域干流全长约370 km,其中安徽段长240 km。新安江地处我国亚热带湿润季风气候区,全年四季分明;年平均气温18.0℃(1—29℃),年降水量约1800 mm,主要集中在4—9月,占全年降水量的80%[31]。
1.2 样点设置与标本采集
沿五股尖河源至河口的纵向梯度,每间隔3—8 km设置采样点,共27个样点。野外作业过程中,根据样点的可抵达性和可操作性来选择具体调查样点,且各调查样点远离诸如水坝、农田和城镇等明显人为干扰(至少间距1 km)。根据1∶300000比例的水系图对各个调查样点所处河段的河流级别(stream order)大小进行量化[32],27个调查样点位于1级至5级河流之中,其中1—5级河流的调查样点数分别为4个、6个、6个、5个和6个(图1)。分别于2017年5月(丰水期)和11月(枯水期)共两次对上述调查样点进行鱼类取样。根据调查样点的水深情况选择具体渔具类型:水深不足1 m的1—3级河段样点,采用背式电鱼器直接涉水取样;水深超过1 m的4—5级河段样点,采用船运电鱼器沿河流岸边进行取样。各样点采样河长为100 m,采样时间40 min。依据Nelson和陈小勇的目科级分类系统,在鱼类新鲜状态下进行鉴定(吻虾虎鱼仅鉴定至属的水平),记录各样点的鱼类物种组成及其数量。疑难种以甲醛固定后进一步鉴定[33- 34]。
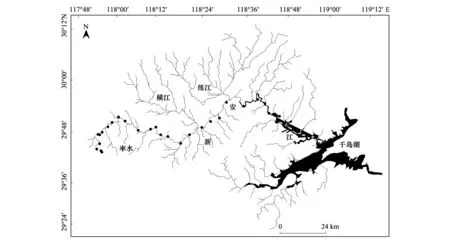
图1 新安江研究样点示意图Fig.1 Sampling sites in the Xin′an River, showed by black circles黑色圆圈代表采样点
1.3 环境因子测定
每次采样时,使用GPS现场标记经纬度并测量局域环境栖息地变量,包括:水宽(m)、水深(m)、流速(m/s)、水温(℃)、溶氧(mg/L)、pH、电导率(mS/cm)。将每个调查河段分为3个等距的断面,在每个断面上测量水宽;其次,在每个断面上选取3个等距的截点,在每个截点处测量水深,并在每个截点的60%水深处使用便携式流速仪(FP111)测量水流流速;水深超过1.5 m深水区使用手持式测深仪(Speedtech SM- 5A)测量水深。水宽、水深和流速均取各断面均值。使用多参数水质监测仪(YSI Professional Plus)测量每个样点的水温、溶氧、pH和电导率。
利用ArcGIS 10.0和精度为30 m的DEM卫星图层,提取各取样河段所在集水区(catchment)的土地面积,在将土地利用/覆盖划分为耕地、林地、城镇、滩地、草地共五种类型的基础上,提取出上述各类型土地的占地面积,进而结合集水区的土地面积计算出上述耕地、林地、城镇、滩地和草地的相对面积(%)。进一步,利用ArcGIS软件计算各样点至河源的河道距离,将其视为各样点的距河源距离。
参照Yan 等的支流空间位置划分方法[35],根据 1∶300000比例的水系图,对各调查样点在新安江河流网络中的空间位置进行量化,包括:河流级别[32](stream order)、流量量级[36](link magnitude)、汇合量级[37](confluence link,C-link)、下游量级[38](downstream link,D-link)。
1.4 功能性状测度及功能群构建
考虑到鱼类物种(尤其是鲤科鱼类)在不同生命阶段(如幼鱼期和成鱼期)具有潜在的食性及其他功能性状的转化,本研究仅针对每物种的成鱼进行性状测度;由于本文着重研究鱼类的功能群(而非功能多样性)的空间变化,因而功能性状测度针对物种进行(忽略不同样点间的潜在种内功能性状变异)。当某物种的成鱼标本不足30尾时,全部标本用于性状测度;当某物种的成鱼标本超过30尾,随机抽取30尾用于性状测度。参照Villéger等的方法,共测度11组绝对性状,并将其换算成7组比例性状(表1),这7组性状共反映了鱼类的营养特征、运动能力、栖息地利用3个不同功能特征[39- 42]。

表1 基于形态学所测功能特征
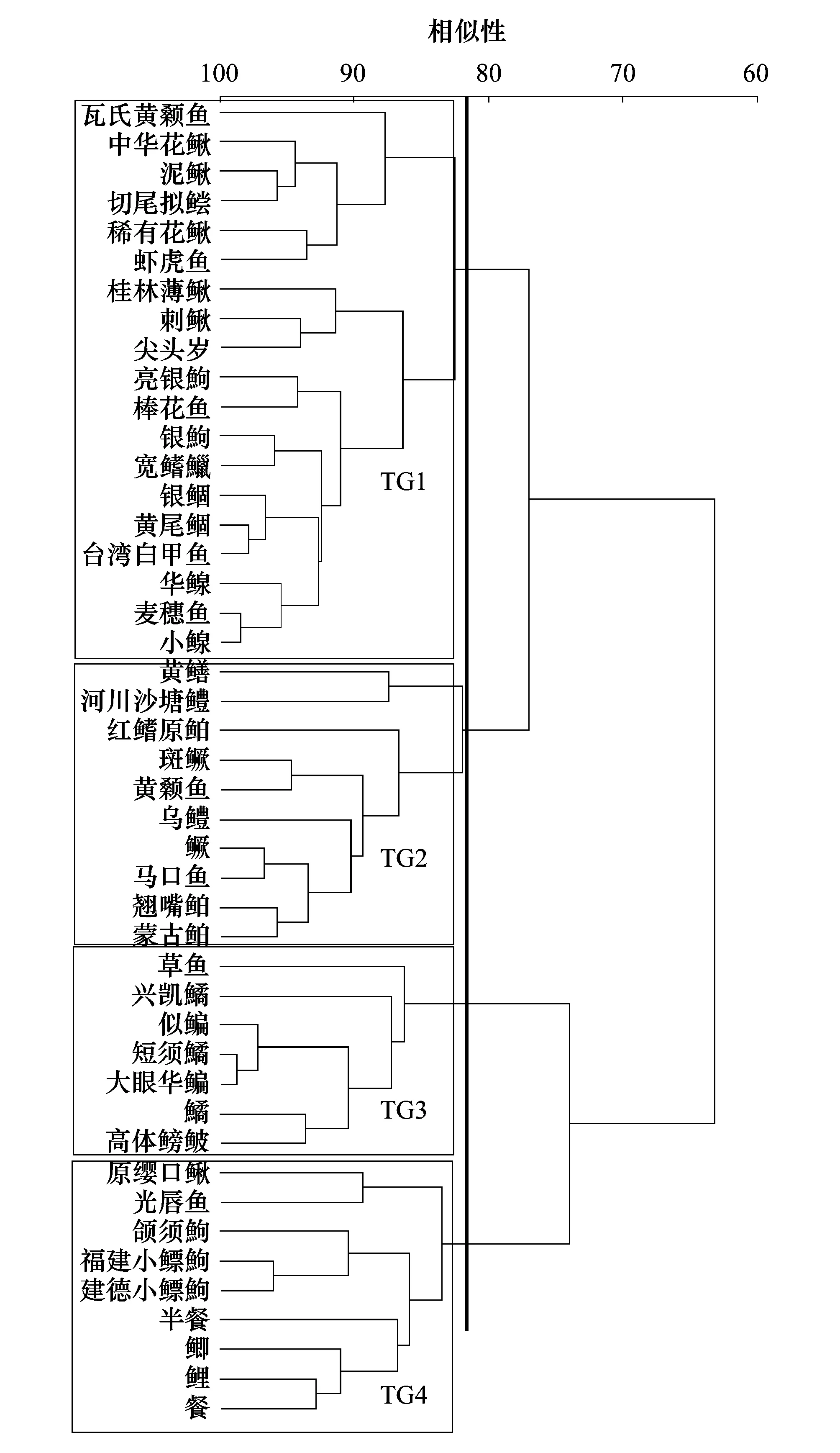
基于上述测度的功能性状,在Primer 5.0软件下构建物种间Bray-Curtis 相似性矩阵,运用聚类分析确定全部采集的44种鱼类在营养、运动及栖息地利用上的聚类分组情况,分别确定鱼类的营养功能群(trophic group,TG)、运动功能群(locomotion group,LG)及栖息地利用功能群(habitat-preference group;HG);进一步,根据每种鱼类在上述3个功能群上的隶属情况,确定研究区域内鱼类的“营养-运动-栖息地利用”复合功能群(combined group,CG)组成。
1.5 数据分析
计算每物种的出现频率(Frequency of the occurrence,Fi)和相对多度(Relative abundance,Pi),然后依照Fi≥40%、10%≤Fi<40%以及Fi<10%来确定每个物种为常见种、偶见种或稀有种,另外按照Pi<10%和Pi≥10%将物种定义为非优势种和优势种。出现频率与相对多度的计算公式分别为Fi=(Si/S)×100% 与Pi=(Ni/N)×100%,式中Si代表物种i出现样点数,S代表总样点数;Ni代表物种i的个体数,N代表全部物种的个体数。
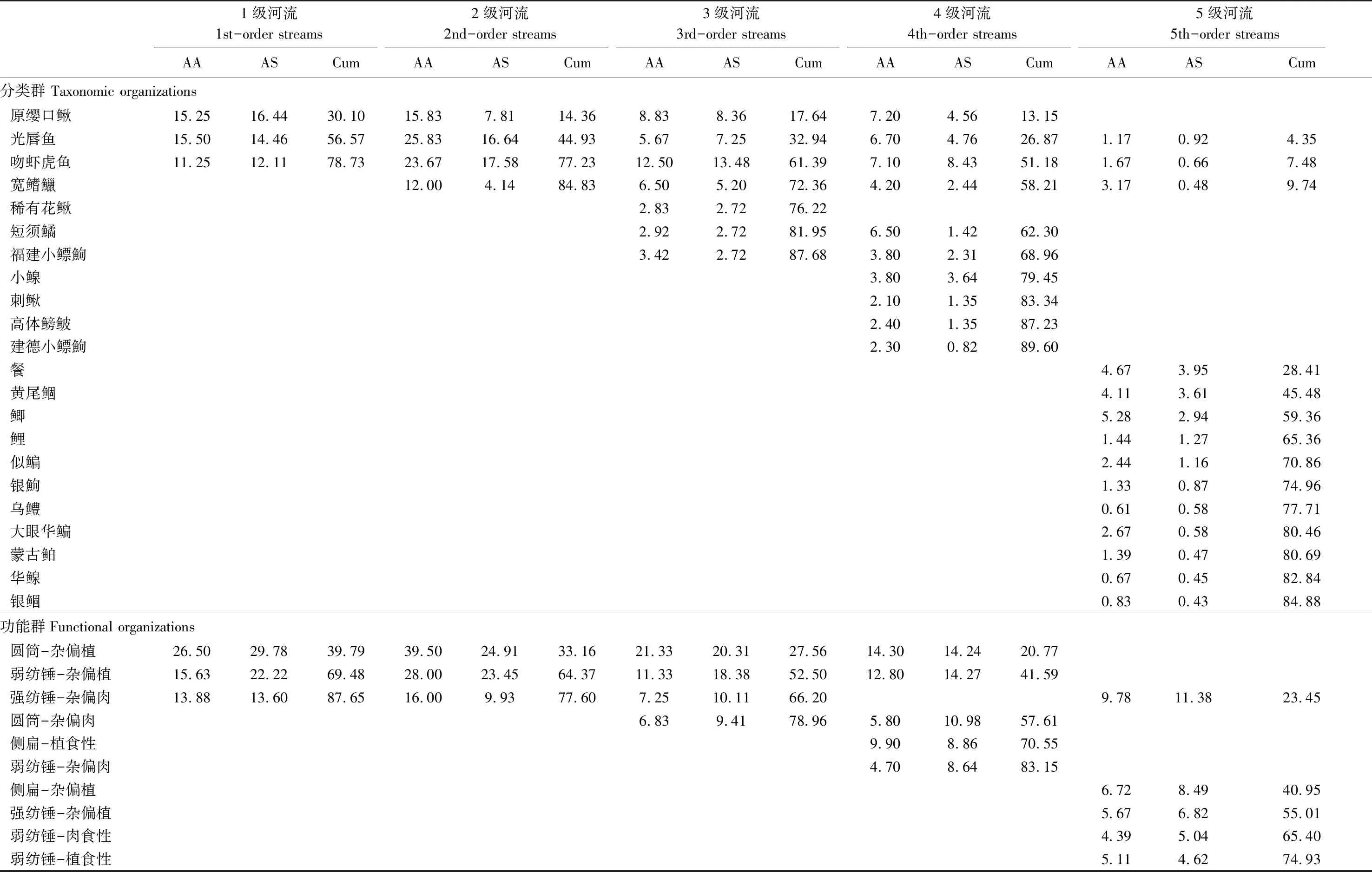
根据SPSS 20.0软件,运用Pearson′s相关性检验每个自变量与距河源距离之间的相关性,进而确定自变量的纵向梯度分布规律;其中,因局域栖息地变量存在季节变化(陆地景观变量和支流空间位置变量均不存在季节变化),根据其5月和10月测度数据的均值来进行上述分析。在Primer 5.0软件下,对鱼类分类群和功能群组成数据进行log10(X+1)转换,再运用双因素交互相似性分析(Two-way crossed ANOSIM)检验河流级别和季节对结构的影响,视GlobalR值确定鱼类群落的分离程度:R>0.75,群落完全分离;0.5 根据CANOCO 4.5软件,采用冗余分析(Redundancy analysis, RDA)分别检验鱼类分类群和功能群结构与距河源距离之间的关系。全部结构数据都进行了log10(x+1)转换, 以降低极端数值的负面影响。 Pearson′s相关分析结果显示,除了溶氧和草地面积不随距河源距离显著变化以外(P>0.05),其他12组环境变量都随距河源距离显著变化(P<0.05),其中,流量量级、下游量级、水温、电导率、水宽、水深、pH、耕地面积、滩地面积和城镇面积与距河源距离呈正相关,而汇合量级、流速、林地面积和草地面积与距河源距离呈负相关(表2)。 2.2.1分类群 共采集鱼类3004尾,隶属4目10科44种。鲤形目物种数最多,达34种,其中鲤科鱼类计35种,占总物种数的77.78%。1—5级河流各样点采集的平均物种数分别为6种、7种、10种、14种和12种,由上游至下游呈上升趋势。翘嘴鲌(Culteralburnus)、黄尾鲴(Xenocyprisdavidi)、红鳍原鲌(Cultrichthyserythropterus)、华鳈(Sarcocheilichthyssinensis)、似鳊(Pseudobramasimoni)以及银鲴(Xenocyprisargentea)等仅出现在5级河流中;光唇鱼(Acrossocheilusfasciatus)、宽鳍鱲(Zaccoplatypus)、吻虾虎鱼(Rhinogobiussp.)、原缨口鳅(Vanmaneniastenosoma)以及稀有花鳅(Cobitisrarus)在1—4级河流均有出现;而台湾白甲鱼(Onychostomabarbatulum)和尖头鱥(Phoxinusoxycephalus)仅见于1级河流。光唇鱼、宽鳍鱲、吻虾虎鱼、原缨口鳅、稀有花鳅、建德小鳔鮈(Microphysogobiotafangensis)以及福建小鳔鮈(Microphysogobiofukiensis)出现频率均大于40%,属于常见种;而台湾白甲鱼、尖头鱥(Phoxinusoxycephalus)以及斑鳜(Sinipercascherzeri)的出现频率均小于10%,属于稀有种。光唇鱼、宽鳍鱲以及吻虾虎鱼为新安江流域的优势种,其相对多度大于10%(表3)。 表2 环境因子概况及其与距河源距离的关系 Table 2 Summary statistic for environmental variables including tributary spatial position, local habitat and land use/cover, and their relationships with the distance from headwater(DFH) 环境变量Environmental variables缩写Abbrev.最小值-最大值Min-Max平均值±标准差Mean±SD与距河源距离相关性Relationship with Distance from headwater(DFH)RP支流空间位置Tributary spatial position流量量级 Link magnitudeLink1—14452.74±51.010.668∗∗汇合量级 Confluence linkC-link4—5231.63±16.66-0.782∗∗下游量级 Downstream linkD-link4—14453.30±50.430.943∗∗局域栖息地Local habitat溶氧Dissolved oxygen/(mg/L)DO0.84—2.951.46±0.58-0.351ns水温 Water temperature/℃WT17.30—31.8026.35±4.480.947∗∗电导 Conductivity/(mS/cm)CO80.0—219.5129.36±46.460.519∗∗水宽Wetted width/mWW 2.00—312.80101.62±98.000.968∗∗水深Water depth/mWD0.17—12.002.39±3.410.690∗∗流速Current velocity/(m/s)CV0.10—0.800.4±0.24-0.671∗∗pH值PH5.85—9.227.72±0.830.762∗陆地景观变量Landscape condition林地面积Forest area/%FA72.23—94.7183.02±6.53-0.839∗∗草地面积Meadow area/%MA2.36—8.335.17±1.73-0.224ns耕地面积Agriculture area/%AA1.86—21.9911.04±6.450.786∗∗滩地面积Wetland area/%WA0—0.870.22±0.320.653∗∗城乡面积Urban area/%UA0—1.790.56±0.600.640∗∗ **,P< 0.01;*,P< 0.05;ns,P> 0.05 2.2.2功能群 聚类结果显示,在相似性系数为91.5%时,采集的44种鱼类被划分为5个运动功能群(分别为LG1—LG5),其中LG3、LG4和LG5的组内相似性主要由“体高/体长”比决定。 LG3的体高/体长比最大比值范围为0.32—0.47,所含物种包括短须鱊、兴凯鱊、高体鳑鲏、鳜和斑鳜等,对应的体型特征为侧扁型;LG4的体高/体长比最小,仅黄鳝一种,其比值为0.05,对应的体型特征为鳗型;而LG5的体高/体长比居中,比值范围为0.09—0.19,所含物种包括刺鳅、稀有花鳅、泥鳅、中华花鳅以及河川沙塘鳢等,对应的体型特征为圆筒形。而LG1和LG2的体高/体长比范围为0.17—0.39,对应的体型特征为纺锤形,且两者的组间相似性差异主要由“最大尾鳍高/最大尾柄高”比决定;其中,LG1的最大尾鳍高/最大尾柄高比高达2.45—3.12,所含物种包括黄尾鲴、银鲴、草鱼以及银鮈等,其游泳能力相对较强,被称为“强纺锤形”;LG2的最大尾鳍高/最大尾柄高比为1.82—2.38,所含物种包括麦穗鱼、马口鱼、黄颡鱼等,其游泳能力相对较弱,被称为“弱纺锤形”(图2)。在相似性系数为82.5%时,44种鱼类被划分为4个营养功能群(分别为TG1—TG4),其组间相似性差异主要由“肠长/体长”比决定。TG2的肠长/体长比最小,起比值为0.13—0.33,所含物种包括黄鳝、河川沙塘鳢、鳜、乌鳢、马口鱼等,对应的食性特征为肉食性;TG3的肠长/体长比最大,比值范围为3.05—6.86,包括草鱼、兴凯鱊、高体鳑鲏等,对应的食性特征为植食性。TG1和TG4的肠长/体长比居中,其中TG1的肠长/体长比略低,(0.41—0.78),包括切尾拟鲿、刺鳅、麦穗鱼、小鳈以及棒花鱼等,对应的食性特征为杂食偏肉食性;TG4的肠长/体长比略高(0.93—2.35),包括鲫、鲤、福建小鳔鮈、餐等,对应的食性特征为杂食偏植食性(图3)。 图2 基于聚类分析划分5类运动功能群Fig.2 Classification in locomotion groups for fish assemblages based on Cluster AnalysisLG1:运动功能群1(强纺锤形) Locomotion group 1(spindle shape with strong athletic ability); LG2:运动功能群2(弱纺锤形) Locomotion group 2(spindle shape with week athletic ability);LG3:运动功能群3(侧扁型) Locomotion group 3(compressed shape);LG4:运动功能群4(鳗型) Locomotion group 4(eel shape);LG5:运动功能群5(圆筒形),Locomotion group 5(cylindrical shape) 图3 基于聚类分析划分4类营养功能群 Fig.3 Classification in trophic groups for fish assemblages based on Cluster AnalysisTG1:营养功能群1(杂食偏肉食性) Trophic group 1(Omnivores slant carnivorous);TG2:营养功能群2(肉食性) Trophic group 2(Carnivorous);TG3:营养功能群3(植食性) Trophic group 3(Herbivores);TG4:营养功能群4(杂食偏植食性) Trophic group 4(Omnivores slant herbivores) 根据反映鱼类栖息地利用的2组功能性状(“眼位/头高”比、“体高/体宽”比)的聚类分析结果,无法将44种鱼类进行有效划分。因此,本文仅根据鱼类的运动特征和营养特征来构建44种鱼类的“运动—营养”复合功能群,共含14个复合功能群(表3)。 表3 新安江流域鱼类的物种组成、营养、运动和复合功能群类型及其出现频率和相对多度 双因素交互相似性分析显示,5月和11月间的鱼类分类群和功能群组成均无显著的季节变化(GlobalR=0.024,P> 0.05,分类群;GlobalR=0.081,P> 0.05,功能群),但1级—5级河流间的分类群和功能群都具有显著的空间变化(GlobalR=0.402,P< 0.01,分类群; GlobalR=0.358,P<0.01,功能群),且分类群(Global R值较高)的空间变异稍高于功能群。由NMS图可知,无论是分类群还是功能群,1级—3级河流间的群落结构重叠度较高,而4级—5级河流间的重叠度较低(图4)。 图4 1—5级河流间鱼类的分类群和功能群结构NMS双标图Fig.4 Non-metric multidimensional scaling (NMS) analysis plots of taxonomic and functional organizations of stream fishes in the Xin′an River 运用相似性百分比分析确定出维持各级别河流鱼类群落组内相似性的关键物种,结果显示,由1级至3级河流,关键种的变化仅表现为“物种获得”1个过程,即:在光唇鱼、吻虾虎鱼和原缨口鳅的基础上(1级河流),逐渐获得宽鳍鱲(2级河流)和稀有花鳅、福建小鳔鮈、短须鱊以及切尾拟鲿(3级河流);由3级至5级河流,关键种的变化表现为“物种丢失”和“物种获得”2个过程,即:由3级至4级,稀有花鳅丢失,而小鳈、刺鳅和高体鳑鲏等获得;由4级至5级河流,原缨口鳅、短须鱊、小鳈等丢失,而鲫、黄尾鲴和似鳊等获得(表3)。功能群也经历了类似的空间变化过程,由1级至3级河流,关键功能团的变化仅包含“功能获得”过程,即:在具“圆筒-杂偏植”、“弱纺锤-杂偏植”和“强纺锤-杂偏肉”的基础上(1—2级河流),获得了“圆筒-杂偏肉”(3级河流)。由3级至5级河流,关键功能团的变化则包括“功能丢失”和“功能获得”2个过程,即:由3级至4级河流,“强纺锤-杂偏肉”丢失而“侧扁-植食性”和“弱纺锤-杂偏肉”获得;由4级至5级河流,“圆筒-杂偏植”、“弱纺锤-杂偏植”、“强纺锤-杂偏肉”、“侧扁-植食性”等丢失,而“侧扁-杂偏植”、“强纺锤-杂偏植”、“弱纺锤-肉食性”、“弱纺锤-植食性”等获得(表4)。 进一步,运用冗余分析检验了鱼类分类群和功能群与距河源距离之间的关系,结果显示,分类群和功能群都随距河源距离显著变化,且分类群与距河源距离的相关性略高于功能群(r=0.784,P<0.01,分类群;r=0.745,P<0.01,功能群)。根据RDA双标图,尖头鱥、台湾白甲鱼、稀有花鳅、原缨口鳅、切尾拟鲿等物种与距河源距离呈负相关,而黄颡鱼、兴凯鱊、似鳊以及乌鳢等与距河源距离正相关。就功能群而言,圆筒-杂偏植、弱纺锤-杂偏植、圆筒-杂偏肉等功能团与距河源距离呈负相关,而侧扁-肉食性、侧扁-植食性、圆筒-肉食性等功能团则与河源距离正相关(图5)。 图5 距河源距离与鱼类分类群和功能群结构关系的冗余分析图Fig.5 Redundancy analysis diagrams for the relationship between distance from headwater and taxonomic and functional organizations 向前选择结果显示,显著影响鱼类分类群和功能群的局域栖息地变量一致,均为水宽和流速,陆地景观变量也一致,均为耕地面积和滩地面积,但支流空间位置变量不一致,其中影响分类群的是汇合量级和下游量级,而影响功能群的是流量量级。根据方差分解结果,三类自变量对分类群和功能群的总解释率分别为33.6%和38.5%;局域栖息地、陆地景观和支流空间位置对分类群的独立解释率分别为4.8%、1.8%和2.3%,对功能群的独立解释率分别为6.3%、3.7%以及1.0%,三类环境变量的共同解释率分别为16.9%和27.4%。此外,根据“rda”函数解释环境变量与群落结构相关性分析结果,分类群受局域栖息地和支流空间位置两类变量的显著影响(P<0.05),而功能群则受局域栖息地和陆地景观变量的显著影响(P<0.05)(图6)。 河流生态系统具有极高的空间异质性,从上游至下游,河流可被视为一个集连续性、等级性和异质性于一体的线性结构[1- 2];沿着这一线性结构梯度下,系列理化因子都呈现出明显的纵向梯度变化规律[3]。本研究发现,就局域栖息地因子而言,除了溶氧以外,其他7组栖息地变量均随距河源距离显著变化:从上游至下游,水温、水宽、水深、电导率和pH值都显著上升,而流速显著下降。此外,本研究还发现,集水区的土地利用面积也随河源距离显著变化(除草地面积以外),其中林地面积显著下降,但耕地、滩地和城乡面积显著上升。沿河流纵向梯度下的栖息地条件变化,也带来了鱼类群落组成和数量特征的变化[18]。就物种组成而言,Huet(1957)针对欧洲地区的河流鱼类纵向梯度变化提出“生物带”现象,即从上游至下游,其鱼类区系组成变化经历了“鲑亚科-茴鱼科-鲤科”等变化;类似地,在北美温带地区,从上游至下游,其鱼类组成也经历了由“鲑亚科-鲈形目-鲇形目”变化[3,15]。本研究发现,新安江流域1—5级河流间的鱼类群落组成存在显著差异,且鱼类群落随距河源距离显著变化。尖头鱥、台湾白甲鱼喜低温,局限分布于河流上游的高海拔山地溪流;光唇鱼、吻虾虎鱼、原缨口鳅、宽鳍鱲等物种具有亲流性特点[44],因而主要分布于河流的中上游河段,尽管光唇鱼和宽鳍鱲在中下游河段也有少量分布,但数量明显下降;稀有花鳅和中华花鳅喜底质为砂砾、水流速度较缓的水体,主要分布于中游河段;小鳈、短须鱊等物种喜缓流,主要分布在中下游河段,而鲫、黄尾鲴、似鳊等静水性物种主要分布于下游河段。与物种组成类似,河流鱼类的营养、运动等功能特征也具有一定的纵向梯度变化规律[45- 47];以营养特征为例,河流上游水体的初级生产力极其低下,鱼类的饵料主要来源于河岸两侧的陆地生态系统;相反,河流下游具有较高的初级生产力。因此,沿河流上游至下游,鱼类群落营养往往呈现出“无脊椎动物食性-杂食性-草食性-肉食性”的梯度变化规律[3]。本研究发现,尽管中上游河段的鱼类体型较为多样(包括圆筒形、强纺锤形、弱纺锤形),但其食性较为单一,主要为杂食性;在中下游河段,则出现了侧扁体型物种(如高体鳑鲏、兴凯鱊、鲫、鳜等),也出现了草鱼、大眼华鳊、高体鳑鲏、短须鱊等植食性物种以及乌鳢、翘嘴鲌、鳜、黄颡鱼等肉食性物种。就体型而言,尽管河流上游河段的落差较大、水流较急,但因山地溪流系统中普遍存在的 “急滩-深潭”序列(riffle-pool sequence;两类生境彼此交替重复出现),其中的深潭生境的水流相对较缓,可为圆筒形体型、游泳能力较弱的物种提供栖息场所;同时,诸如吻虾虎鱼、原缨口鳅等圆筒形体型物种在河源溪流中的急滩生境也常有分布,这些溪流底栖型鱼类可通过一些特殊的构造来适应急流条件,包括高度特化的腹鳍(吻虾虎鱼,腹鳍愈合成吸盘状构造)以及胸鳍(原缨口鳅,胸鳍和腹鳍平展并向外侧延伸)[44]。就营养特征而言,因河流上游河段具有营养外源性特点(因水流清急、底质粗糙、岸边植被覆盖等的影响,除了固着藻类以外无其他水生植物生长),营养来源主要为水生昆虫、陆生植物的枯枝烂叶等,因而其中的鱼类为杂食性[3];受昆虫羽化、植物生长等物候节律的影响,这些杂食性鱼类的食物组成具有明显的季节变化特点[48];相反,在中下游河段,因水生维管束植物可以大量生长,其营养特点也转变为以内源性为主(至少内源性营养所占比例大幅度增加),因而其中出现植食性物种;相应的,因河流中下游河段的栖息地稳定性升高、鱼类种类数增多且食性等生态位分化加强,肉食性物种才逐渐增多[3]。 表4 基于百分比分析的1级—5级河流中的关键物种和功能群 AA:平均多度 Average abundance;AS:平均相似性 Average similarity;Cum:累计贡献率 Contribution 图6 三类环境变量影响分类群和功能群的方差分解图Fig.6 The variation partitioning of effects of environmental variables on taxonomic and functional organizations of fish assemblagesns:P > 0.05;*:P< 0.05;**:P< 0.01 从一个群落到另一个群落,其物种组成变现为物种丢失和/或物种获得[8];若仅含其中1个过程(物种丢失或物种获得),则物种数较少的小群落是另一个群落的嵌套子集,其群落变化呈现为群落嵌套(nestedness)格局;若物种丢失和物种获得兼之,则群落间存在物种周转,其群落变化呈现为群落周转(turnover)格局[9]。在河流生态系统中,沿着“上游-下游”纵向梯度,因栖息地复杂性和多样性的变化,河流鱼类物种数往往逐渐增多并可能在中游河段达到最大值[4- 6];其物种组成常通过递增(addition)或替换(replacement)模式变化[7],分别符合群落嵌套和群落周转格局。有研究发现,沿着“上游-下游”纵向梯度,若环境条件变化具有渐变性特点,则鱼类物种组成常以嵌套格局变化[10- 13];但若环境条件变化具有间断性特点(如陆地景观由山地变为丘陵或平原),则其鱼类物种组成变化符合周转格局[14- 17]。本研究发现,从1级至3级河流,鱼类群落中的关键种变化仅含物种获得1个过程,呈群落嵌套格局;从3级至5级河流,其关键种变化则包含物种获得和物种丢失2个过程,呈群落周转格局。首先,从水系组成来看,新安江干流上游有3条主要支流,分别是率水、横江和练江,本研究中27个调查样点中,1级—3级河段样点位于率水子流域,4级河段样点位于率水-横江汇合点至新安江-练江汇合点之间,而5级河段调查样点位于新安江-练江汇合点下游(图1)。其次,从地形地貌来看,尽管新安江地处皖南山区,但其中的部分河段位于徽州盆地;本研究中的1级—3级调查河段的坡度较大,位于徽州盆地外缘;4级调查河段的坡度较小,位于盆地之中;而大部分5级调查河段的坡度也较大,位于盆地外缘(表5)。因此,由1级至3级河流,可能因环境条件的渐变特点,其鱼类群落中的关键物种/功能团的变化呈嵌套格局;相反,由3级至5级河流,无论是主要支流的汇入还是地形地貌的变化,都可能导致其环境条件的骤变,因而其鱼类群落中的关键物种/功能团的变化呈周转格局。 表5 新安江样点信息表 河流鱼类群落组成和数量的分布,是历史因素、非生物因素和生物因素的联合作用产物[49-51]。就现时的非生物因素而言,首先,由于鱼类的栖息地选择、生理需求等存在种间差异,因而沿河流纵向梯度下的水温、水深、流速、底质大小等栖息地变量的变化对河流鱼类群落的分布有着重要影响[52-55]。其次,由于河流系统中的水源补给、营养输入等受集水区的地形地貌、植被盖度、土地利用等的影响,因而陆地景观条件也直接或间接影响河流鱼类物种的分布[56-58]。本研究发现,在新安江流域,水宽和流速等栖息地变量及农田和滩地面积等景观变量对鱼类的分类群和功能群都有着显著影响。此外,局域鱼类群落的组成与数量还受到鱼类的迁入/迁出等扩散过程的影响[13,35,59]。河流系统是一个由干流与各级支流共同组成的树状生态系统,简称为河网结构[60];干流与支流及各级支流之间的连通性,决定了河网结构下的生态因子及其过程的空间的相关性。不同的支流在河网结构中的空间位置不同,其鱼类的迁入、迁出速率也有潜在差异,因而对局域鱼类群落的组成也构成重要影响[61- 63]。本研究发现,流量量级及汇合量级、下游量级等支流空间位置变量对新安江鱼类分类群和功能群有着显著影响。尽管河流鱼类群落受局域栖息地、景观特征和支流空间位置等因素的联合影响,但是,在特定的空间尺度下,各类因素在影响鱼类群落中的相对重要性可能不同[2]。本研究发现,就局域栖息地与景观特征而言,局域栖息地对鱼类群落空间变化的解释率高于景观变量,该现象也见于Zhu等的研究结果[31]。究其原因,可能有二,其一,农业、城镇化等土地利用是通过改变水源供给、营养输入等途径来间接影响河流系统及其鱼类群落,而水温、流速等局域栖息地特征是直接影响河流鱼类群落[61- 64];其二,以城镇化为例,有研究表明,河流鱼类群落对这一人类干扰的响应,并不取决于城镇占地总面积,而是依赖于其中的不渗水土地面积[65],但在本研究中,受技术手段的限制,我们仅能提取出较为粗放的土地利用类型及其占地面积。此外,本研究还发现,支流空间位置对新安江鱼类分类群的影响也低于局域栖息地,而功能群则不受支流空间位置的显著影响。局域栖息地特征是河流系统中的非生物环境条件,其对鱼类群落的影响反映了“环境过滤”机制在群落构建中的作用;相反,支流空间位置主要是通过影响局域鱼类群落的迁入/迁出速率而对鱼类群落时间影响,其对鱼类群落的影响反应的是“扩散限制”机制在群落构建中的重要性。在本研究中,局域栖息地和支流空间位置对鱼类分类群有着显著的联合影响,该结果说明研究区域内的鱼类群落的构建既符合以生态位理论为基础的“确定”模型,又在一定程度上符合基于中性理论的“随机”模型,但前者相对于后者而言更为重要。Yan等在河流网络尺度下对皖南山区的青弋江流域鱼类群落的空间格局进行了研究,着重比较了局域栖息地和支流空间位置影响局域鱼类群落的相对重要性,经研究发现,流域中大量存在的水坝可能通过阻断鱼类洄游通道、限制鱼类扩散过程来削弱支流空间位置在影响鱼类群落中的相对重要性[35]。在本研究的野外调查过程中,尽管我们在选点时尽量避开明显人为干扰,但对于新安江流域也普遍存在的中小型水电站、低头坝等水坝却无法完全回避。因此,与Yan等的研究结果类似,可能由于水坝等人类干扰在一定程度上限制了鱼类的迁入/迁出过程,故而造成支流空间位置变量对鱼类群落的响应重要性较低[35]。 传统的群落生态学研究主要是对基于物种组成的分类群进行研究,以探明分类群的时空变化及其对环境变化/人类干扰的响应[21]。沿着某一环境梯度下,河流鱼类分类群之所以呈现出一定的空间异质性,主要是因为不同的鱼类物种的生境选择、生态需求等存在种间差异。然而,环境变化的直接作用受体并不是物种本身,而是物种所具有的运动、营养、繁殖等功能性状[22- 24]。有研究表明,同分类群相比,鱼类功能群组成和数量变化能更为直接的反映环境变化对鱼类群落的影响,能更有效的揭示生态系统的过程、抗性和价值等。本研究发现,就局域栖息地和景观条件而言,这两类环境因素与鱼类功能群的相关性都高于分类群,这进一步说明河流鱼类功能群与环境条件之间的关系更为密切。此外,本研究还发现,尽管分类群变化受局域栖息地和支流空间位置的联合影响,但支流空间位置对功能群变化无显著影响。既然鱼类的扩散过程是下游量级和汇合量级等支流空间位置变量影响鱼类群落分布的重要机制,那么,本研究结果说明,至少在新安江流域,鱼类的扩散过程尽管在一定程度上影响鱼类的分类群结构,但对鱼类的功能群结构无显著影响。当环境过滤起作用时,相似的环境条件下有相似的鱼类群落(包括分类群和功能群);当扩散过程起作用时,相似的环境条件下也可能有不相似的鱼类群落(仅为分类群),但这些鱼类群落在功能特征上却可能比较类似,这体现了鱼类的物种组成与功能特征的不对等性,即“功能冗余”现象。换句话说,功能冗余这一现象可能削弱了扩散过程对鱼类功能群的影响。此外,也正是由于功能冗余的作用,造成了本研究中的另一个研究结果,即1—5级河流间鱼类功能群的空间变异稍低于分类群。以体型为例,新安江上游优势物种的体型较为多样,圆筒形、强、弱纺锤形等体型鱼类都具有;至河流中下游,除了新增了侧扁形体型鱼类以外,以上的圆筒形和纺锤形体型鱼类仍然保留——尽管物种组成可能不同。类似地,以食性为例,新安江上游优势物种的食性主要为杂食性,至河流中下游,尽管逐渐获得了植食性和肉食性物种,但也仍保留了很多杂食性鱼类(物种组成虽有很大不同)。 综上,本研究表明,沿着河流“上游-下游”纵向梯度下,局域栖息地条件、集水区景观特征等环境条件显著变化,鱼类的物种组成(分类群)和功能特征(功能群)也相应变化,但功能群的空间变异略低于分类群。在中上游河段,鱼类分类群的纵向梯度变化呈嵌套格局,主要表现为亲流性物种的获得过程;在中下游河段,分类群呈嵌套格局,主要表现为亲流性物种的丢失及缓流/静水性物种的获得过程;从上游至下游,鱼类功能群的变化主要表现为侧扁形体型及肉食性和植食性物种的获得过程。鱼类分类群受局域栖息地和支流空间位置变量的联合影响,是环境过滤和扩散过程的共同作用产物,而功能群的构建仅受环境过滤影响。因此,为合理保护和管理新安江流域的鱼类多样性,有必要兼顾局域栖息地、土地利用/覆盖及河道连通性的保护和恢复。
2 结果
2.1 环境变量的纵向梯度格局
2.2 分类群和功能群的组成概况



2.3 分类群和功能群的时空变化


2.4 三类自变量对分类群和功能群的影响
3 讨论


猜你喜欢
舰船科学技术(2022年21期)2022-12-12
数学物理学报(2022年1期)2022-03-16
山西大学学报(自然科学版)(2021年5期)2021-12-25
数学物理学报(2021年6期)2021-12-21
数学物理学报(2021年5期)2021-11-19
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
汉语世界(The World of Chinese)(2018年3期)2018-10-22
厦门航空(2018年4期)2018-04-25
中国医学装备(2015年10期)2015-12-29
太阳能(2015年12期)2015-04-12
