
生态因子对大理苍山种子植物多样性分布格局的影响
2019-08-30 07:10:54尹志坚李新辉
西南林业大学学报 2019年5期
杨 涛 尹志坚 李新辉
( 1. 西南林业大学生态与水土保持学院,云南 昆明 650233;2. 国家林业局昆明勘察设计院,云南 昆明 650216)
物种是构成生物多样性的基本单元,也是探讨物种多样性演变、自然资源和生态功能发挥作用的基本单位[1],物种α多样性衡量了生境内或者样方内物种的组成情况,物种丰富度和Shannon多样性指数是经常使用的衡量指标。大部分的研究表明,物种丰富度与海拔之间呈现先上升后下降的单峰曲线格局[2],这些格局的形成很可能与降水量、温度、地形等因素有关系[3]。
物种Beta多样性则是用来表征物种组成在时空尺度上的变化,即不同区域、样方或时间之间的物种变化的量度。一般情况下,环境的异质性越高,β多样性也会越高。β多样性的衡量方法很多,如Bray-curtis指数、Jaccard相异指数等,它主要与降水量差异、温度变化、地形差异、地理距离等因素有关[4-5]。
研究物种α和β多样性与环境因子之间的关系、以及内在的影响因素是当前生物多样性研究的重点之一。本研究关注苍山地区植物物种α多样性和β多样性的变化样式及其影响因素,研究了植物多样性与年平均降水量、年平均温度、坡度、坡向、岩石类型和土壤类型等因子的关系,分析它们对植物多样性的解释能力,推测本研究区植物多样性的形成原因和维持机制。
1 研究区概况
苍山位于云南省大理白族自治州境内,由19座海拔3 074~4 122 m的山峰组成,地跨大理市、洱源县和漾濞县3县(市),地处东经99°55′~100°12′、北纬 25°34′~26°00′,最高峰马龙峰(4 122 m)与东坡的大理湖盆(1 966 m)相对高差2 156 m[6]。
苍山处于低纬度高原,气候属亚热带高原季风气候,分为旱雨两季。年均气温15.1 ℃,年平均降水量1 000~1 100 mm,相对湿度65%[7]。山麓到山顶分布有亚热带、暖温带、温带以及寒温带。
2 研究方法
2.1 数据来源
植物物种以及各种植物分布的海拔范围信息是本研究的基础,根据《大理苍山种子植物区系的研究》[8],删除没有海拔记录的物种,建立参与分析的种子植物名录,共有种子植物131科596属约1 519种,并录入其分布海拔范围等信息数据建立物种数据库。
年平均温度和年平均降水量的数据来源于网站 http://www.worldclim.org/;DEM 数 据 (Digital Elevation Model)来自于 http://gdem.ersdac.jspacesystems.or.jp/feature.jsp,分辨率为30 m×30 m。本研究所使用的土壤类型和岩性类型数据来源为http://www.isric.org/data/data-download。
2.2 分析方法
2.2.1 地形分异特征
首先,由低海拔到高海拔,根据苍山地区的海拔特征可将该区分为若干个海拔段。最高峰马龙峰4 122 m,东坡大理湖盆1 966 m,所以从2 000 m开始划分海拔段,同时由于苍山地区海拔在4 000 m以上的面积比较小,因此将4 000 m以上单独作为1个海拔段。本研究从2 000 m开始,每100 m为1个海拔段,共21个海拔段。
2.2.2 环境因子及地形因子计算
为了研究苍山地区植物群落与环境因子(以年平均降水量和年平均温度为代表)之间的关系,从 WorldClim v1.4(http://www.worldclim.org)这一数据库中获得每个海拔段内各个点的降水量、温度等数据,并求得各海拔段内的年平均降水量、年平均温度。环境因子之间差异则是通过对各海拔段内年平均降水量、年平均温度,运用R软件fossil软件包[9]计算得出。
DEM以及由此生成的DEM模型能够容易地提取坡向、坡度等各种地形特征参数,同时能够对具有不同地形特征的区域面积进行计算[10]。为此基于研究区的DEM模型以及土壤和岩石图层,利用ArcGIS,在每一个海拔段内,计算每一度的坡度(0°~90°)所占有的面积、每一度坡向的面积(0°~259°,以正南为 0°)、每一种土壤类型及每一种岩石类型所占有的面积。使用Shannon公式计算每一个海拔段内的岩石、土壤、坡度、坡向的α多样性指数,同时使用Braycurtis公式计算各海拔段之间的岩石、土壤、坡度、坡向的Bray-curtis指数。将年平均降水量、年平均温度、岩石、土壤、坡度、坡向、坡位或者相关的差异作为解释变量本别对Shannon多样性和Beta多样性进行分析。
为了分解物种多样性矩阵总的变异,将坡向、坡度、岩性、土壤、年平均温度差异、年降量差异这6个因子经过packfor[11]软件包中的forward.sel函数对因子进行筛选,保留相关性显著的因子。然后将这些因子使用vegan软件包中的varpart这一函数进行进行进一步的分析,该方法可以将一个变量的变异划分为2~4个解释变量的解释力及组合,各个因素解释力的大小则是用RDA分析校正了的R2来衡量的[12]。
2.2.3 物种多样性测度方法
1)α多样性测度方法。Shannon指数:
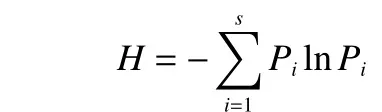
式中:Pi为种i的相对比例,s为物种数目。
2)β多样性测度方法。
Bray-curtis指数(CN):
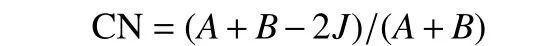
式中:A为样地a的物种数;B为样地b的物种数;J为样地a和b共有物种数。
3 结果与分析
3.1 属、种丰富度随海拔梯度的变化
通过计算在不同的海拔段属、种的丰富度发现,2个分类群水准的丰富度具有相似的变化规律;属、种丰富度与海拔梯度均呈现单峰曲线关系,在低海拔区域,随着海拔的升高,丰富度呈逐渐上升趋势,而在高海拔区域随海拔的升高,丰富度呈逐渐下降趋势,即在中间海拔区域丰富度最高,其中,物种丰富度的最大值出现在海拔2 500 m左右,属的丰富度的最大值出现在2 400 m左右(图1)。
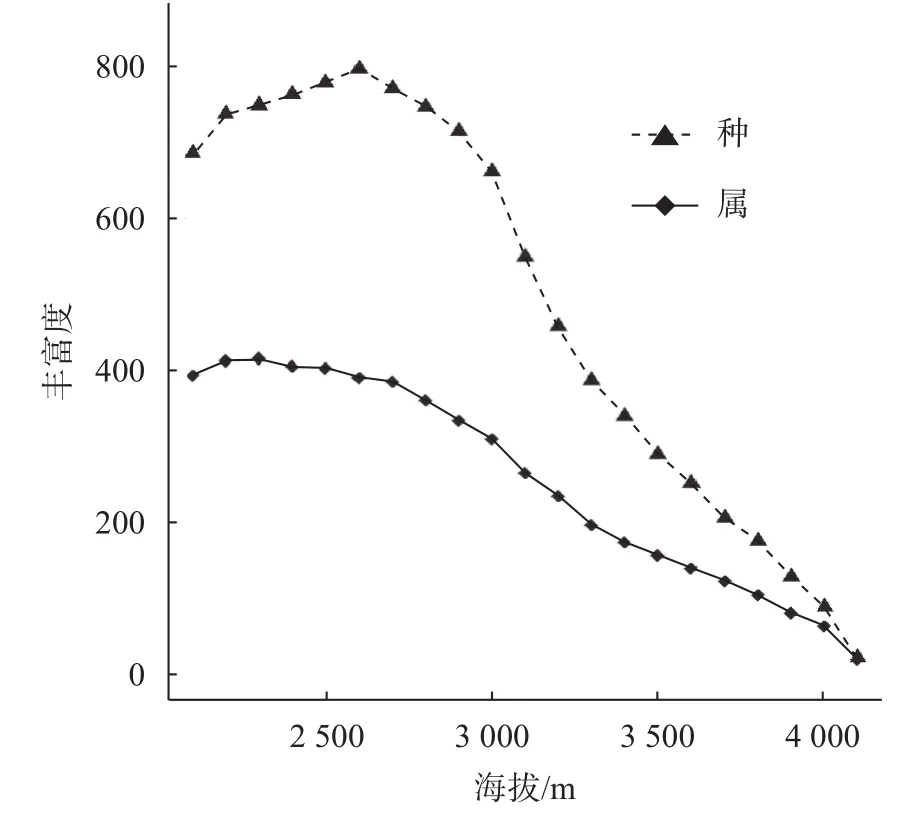
图 1 苍山地区种子植物属和种的丰富度沿海拔梯度变化Fig. 1 The genus and species richness of seed plants in Cangshan area vary along the elevation gradient
苍山地区沿海拔森林类型依次为暖性云南松林(2 000 m左右)、半湿性暖性常绿阔叶林(2 500 m左右)、温凉针阔混交林(2 800 m左右)、温凉针叶林(3 100 m左右)、苍山冷杉林(3 500 m左右)、杜鹃灌木林(3 800左右)以及杜鹃灌丛和草甸(4 000 m以上)直至马龙山顶[13]。在各海拔带中菊科(Asteraceae)植物优势明显,各海拔带内也均有优势明显的属(表1)。
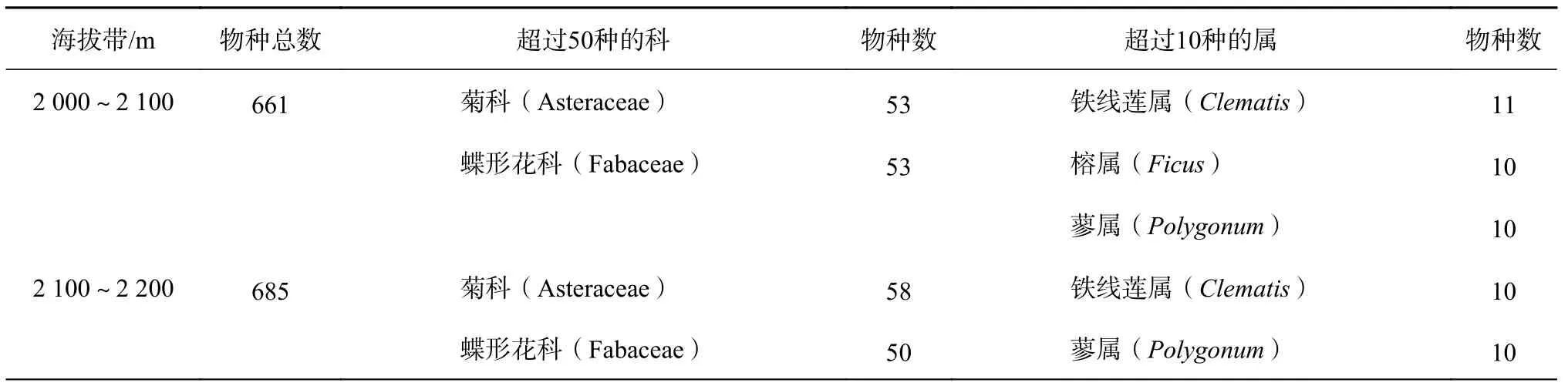
表 1 苍山地区各海拔带内物种数以及主要科属统计Table 1 Number of species and major families in various altitude zones in Cangshan area

续表 1

续表 1
3.2 α 多样性
由Pearson相关矩阵不仅可以看出α多样性与环境因子之间的变化关系,还可知Shannon多样性指数与年平均降水量、年平均温度、岩性相关性最大(图2)。其中,与年平均温度和岩性呈正相关,其Pearson相关系数分别为0.872(P<0.01)和0.835(P<0.01);与年平均降水量呈负相关,相关系数为-0.893(P<0.01)。
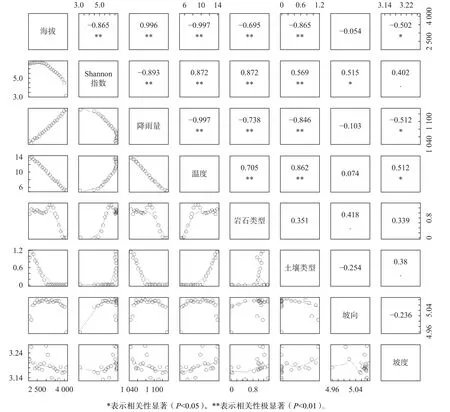
图 2 Shannon多样性指数与各环境因子之间、及各环境因子之间的相关性Fig. 2 Correlation between Shannon diversity index and various environmental factors, and the correlation among environmental factors
经过因子筛选,平均降水量和岩石类型的组合对Shannon多样性的解释量最大,年平均降水量对α多样性的解释比例为79%;坡向对α多样性的解释比例为68%,两者共同解释比例为85%(图 3)。

图 3 年平均降水量、岩石类型对α多样性的解释比例Fig. 3 Interpretation of the annual average precipitation and lithology to α diversity
3.3 β 多样性
苍山地区β多样性与各环境因子之间均呈显著的相关性,β多样性与各环境因子之间、各环境因子之间均呈正相关(图4)。其中,β多样性与各环境因子之间的相关性中,Pearson相关系数最大的为年平均降水量、年平均温度、岩性,分别 为 0.912( P<0.01)、 0.908( P<0.01)、 0.885(P<0.01)。

**表示相关性极显著(P<0.01)。图 4 β多样性与各环境因子之间、各环境因子之间的相关性Fig. 4 Correlation between β diversity and various environmental factors, and the correlation among environmental factors
年平均降水量变化、岩性差异组合对β多样性的解释量最大,年平均降水量变化对β多样性的解释率为83%,岩性差异对β多样性的解释率为78%,两者共同作用的解释率为89%(图5)。
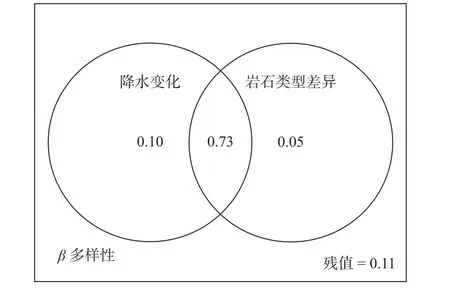
图 5 年平均降水量、岩性对β多样性的解释比例Fig. 5 Interpretation ratio of annual average precipitation and lithology to β diversity
4 结论与讨论
物种丰富度是表示物种多样性最真实、最客观的指标之一[14]。大多数的研究表明中海拔水平拥有最高的位置丰富度[15-17]。在本研究中,物种丰富度随海拔的升高呈现单峰曲线关系,也就是说中海拔物种丰富度水平最高,属的丰富度也显示了相近的趋势。苍山地区种子植物的物种丰富度随海拔的升高呈现先增加后减少的分布格局,在海拔2 500 m左右物种丰富度最大。海拔梯度的变化事实上对应着降水和热量条件的梯度变化[18-19],所以海拔的影响事实上响应了气候因子的综合影响[20-21]。在大多情况下,低海拔的湖盆地区(2 100 m以下)受河谷气候条件和特殊的地形环境影响,这一区域温度较高而降水相对较少,植物生长季节所需的热量充足而水分常相对缺少,而且这一区域受人为干扰程度较大;相反,在高海拔地段水分虽然充足,但植物分布的主要限制因子或许又变成了热量,该区域的寒冷气候使土壤生成和植物生长缓慢,加之又有各种自然灾害,严酷的环境超出了绝大多数物种生存所能够忍受的极限,故而物种也较少。相比于低海拔和高海拔区域,在中海拔地段可能是热量和水分分配最适合的生境,是两个极端区域植物种类分异的过渡区域,再加上人为干扰较少,从而使资源的可利用率提高,因此中海拔区域物种多样性较高[22]。
群落或生境的物种结构及组成的复杂程度能够用Shannon多样性指数来全面反映。在本研究中,采用Shannon多样性指数研究苍山地区种子植物α多样性,它与物种丰富度一样,随海拔上升呈现相似的单峰曲线关系。通过因子的选择和方差分析表明对Shannon多样性指数解释力最大的组合是年平均降水量和岩石类型两个因子,而不是显著性最高的2个因子组合(年平均降水量、年平均温度),这是由于因子之间的共线性关系导致的,正如年平均降水量和年平均温度之间有明显的共线性关系,而年平均降水量和岩石类型之间没有显著的共线性关系,而且通过因子的选择发现,除了年平均降水量这个最大的因素外,岩石类型的贡献最大而达到显著,故它们的组合能解释研究区最多的种子植物α多样性,联合达到了85%。
β多样性的格局是由样方或区域之间的环境或者地理差异作用下形成及维持的。本区域的种子植物β多样性与年平均降水量差异、年平均温度差异、岩石类型差异、土壤类型差异、坡度差异和坡向的差异都有极显著的关系,说明这些因子差异都在一定程度上影响了研究区β多样性的形成或维持,其中与年平均降水量差异的关系最大。而经过因子的选择和方差分析表明对β多样性指数解释力最大的组合是降水和岩石类型的差异,而不是降水差异和温度差异两个显著性最高因子组合,这是由于降水差异和温度差异两者具有显著的共线性关系,在去除降水差异对β多样性影响之后,温度差异对β多样性将不具有显著的影响,而是岩石类型的差异对其有显著的影响。
苍山地区各海拔带之间的β多样性形成和维持机制主要是由于环境因子的差异,即符合生态位理论的预测。各海拔带内环境因子的差异导致处在这一生态位的物种之间的差异[23-24],如碟形花科、榕属、铁线莲属等科属需要较高的温度,使它们在相对低海拔带优势明显;而菊科、蓼属等科属由于生态位宽度较宽使它们在各海拔带均优势明显;而杜鹃花科、紫堇属、杜鹃属等科属适应不了高温,对降水量要求更高所以主要分布于较高的海拔带。另有中性理论认为群落内物种生物学特征类似时,由于扩散限制也能产生相似度随地理距离增加而减小的格局,即地理距离就能很好地解释β多样性。本研究由于无法取得各海拔段之间的实际距离,故而也就无法衡量中性理论对本区垂直海拔带的物种β多样性的作用,但是用各海拔带之间的垂直距离差异作为解释变量时,虽然与物种β多样性具有显著的相关,但是做因子筛选时被筛除。而且对物种β多样性解释量最大的组合是属于环境因子的组合,达到了89%,所以本区垂直方向上的β多样性格局符合生态位理论的预测,生态位过程对本区的β多样性形成和维持起着主要的作用。
人为干扰程度的增加,对生物的生境造成威胁,影响了一些物种的正常生存和繁衍,对区域生物多样性造成了威胁[25],加之在全球气候变化的背景下,对生物多样性热点地区或是生态环境性脆弱地区进行生物多样性相关研究,对生物多样性保护、保护区建立以及保护方案的制定能提供理论依据,意义重大。生境异质性对多样性格局的影响力可能在一定程度上取决于植物的生物地理分布区性质[26],本研究表明研究区域物种丰富度、属的丰富度以及物种的Shannon多样性随海拔都呈现出先增加后减小的单峰曲线格局,这与随海拔而发生变化的环境因子有密切的关系,其中年平均降水量和岩石类型的组合能最大地解释本区种子植物α多样性格局。β多样性随环境距离的增大而增大,降水差异和岩石类型差异对β多样性具有最大的解释量组合,生态位对本区β多样性形成和维持起主要作用。
[ 参 考 文 献 ]
[1]Mallet J. Species: the units of biodiversity [J]. Trends in Ecology & Evolution, 1997, 12(11): 453-454.
[2]Rahbek C. The relationship among area, elevation, and regional species richness in neotropical birds [J]. The American Naturalist, 1997, 149(5): 875-902.
[3]McCain C M, Grytnes J A. Elevational gradients in species richness[M]. Chichester: Encyclopedia of Life Sciences (ELS), 2010.
[4]Arnan X, Cerdá X, Retana J. Partitioning the impact of environment and spatial structure on alpha and beta components of taxonomic, functional, and phylogenetic diversity in European ants [J]. Peer J, 2015, 3: e1241.
[5]De Caceres M, Legendre P, Valencia R, et al. The variation of tree beta diversity across a global network of forest plots [J]. Global Ecology and Biogeography,2012, 21(12): 1191-1202.
[6]云南省林业调查规划院大理分院. 云南省大理市森林资源规划设计调查报告[R]. 大理: 云南省林业调查规划院大理分院, 2005.
[7]穆静秋. 大理苍山生物多样性现状与保护措施 [J]. 林业调查规划, 2006, 31(1): 79-82.
[8]尹志坚. 大理苍山种子植物区系的研究[D]. 北京: 中国科学院研究生院, 2012.
[9]Vavrek M J. Fossil: palaeoecological and palaeogeographical analysis tools [J]. Palaeontologia Electronica,2011, 14(1): 1-16.
[10]邬伦, 刘瑜, 张晶, 等. 地理信息系统: 原理、方法和应用[J]. 北京: 科学出版社, 2001.
[11]Chamberland J M, Lanthier G, Boisclair D. Comparison between electrofishing and snorkeling surveys to describe fish assemblages in Laurentian streams [J]. Environmental Monitoring and Assessment, 2014, 186(3):1837-1846.
[12]Peres-Neto P R, Legendre P, Dray S, et al. Variation partitioning of species data matrices: estimation and comparison of fractions [J]. Ecology, 2006, 87(10):2614-2625.
[13]曾觉民. 云南自然森林分类系统及地理分布研究 [J].西南林业大学学报(自然科学), 2018, 38(6): 1-18,231.
[14]唐志尧, 方精云. 植物物种多样性的垂直分布格局 [J]. 生物多样性, 2004, 12(1): 20-28.
[15]Lieberman D, Lieberman M, Peralta R, et al. Tropical forest structure and composition on a large-scale altitudinal gradient in costa rica [J]. The Journal of Ecology, 1996, 84(2): 137-152.
[16]Rahbek C. The elevational gradient of species richness:a uniform pattern? [J]. Ecography, 1995, 18(2):200-205.
[17]刘彬, 布买丽娅木·吐如汗, 艾比拜姆·克热木, 等. 新疆天山南坡中段种子植物区系垂直分布格局分析 [J]. 植物科学学报, 2018, 36(2): 191-202.
[18]Marrs R H, Proctor J, Heaney A, et al. Changes in soil nitrogen-mineralization and nitrification along an altitudinal transect in tropical rain forest in costa rica [J].The Journal of Ecology, 1988, 76(2): 466-482.
[19]Tian Z H, Bai H Y, Su K, et al. Reconstruction and response of tree-ring width chronology at various altitudes to climate change on Taibai Mountain[C]//Proceedings of the 6th International Conference on Informatics, Environment, Energy and Applications, New York: ACM, 2017: 60-66.
[20]Whittaker R H. Vegetation of the Siskiyou mountains,Oregon and California [J]. Ecological Monographs,1960, 30(3): 279-338.
[21]陈超男, 朱连奇, 田莉, 等. 秦巴山区植被覆盖变化及气候因子驱动分析 [J]. 生态学报, 2019, 39(9): 1-9.
[22]王国宏. 祁连山北坡中段植物群落多样性的垂直分布格局 [J]. 生物多样性, 2002, 10(1): 7-14.
[23]董冬, 许小天, 周志翔, 等. 安徽九华山风景区古树群落主要种群生态位的动态变化 [J]. 生态学杂志,2019, 38(5): 1292-1304.
[24]邢亚蕾, 魏天兴, 葛根巴图. 鹫峰国家森林公园残次林物种多样性及生态位特征 [J]. 植物研究, 2015,35(6): 915-922.
[25]杨绣坤. 自然保护区对保护生物多样性的意义 [J]. 农业与技术, 2018, 38(6): 253.
[26]冯建孟, 胡小康. 环境因子对滇西北地区植物多样性分布格局的影响 [J]. 信阳师范学院学报(自然科学版), 2019, 32(01): 62-68.
猜你喜欢
《学习方法报》地理商务星球七年级(2023年13期)2023-10-11 02:44:46
环球时报(2023-03-21)2023-03-21 19:17:23
农业工程技术(2021年25期)2021-12-06 09:08:02
创作评谭(2020年4期)2020-07-18 15:31:16
启蒙(3-7岁)(2019年8期)2019-09-10 03:09:08
中国诗歌(2018年6期)2018-11-14 13:24:12
金桥(2018年9期)2018-09-25 02:53:28
时代农机(2018年2期)2018-05-21 07:45:10
江西农业(2018年23期)2018-02-11 07:26:59
连环画报(2016年7期)2017-01-20 22:01:18
