
外来植物肿柄菊结籽量影响因素分析
2019-08-22 02:08:32王四海陈剑李宁云孙卫邦吴超
生态环境学报 2019年7期
王四海 ,陈剑,李宁云,孙卫邦,吴超
1. 云南省林业科学院/云南省森林植物培育与开发利用重点实验室/国家林业局云南珍稀濒特森林植物繁育和保护重点实验室,云南 昆明 650201;2. 西南林业大学/国家高原湿地研究中心,云南 昆明 650224;3. 中国科学院昆明植物所/云南省极小种群野生植物综合保护重点实验室,云南 昆明 650204
外来植物在入侵过程中与环境之间存在一系列的动态反馈作用(Yelenik et al.,2013),对新环境的适应性变化能反映植物入侵的潜能(Colautti et al.,2013;Clark et al.,2013;Alexander et al.,2019)。繁殖体压力是入侵植物在新环境中群落建立的关键性因素,种子的长距离扩散是入侵植物侵占新领地的主要途径,种子量在某种程度上决定着入侵植物在新环境中成功建立的可能性(Davies et al.,2007;Davies,2008)。对于植物本身而言,种子量有单位面积植株数、分枝数、开花(序)量、结实量、结籽率等多种因素共同作用,这些决定种子产量的构成因素受生长环境的多方面影响(Winn et al.,1987)。在局域尺度范围内(相同气候类型),因立地条件的差异入侵植物的群落结构特征变化影响着结籽量;在区域尺度范围(不同气候类型),温度、降雨等多种气候环境因子影响着植物结籽量,入侵植物的种子产量会随着环境梯度变化而改变,特别是有些外来入侵植物到达分布区边界时很难产生能够繁育的种子(Colautti et al.,2013;Groeneveld et al.,2014;徐高峰等,2014;Hulme,2017)。从局域尺度和区域尺度上查清入侵植物结籽量变化规律和影响因素,可以了解在不同环境条件下入侵植物的扩散潜能和发展动态,对于外来入侵植物的预警和预防都有重要意义。
肿柄菊[Tithonia diversifolia (Hemsl.) A. Gray]是菊科肿柄菊属植物,原产于墨西哥及中美洲地区,曾作为观赏植物、绿肥和防止土壤侵蚀植物被广泛引种到亚洲、非洲、北美和澳洲的许多国家和地区。目前在东南亚、非洲、太平洋的一些热带和亚热带地区成为入侵杂草(Yang et al.,2012)。肿柄菊在热带和亚热带为丛状生长的灌木或亚灌木,在入侵地区形成单优群落。在我国的云南、广西、广东、福建、海南、台湾已普遍生长,部分地区已造成严重危害,并且有向更高纬度和海拔快速扩散的趋势(王四海等,2004;罗瑛等,2009;朱枫等,2018)。肿柄菊能通过靠近地表的茎生出不定根进行克隆繁殖,克隆繁殖植株只能在母体附近,主要能增加群落的密实度和向外蔓延(王四海等,2004),肿柄菊新群落的建立要依靠种子的较远距离传播,因此种子量也就决定了肿柄菊建立新群落能力。丛密度、枝密度、每枝果序数等群落特征都是种子产量的构成因素,群落特征差异必然会造成结籽量的不同,肿柄菊的单株结实量具有较大差异(Muoghalu et al.,2005),但是肿柄菊群落结构如何影响结籽量还缺乏研究。入侵植物在跨气候类型扩散过程中,由于温度、降雨、海拔等多种环境因素的改变导致开花物候、花形态、传粉效率会发生一系列变化,从而结籽量也发生着改变(Colautti et al.,2013;Groeneveld et al.,2014;王颖洁等,2017)。肿柄菊的果序直径、每果序结实量、结籽率在区域尺度范围具有较大差异(王四海等,2008),但对于这种差异的规律性还缺乏了解,对造成差异的主要环境因子也并不清楚。本文探讨肿柄菊群落的丛密度、枝密度、每丛分枝数、每枝果序数等群落特征对果序产量的影响规律;在区域尺度上查明肿柄菊每果序结实量和结籽率沿环境梯度变化规律,以及影响结实和结籽的主要环境因素。通过肿柄菊在局域和区域尺度上结籽量变化规律的研究,本文旨在为肿柄菊扩散潜力的预测和预防提供科学依据。
1 研究方法
1.1 研究区域概况
研究区域选择在云南有肿柄菊分布的 101°-103°E之间纵跨南北的条带区域,该区域纬度变幅21°10′-26°30′N(图 1),海拔变幅 114-4247,年降雨量变幅 578-2280 mm,5-10月为雨季,11月至次年4月为旱季。该区域不仅有北热带、南亚热带、中亚热带、北亚热带、南温带至中温带的连续气候类型,同时由于受复杂地形和大气环流的影响,总体上各种气候类型呈现出相互镶嵌的特点,影响植物分布的两个主要因素温度和降雨组合类型多样(陈宗瑜,2001),这种自然条件下肿柄菊对环境的多种适应性关系能够更好呈现。

图1 采集点及最近气象站的位置Fig. 1 The locations of sampling sites and the nearest meteorological stations
1.2 群落调查和特征统计
为了探讨在局域尺度范围内肿柄菊群落结构特征对果序量的影响,在2016年和2017年的1-2月肿柄菊花果期,在云南玉溪市和普洱市肿柄菊分布最为普遍的海拔段(900-1300 m)(朱枫等,2018),随机选取人为干扰较少肿柄菊群落分布典型地段(图1),在群落内设置4 m×4 m的样方,在每个样方内逐枝采集丛状生长的分枝(分枝处距地面<20 cm),并计数每枝的果序量,记录丛、分枝、果序之间的从属关系。为了反映群落果序量和群落特征是适应不同环境的随机相应变化,每个样方为独立设置,没有重复,共设置 20个样方,其中在玉溪市的新平县设置 16个样方,普洱市的思茅区设置4个样方(图1)。并统计每个样方的主要特征,丛密度=样方内的植株丛数/样方面积,枝密度=样方内所有分枝数/样方面积,每丛枝数=样方内所有分枝数/样方内的植株丛数,每枝果序数=样方内果序总数/样方内所有分枝数,果序密度=样方内果序总数/样方面积。
1.3 果序采集和结实量测量
为了探讨在区域尺度范围肿柄菊结籽量的变化规律,在1.1研究区域内从南至北选取了有肿柄菊分布的18个小地点,18个地点覆盖了从热带至北亚热带(目前肿柄菊分布的最冷区域)的各种气候类型(图1)。在2016年的1-2月肿柄菊果实成熟季节采集果序,并记录采样点海拔和经纬度。每个地点至少采集 30个植株个体的果序,在每个地点的采集袋中随机选 30个果序,统计每头果序的结实量(全部瘦果)和结籽量(含有种子的瘦果,即饱满的瘦果),并统计每个地点每果序平均结实量和结籽量,以及结籽率。每果序平均结实量为每个地点 30头果序结实量的平均值,每果序平均结籽量为每个地点 30头果序结籽量的平均值,结籽率=每果序平均结籽量/每果序平均结实量。
1.4 主要环境数据获取
果序采样点温度数据是通过最近气象站的数据推导获得,根据海拔每上升 100 m温度降低0.6 ℃的规律,具体公式如下:TSS=TMS-0.6(ASSAMS)/100,TSS是采样点的温度,TMS是最近气象站的温度,ASS是采样点的海拔,AMS是气象站的海拔(图1)。降雨数据使用最近气象站降雨数据。温度和降雨数据都是 30年的平均值(云南省气象局,1982)。海拔和经纬度数据在采样点用GPS现场采集。
1.5 数据的统计分析
用SPSS 16.0进行Pearson相关分析,分别分析了群落果序密度与群落丛密度、枝密度、每丛分枝、每枝果序数之间的相关关系,以及与果序密度有相关性的群落特征因素之间的相关关系;同时分别分析了结实量和结籽率与年均温、1月均温、7月均温、年降雨量、雨季降雨量和旱季降雨量,以及纬度和海拔的相关关系。每果序平均结实量和结籽量用SPSS 16.0进行单因素方差分析。相关关系图和统计图用Microsoft Excel 2010绘制。
2 结果与分析
2.1 群落特征对果序量的影响
丛状生长的肿柄菊为头状花序,花序成熟后形成果序,果序的多少直接影响到结实量。肿柄菊群落每平方米果序量从60-300余头不等,果序密度与群落丛密度和每丛平均分枝数没有显著的相关关系(P>0.1);果序密度与群落枝密度有极显著的正相关关系(P<0.01),与每分枝果序数呈显著的正相关关系(P<0.05)(图 2)。这表明丛状生长的肿柄菊,群落的丛密度和每丛分枝之间可有多种不同的状态,丛密度和每丛分枝数都不会单独影响群落果序量,群落总体枝密度对群落果序量有着明显的影响,其次是每分枝的果序量。
进一步分析发现,肿柄菊群落枝密度与丛密度有极显著的正相关关系(P<0.01),枝密度与每丛的分枝数没有相关性(P>0.1)(图3);每分枝的果序数与群落丛密度和枝密度都没有相关性(P>0.1)(图 4)。综合以上结果可以看出,肿柄菊群落产生的果序量由肿柄菊丛密度、枝密度和每分枝果序量等群落特征共同决定。
2.2 不同地点间每果序结实量和结籽量的差异
被选择的 18个采样地点平均每果序结实量148.4-196.4粒,在采样点间每果序的结实量具有极显著差异(P<0.001),但是变异系数仅为8.1%。然而,在采样点间平均每果序结籽量0.8-98.1粒,变异系数为 82.7%,结籽率为 0.43%-55.1%,每果序的结籽量具有更大的差异(P<0.001)(图5,表 1)。尽管每果序结实量和结籽量在采样点间都具有极显著的差异,但是每果序结籽量的变化幅度显然要大的多,这也表明每果序结籽量更易受环境变化的影响。
2.3 主要环境因素对结实量的影响
肿柄菊每果序平均结实量与年均温和 1月均温有微弱的负相关关系(P<0.1),与7月均温没有相关关系(P>0.1),其它与年降雨量、雨季降雨量、旱季降雨量、纬度和海拔均没有显著相关关系(P>0.1)(图 6)。肿柄菊每果序结实量有随着分布区温度降低结实量增加的趋势,但这种趋势并不显著;每果序结实量不受降雨量、纬度和海拔的显著影响。

图2 肿柄菊群落果序密度与丛及丛分枝关系Fig. 2 Relationships between capitulum density, and cluster and branch in Tithonia diversifolia community

图3 肿柄菊群落枝密度与丛特征关系Fig. 3 Relationships between branch density and cluster in Tithonia diversifolia community
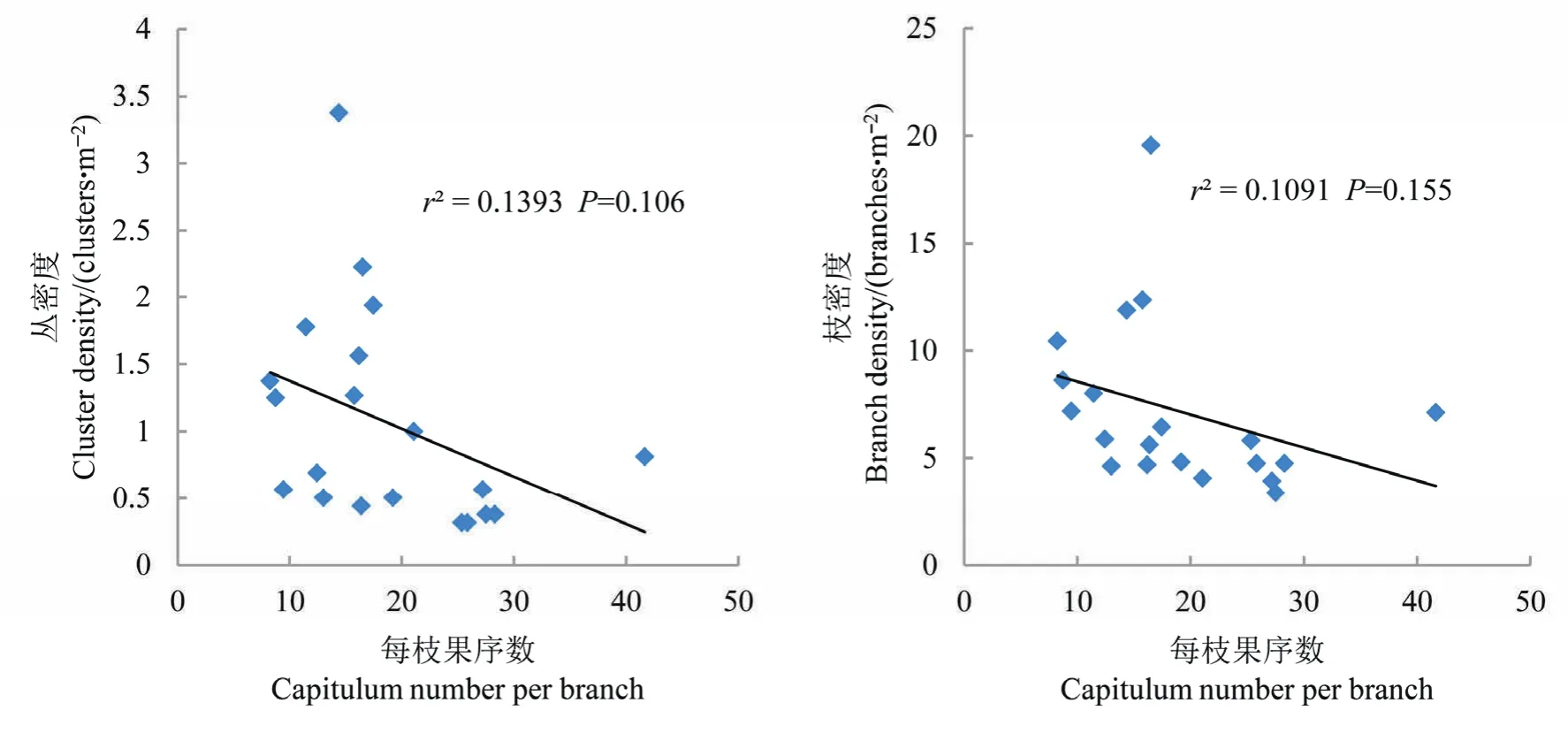
图4 肿柄菊群落每枝果序数与丛密度和枝密度关系Fig. 4 Relationships between capitulum number per branch, and cluster density and branch density in Tithonia diversifolia community

图5 18个地点的肿柄菊每果序平均结实和结籽量Fig. 5 The mean fruit and seed number per capitulum of Tithonia diversifolia from 18 sampling sites

表1 18个地点的肿柄菊每果序平均结实和结籽量的变化程度Table 1 Summary of mean numbers of fruits and seeds per capitulum of Tithonia diversifolia across all 18 sampling sites
2.4 主要环境因素对结籽率的影响
肿柄菊结籽率与年均温和 1月均温有显著的正相关关系(P<0.05),与7月均温有微弱的正相关关系(P<0.1),这表明肿柄菊的结籽率明显受温度的影响。肿柄菊结籽率与年降雨量、雨季降雨量和旱季降雨量都没有显著的相关关系(P>0.1),这表明肿柄菊的结籽率与分布区的降雨量关系不密切(图7)。肿柄菊结籽率与纬度不相关(P>0.1),与海拔有显著的负相关关系(P<0.05)(图7),这可能因为在采样区由于受地形影响,各种气候类型呈镶嵌型分布,温度随着纬度的升高并非有规律地连续性降低,与纬度因素相比温度更易受海拔变化的影响,所以结籽率与海拔高度呈现出显著的负相关关系。
3 讨论
3.1 肿柄菊结籽量对扩散的影响

图6 温度、降雨、纬度和海拔对肿柄菊每果序结实量的影响Fig. 6 Relationships between the mean number of fruits per capitulum of Tithonia diversifolia and temperature, rainfall, latitude and elevation

图7 肿柄菊结籽率与温度、降雨、纬度和海拔之间的关系Fig. 7 Relationships between seed set ratio of Tithonia diversifolia and temperature, rainfall, latitude and elevation
入侵植物的成功扩散要经过繁殖体的散布、幼苗建成、定居建群等多个环节,定居建群成功后再进一步扩散(刘建等,2010)。在扩散的整个过程中繁殖体的散布是首要环节,繁殖体的散布量是成功入侵的关键性因素,直接决定着最初的种群能否建立(Lockwood et al.,2005;Wonham et al.,2013)。陆生入侵植物的长距离扩散主要是依靠种子,种子的散布量也就决定了入侵植物侵占新领地的初始能力(Rejmanek et al.,1996;Davies et al.,2007)。
肿柄菊结籽量由果序量、每果序结实量和结籽率共同决定。肿柄菊群落的果序密度最少和最多可相差5倍多(图2),这种差异是由丛密度、枝密度、每枝果序数等群落特征共同决定。在云南的南亚热带和中亚热带地区肿柄菊分布最为普遍;在云南南部的热带地区由于受扩散条件的限制群落分布偏少,在北亚热带为近些年刚扩散而建立的群落,目前主要表现为零星点状分布(朱枫等,2018)。本文只在肿柄菊分布最为普遍的南亚热带和中亚热带区域进行群落调查,在热带、南亚热带和中亚热带肿柄菊的群落结构没有明显的差异(王四海等,2004),群落的果序密度在这些气候类型中应差别不大。在北亚热带小面积的零星群落必然会导致果序量总体较少。肿柄菊每果序结实量在不同地点间具有极显著差异(P<0.001),但是除了与温度有微弱的负相关关系外,与降雨、纬度、海拔等都没有显著的相关关系,这说明每果序结实量更多还是受局部生境条件的影响,在区域尺度范围并没有表现出明显的规律性,也就是说每果序结实量这一因素在区域范围并不会明显影响结籽量。
肿柄菊的结籽率最小为0.43%,最大为55.1%,两者相差120多倍,并与温度和海拔都表现出显著的相关关系(图7),这表明肿柄菊在跨气候类型扩散过程中结籽率明显在变化。结籽率不足 5%地点有彩云镇(CYZ)、甸中镇(DZZ)、弥勒北(MLB)和玉溪北(YXB),这些地点都是海拔和纬度较高区域。值得注意的是,最北端的鸡足山镇(JZS)虽然纬度和海拔都较高,但该地点属于金沙江干热河谷气候类型,所以结籽率较高。目前肿柄菊分布的最冷区域为北亚热带地区,在这一区域极低的结籽率和零星点状分布的群落只能产生极其有限的种籽量。尽管一些外来入侵植物可以通过表型可塑性和进化适应性来改变气候生态位以适应新的环境(Broennimann et al.,2007;Gallagher et al.,2010;Alexander et al.,2019),但是对于陆生入侵植物而言这种生态位适应性偏移是很有限度的(Petitpierre et al.,2012;Tingley et al.,2014),所以肿柄菊结籽率对不同气候的适应性不会明显改变。在较冷区域肿柄菊结籽量的大量减少会导致繁殖体压力极大降低,侵占新领地的能力减弱,限制了肿柄菊进一步向更高纬度和海拔扩散能力。
3.2 气候变化对肿柄菊分布范围的影响
气候变化影响着入侵进程的多个阶段,即使没有生态位适应性偏移,外来植物的入侵范围也会随着气候的变化而变化(Hellmann et al.,2008)。气温和降雨是影响植物分布的两个主要因素。外来入侵植物受全球气候变暖和降雨格局变化的综合影响,然而大多数的研究主要集中在气温变暖对植物入侵的影响(Bellard et al.,2013)。如果仅考虑全球气候变暖单一因素的情况下,全球入侵种的范围会进一步扩张(Hellmann et al.,2008;Walther et al.,2009;Hulme,2017)。与全球气候变暖相比,降雨格局和趋势有着更多的异质性,降雨变化对外来种的入侵影响就难以确定(Walther et al.,2009;Dukes et al.,2009;Dukes et al.,2011)。气候变化综合效应对入侵范围的影响就很难预测(Bradley et al.,2010),不同的入侵种响应气候变化的结果有较大差异(Lamsal et al.,2018)。在全球气候变化的情况下,具有多样化山地环境的云南很有可能面临着更温暖的气候和不确定的降雨格局(陈宗瑜,2001),受气温和降雨影响的外来物种未来的分布格局可能很难预测。肿柄菊在云南已有的分布区降雨量最少为金沙江干热河谷地区,年降雨量仅为578 mm,这也是云南降雨最少的区域;分布区降雨量最多地区在云南南部的西盟县,年降雨量多达2800 mm以上(陈宗瑜,2001;朱枫等,2018),可见肿柄菊生长对降雨量的适应性范围广。制约肿柄菊进一步扩散的结籽率因素主要受气温的影响,而不受降雨量的影响,考虑到肿柄菊在云南对降雨量的广泛适应性,所以降雨量的格局变化不会对肿柄菊分布带来大的影响。肿柄菊的结籽率与温度有着显著的正相关关系(图7),温度越低地区肿柄菊的结籽率越小,可以预测在气温变暖的情况下较冷地区分布的肿柄菊结籽率会提高,相应地肿柄菊侵占新领地的能力会增强,扩散范围会增大。在全球气候变化情况下,综合温度和降雨两个主要影响植物分布的因素考虑,尽管在云南山地环境下降雨格局不确定,肿柄菊的入侵范围也会随着气温的升高而扩大。
4 结论
肿柄菊的结籽量由果序量、每果序结实量和结籽率共同决定,果序密度和每果序结实量主要受局部生境条件的影响,在区域尺度范围没有表现出明显的规律性;结籽率在不同的地点间差异极其显著,并与温度和海拔分别表现出显著的正相关和负相关关系,在区域尺度范围结籽率明显影响着肿柄菊的结籽量。目前肿柄菊分布的最冷区域为北亚热带地区,在这一区域的极低结籽率和零星点状分布的群落只能产生极其有限的种子量,结籽量的大量减少会导致繁殖体压力极大降低,侵占新领地的能力减弱,限制了肿柄菊进一步向更高纬度和海拔扩散能力。考虑到肿柄菊生长对降雨量的广泛适应性,在全球气候变化情况下,尽管在云南山地环境下降雨格局不确定,肿柄菊的入侵范围也会随着气温的升高而扩大。
猜你喜欢
发明与创新·中学生(2023年9期)2023-09-21 09:19:02
林业勘查设计(2022年1期)2022-02-15 05:35:12
乡村科技(2021年17期)2021-10-20 08:50:56
安顺学院学报(2021年4期)2021-09-16 08:11:08
林业调查规划(2020年3期)2020-06-03 07:02:36
课外生活(小学1-3年级)(2020年2期)2020-03-09 03:24:26
阅读(快乐英语高年级)(2019年8期)2019-09-10 07:22:44
小火炬·智漫悦读(2018年12期)2018-03-13 09:24:52
莫愁(2017年36期)2017-12-25 05:52:36
中国绿色画报(2016年9期)2017-05-31 08:04:53
