
一株H5N3亚型禽流感病毒全基因组序列分析及其对小鼠的感染性研究
2019-09-04 03:03韩舒雨崔鹏飞崔嘉琦谷文丽邓国华陈化兰
中国预防兽医学报 2019年7期
韩舒雨,崔鹏飞,崔嘉琦,谷文丽,邢 鑫,邓国华,陈化兰
(中国农业科学院哈尔滨兽医研究所兽医生物技术国家重点实验室/农业部动物流感重点开放实验室,黑龙江哈尔滨150069)
禽流感病毒(Avian influenza virus,AIV)属于正粘病毒科流感病毒属,其基因组由8个分节段的单股负链RNA组成,编码11种病毒蛋白。依据AIV核蛋白(NP)和基质蛋白(M)抗原性及基因特性的不同可将流感病毒划分为A、B、C型。根据其表面糖蛋白(Hemagglutinin,HA)及神经氨酸酶(Neuraminidase,NA)进行血清学分型,已鉴定出18种HA和11种NA亚型[1],除了H17N10亚型和H18N11亚型分离自蝙蝠外,其余所有亚型均分离自禽类。根据对鸡的致病力强弱可将其分为高致病性禽流感病毒(Highly pathogenic avian influenza virus,HPAIV)和低致病性禽流感病毒(Low pathogenic avian influenza virus,LPAIV),H5和 H7亚型均属于常见的HPAIV。
自从1996年在广东省鹅体内分离到第一株H5N1亚型AIV[2],H5亚型AIV逐渐在健康水禽中被监测发现,而且随着时间的推移,这些病毒对哺乳动物的感染能力和致病力显著增强[3]。2004年以来国内多地暴发感染AIV,如2005年的我国青海湖野鸟大规模感染H5亚型HPAIV[4],此后又多次报道H5亚型AIV导致野鸟大面积死亡的疫情。同时,该亚型AIV的感染宿主范围逐渐扩大,除了感染家禽和野鸟外,自然界中多种哺乳动物,包括猪、犬、老虎[5-7]等也可以到感染,并常有抗原漂移以及抗原转变的现象[8],甚至对人类的生命健康构成潜在威胁。
随着H5亚型AIV的不断进化和变异,其HA和NA亚型组合愈加丰富,在自然界中可分离到H5N1、H5N2、H5N3、H5N5、H5N6、H5N7、H5N8和H5N9等多种亚型组合的AIV。H5N3亚型AIV最常见的感染宿主是野生水禽和家禽,虽然目前的病毒分离率不高[9],但是与HPAIV相比,H5N3亚型LPAIV感染宿主后由于临床症状不明显,极有可能被忽视,从而给该病毒通过各种机制突变为HPAIV提供机会,因此该亚型病毒对养殖业和人类构成的潜在危害不可小觑。
为了解H5N3亚型AIV的生物学特性,本研究对我国浙江省分离到的一株H5N3 LPAIV进行了全基因组序列分析,利用小鼠为动物模型初步评价H5N3亚型AIV对哺乳动物的感染性,为H5N3亚型禽流感疫情的预警和防控提供相关实验数据支持。
1 材料与方法
1.1 主要实验材料 DK/ZJ/S1368/2017(H5N3)由国家禽流感参考实验室分离、鉴定并保存;6周龄雌性BALB/c小鼠购自北京维通利华实验动物有限公司。
病毒RNA提取试剂盒购自天根生化科技(北京)有限公司;反转录试剂盒购自东洋纺生物科技有限公司;EasyTaqDNA聚合酶购自北京全式金生物科技有限公司;PCR产物胶回收试剂盒购自美国Omega公司;测序反应试剂盒BigDye Terminator 3.1购自美国ABI公司。
1.2 病毒增殖及其鸡胚半数感染量(EID50)的测定将病毒原液用加入青霉素、链霉素、头孢的PBS缓冲液10倍倍比稀释,接种10日龄SPF鸡胚,纯化三代,根据Reed-Muench法计算病毒的鸡胚半数感染量。
1.3 病毒的全基因组序列测定和遗传演化分析 按照RNA提取试剂盒的说明书提取病毒的RNA,以通用引物(5'-AGCA AAAGCAGG-3')为反转录引物,利用反转录试剂盒获得cDNA。参照GenBank中H5亚型AIV全基因组序列,以其非编码区域的保守片段设计引物扩增病毒的8个基因节段,包括HA、NA、碱性聚合酶蛋白1(Polymerase basic protein 1,PB1)、碱性聚合酶蛋白 2(Polymerase basicprotein2,PB2)、酸性聚合酶蛋白(Polymerase acid protein,PA)、核蛋白(Nucleoprotein,NP)、基质蛋白(Matrix protein,M)和非结构蛋白(Non-structural protein,NS)。PCR 反应体系(50 μL):10× TaqPCR Buffer 5 μL,dNTP(10 mmol/L)1 μL,上下游引物(20 pmol)各 0.5 μL,TaqDNA 聚合酶(5 U/μL)0.25 μL,cDNA 1 μL,ddH2O 41.75 μL。PCR 反应程序:94 ℃2 min;94℃ 30 s、52℃ 30 s、72℃ 110 s,35个循环;72℃5 min。PCR产物经琼脂糖凝胶电泳检测,将阳性产物通过胶回收试剂盒回收纯化。然后利用测序试剂盒BigDye Terminator 3.1进行测序,利用SeqMan软件对测序结果进行拼接处理,最终得到完整序列,并对特殊位点的推导氨基酸序列进行分析。利用NCBI数据库在线BLAST对测序结果比对,利用MEGA 7.0软件进行各基因节段进化树的绘制。
1.4 小鼠的感染性实验 将50 μL含有106EID50的病毒稀释液鼻腔接种感染组8只6周龄BALB/c小鼠,对照组5只小鼠每只接种50 μL PBS。感染3 d后从感染组随机选取3只小鼠干冰麻醉后迫杀,取其脑、鼻甲、脾、肾、肺,将脏器匀浆后分别利用鸡胚进行病毒滴定。其余5只从感染日起连续观察14 d,记录小鼠的体质量变化、发病及死亡情况。该试验在中国农业科学院哈尔滨兽医研究所生物安全三级实验室内进行。
2 结果
2.1 全基因组的序列测定 得到病毒株DK/ZJ/S1368/2017(H5N3)全基因组完整序列后通过NCBI的Blast比对分析,并绘制HA基因进化树。结果显示,DK/ZJ/S1368/2017(H5N3)的HA基因和A/Duck/Mongolia/107/2014(H5N7)核苷酸同源性最高(表1),为 98.3%,属于同一分支(图1)。总体来看,DK/ZJ/S1368/2017(H5N3)的8个基因节段来源复杂,分别与NCBI数据库 中 H10N7、H1N1、H6N2、H5N7、H10N3、H3N8等多种不同亚型的AIV的部分基因节段均具有最高核苷酸同源性(表1)。
2.2 氨基酸序列的特殊位点及其进化分析 氨基酸序列分析结果显示,病毒株DK/ZJ/S1368/2017(H5N3)HA蛋白裂解位点处氨基酸序列为PQRETR↓GLF,仅含1个碱性氨基酸,符合LPAIV的分子特征。流感病毒的抗原性除了受到抗原位点处氨基酸序列的影响以外,还受到HA蛋白糖基化修饰的影响。该病毒共有5个潜在糖基化位点,分别位于27 位(NST)、39 位(NVT)、181 位(NNT)、302 位(NSS)、496位(NGT)。受体结合位点及其周围氨基酸的改变可能会改变病毒的受体结合特性,本研究中病毒的HA蛋白第226和第228位点分别为谷氨酰胺(Q)和甘氨酸(G),具有结合禽源受体的分子特征。PB2基因的E627K,D701N的突变可以增加AIV对哺乳动物的致病性和病毒在哺乳动物间的传播能力,但本研究病毒株的PB2蛋白未发生任何突变。
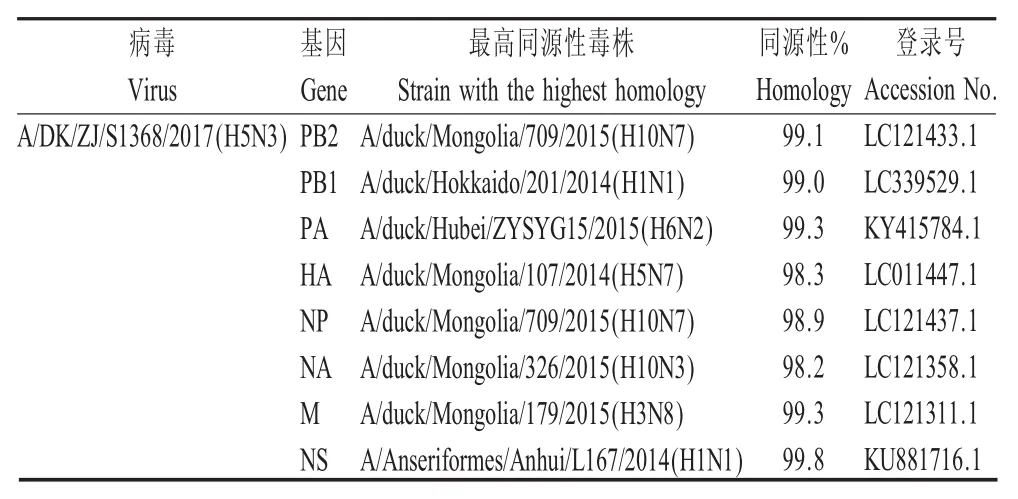
表1 与分离株A/DK/ZJ/S1368/2017(H5N3)的各基因片段最高同源性的病毒株Table 1 BLAST analysis of genes showing the highest homology with that of the A/DK/ZJ/S1368/2017(H5N3)strain
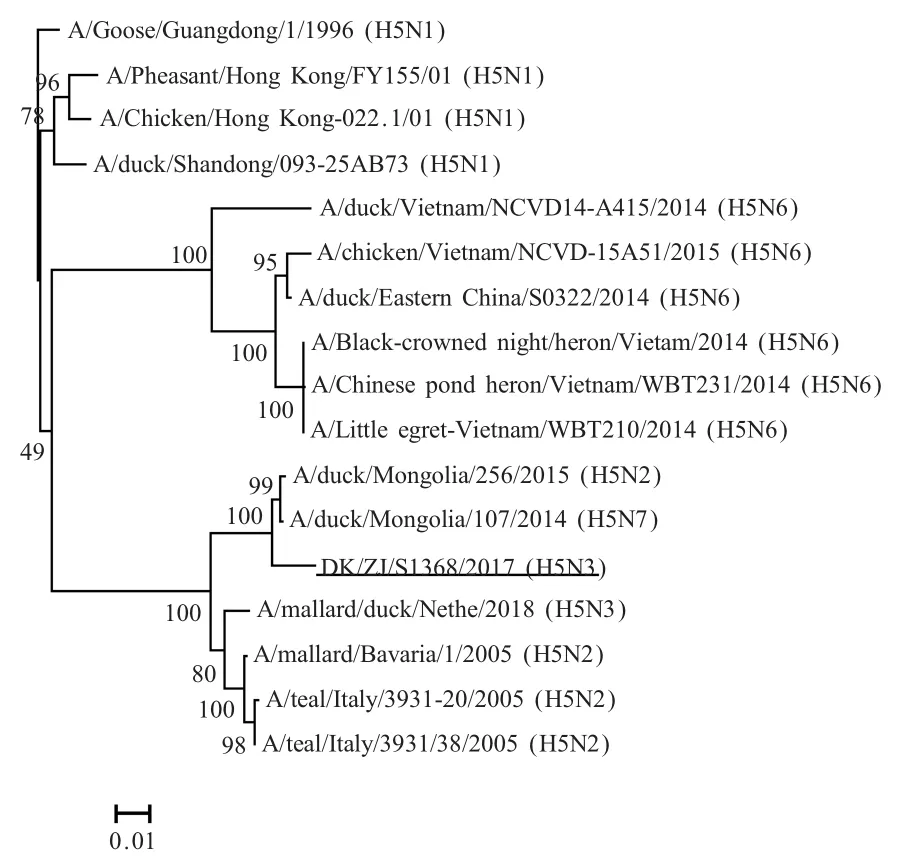
图1 病毒株A/DK/ZJ/S1368/2017(H5N3)的HA基因进化树Fig.1 The phylogenetic tree of HA genes including strain of A/DK/ZJ/S1368/2017(H5N3)
2.3 病毒对BALB/c小鼠的感染性实验结果 小鼠鼻腔接种病毒株DK/ZJ/S1368/2017(H5N3)后,在14 d的观察期内,未观察到小鼠有任何的临床症状,同时该病毒也未引起小鼠体质量下降(图2)。感染后3 d随机迫杀3只小鼠,无菌采集组织样品进行病毒滴定,结果显示,DK/ZJ/S1368/2017(H5N3)AIV病毒株可以在小鼠的肺脏和鼻甲内有效复制,病毒滴度分别为3.83 log10EID50/mL和1.50 log10EID50/mL,但在肾、脑、脾中检测不到病毒(图3),表明病毒不能在肾、脑、脾中有效复制。上述结果表明该病毒对小鼠表现为低致病力。
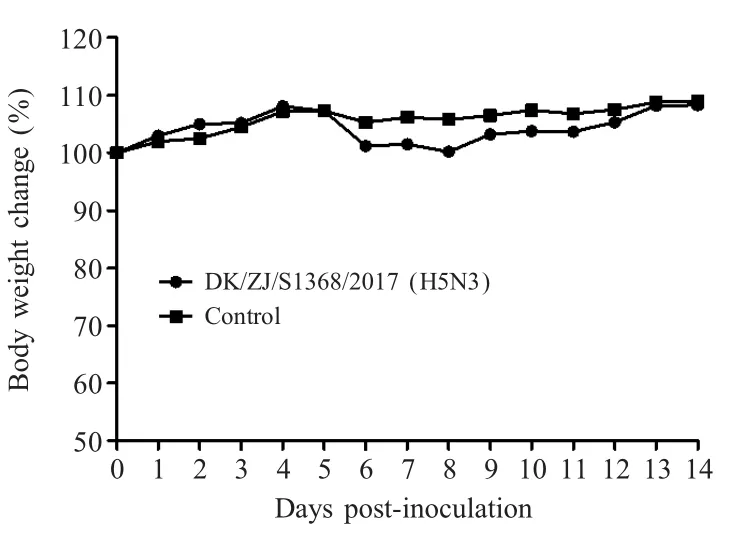
图2 小鼠感染病毒后的体重变化Fig.2 Weight change of mice after inoculated with the A/DK/ZJ/S1368/2017(H5N3)virus
图3 感染病毒3 d后BALB/c小鼠脏器病毒滴定结果Fig.3 Virus titration in mouse organs at 3 days post infection with the A/DK/ZJ/S1368/2017(H5N3)virus
3 讨论
由于AIV的人畜共患性质,全球对其预防和控制尤为关注。一般情况下分离的H5亚型AIV多为HPAIV,但近年来许多H5和H7亚型AIV LPAIV被分离鉴定报道。2013年在中国暴发的H7N9疫情提示LPAIV也可以导致严重感染,甚至导致人类死亡[10]。因此,必须时刻警惕LPAIV对人类构成的潜在威胁。家禽感染LPAIV后,临床症状不明显,很容易被忽视,但经过在宿主体内复制、循环,极有可能通过各种机制突变为HPAIV。本研究中DK/ZJ/S1368/2017(H5N3)是于2017年春季从我国浙江省温州市珊溪镇养殖场分离得到的,特殊位点分析结果显示,该病毒株的HA蛋白裂解位点处氨基酸序列为PQRETR↓GLF,含1个碱性氨基酸,符合LPAIV的分子特征。PB2蛋白的E627K,D701N的突变可以增加AIV对哺乳动物的致病性和病毒在哺乳动物间的传播能力,而经序列分析发现,本研究中病毒的PB2蛋白无上述突变。遗传演化分析结果表明,该病毒的8个基因节段基因来源复杂,内部基因与不同时期分离到的不同亚型的AIV,如H10N7、H1N1、H6N2、H5N7、H10N3、H3N8 等亲缘关系密切,表明该病毒株基因节段来源的多样性。
AIV在不同哺乳动物体内经过循环复制可能会发生突变,使得LPAIV有可能突变为HPAIV。小鼠是被广泛使用于评价AIV对哺乳动物感染性差异的动物模型。本研究中的病毒可以感染小鼠,并可以在小鼠的肺脏和鼻甲内有效复制,但是未能引起小鼠明显的临床症状和体质量下降,更不能导致小鼠死亡。上述实验结果表明这株H5N3病毒基因来源复杂,具有一定感染哺乳动物的能力。
流感病毒的进化机制主要包括突变和重组两种方式。与突变相比,AIV的重组是产生人类流感大流行的有效手段,自上个世纪初期以来4次人类流感大流行中有3次是由重配病毒引起的[11-12]。20世纪90年代后期,出现含有经典猪H1N1病毒的NP、M、NS片段、北美禽源病毒的PB2、PA片段以及季节性人源H3N2的HA、NA、PB1片段的三重重配H3N2病毒[11],这些病毒随后与经典猪源H1病毒和人源H1病毒重配产生H1N1、H1N2病毒导致流感的大流行[12]。大约在2009年,又出现四重重配的H1N1病毒流行事件[13]。2013年,中国上海和安徽省发现人类感染的H7N9病毒的6个内部基因均来源于 H9N2病毒[14]。本研究中 H5N3亚型 AIV的PB2基因、PB1基因、PA基因、NP基因、NA基因、NS基因分别与多种其它亚型AIV有着密切的关系,这些节段可能参与了H5N3亚型AIV的重组。LPAIV,尤其是H5和H7亚型AIV,能够获得突变而成为HPAIV,不仅严重危害家禽养殖业,更对人类公共卫生安全构成潜在威胁。因此H5亚型LPAIV的存在应引起高度关注,加强对H5N3亚型AIV的监测和相关生物学特性的研究具有十分重要的公共卫生学意义。
猜你喜欢
铁道建筑技术(2022年10期)2022-10-28
中国典型病例大全(2022年12期)2022-05-13
中国典型病例大全(2022年9期)2022-04-19
皮肤病与性病(2021年3期)2021-07-30
故事作文·高年级(2019年10期)2019-10-16
中国兽药杂志(2019年4期)2019-05-15
筑路机械与施工机械化(2014年3期)2014-03-01
中学生数理化·七年级数学北师大版(2008年6期)2008-10-22
