
纳板河次生林不同林分群落中优势树种竞争强度探究
2019-08-16 07:11:04赖叶青张远荣胡明星张发山
新农民 2019年2期
赖叶青,张远荣,胡明星,张发山
(昆明驰云林业技术咨询有限公司,云南昆明 650000)
1 前言
一直以来国内在林业上的研究着重于对林分的非空间结构的研究,包括林分的树种组成、直径结构、密度、生长量、生物量及蓄积等指标的描述,它们提供了进行数量测定的基础,适用于森林资源统计与清查,方便地方政府部门或国家相应的部门对其进行管理、调控及监督。这些指标从数量上表示了林分的特征和生产力等一系列的信息,但这些数据由于缺乏空间分布信息,为林业工作者在对森林资源的经营和管理中增加了不少难题。因而涉及林分下单木之间空间关系的林分空间结构的相关特性也就越来越受到重视[1]。
在生态学研究中,优势树种在林分中的竞争强度是极其强,具有比较明显的地位,优势树种的变化表现为一个种群的变化。对一个种群的变化的研究也就变得较为重要,种群的研究主要包括了种群的分布格局、种群的年龄结构及种群的增长模型。种群空间分布格局分为均匀型空间分布、随机型空间分布和集群型空间分布3种类型。
林分中的种群特性决定了种群在林分空间中的分布结构。有不少的研究人员采用集块性指标、方差均方比及Cassie指标等空间结构参数来描述林分中的林木分布格局,但这些林分空间分布参数与距离无关,并且这些指标不能全面反映林分种群的空间分布格局。而以林木个体作为一个个点并定义点坐标为基础数据,能够较好的描述林分空间结构特征[2-5]。对于林分空间结构参数,惠刚盈等以在样地中选择参照树及其周围最近4株的相邻木作为一个基本的研究单元[6]。由此构建的单元:角尺度、混交度及大小比数[7],便可得到林分空间的竞争关系。惠刚盈及建立在平面中点的Ripley K-函数和双相关函数等分析,除此之外,Stanko Trifkovic[8-9]等人相关研究明确了角尺度在林分竞争上相比于Ripley K-函数和双相关函数更有效可行。
林分空间竞争指数有CI1、CI2、CI3、CI4、CI5、CI6。其中CI1计算出的是Hegyi竞争指数,其余模型计算出的是Hegyi竞争指数的变形。其中CI1、CI2和CI3分别是以胸径、树高和平均冠幅表示的简单竞争指数,而CI4、CI5和CI6则是以平方的形式强调竞争木与对象木的不对称性。各竞争指数的计算公式如下:
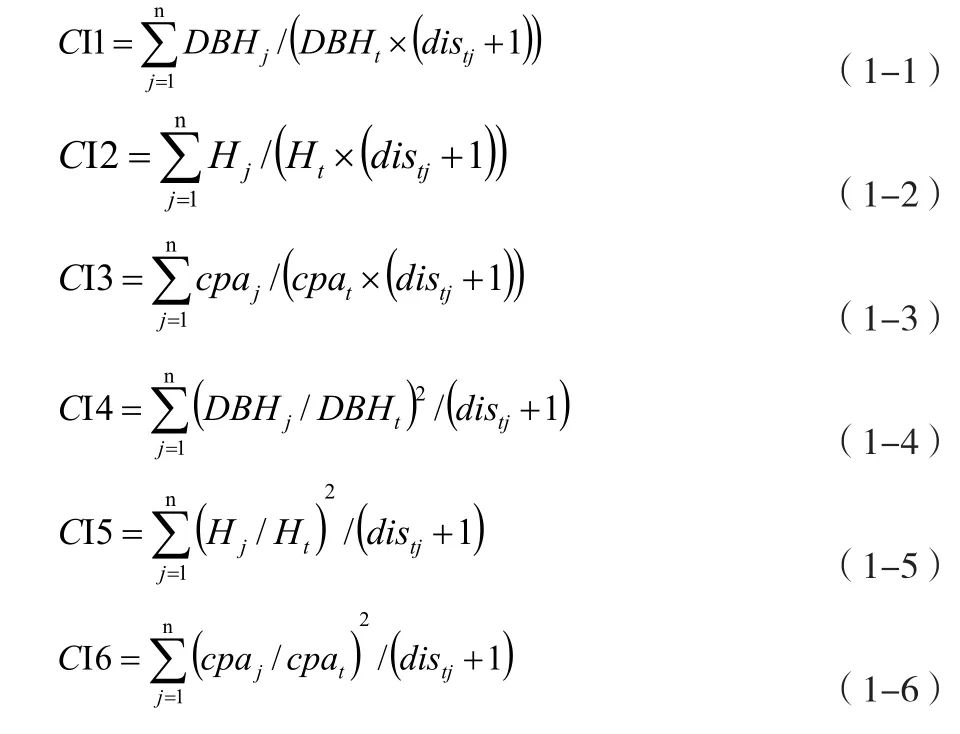
以上公式(1-1)至(1-6)中:DBH为林木胸径(cm),H为树高(m),cpa为平均冠幅(m),t表示第t株对象木,j表示第j棵竞争木;distj表示林分样地中的对象木与竞争木之间的直线距离;n表示第n棵株对象木的竞争木株数。
2 空间结构的研究方法
空间结构中的林分空间结构越来越得到广泛关注,林分空间结构定义为与林分空间位置有关的结构,是林分空间结构及其空间关系的阐述和表达,已成为林分结构研究的焦点。过去对结构的研究一般是针对群落结构中不同层次和级别、不同树种、不同年龄等内容,林分的空间结构主要包括林木的水平的直径分布情况、树种组成及竞争状况等。
2.1 确定空间结构单元
每一林分中的每株参照树及其相邻木间的空间关系构成了林分内最简单也是最基本的空间结构单元。在林分中,往往是这样的多个单元共同构成这片林分的空间竞争关系,其中空间结构单元的大小描述一般是在参照树周围选择3株或4株最近相邻木,但是即便这样,在实际操作中也比较难以量化的表示其中的相对位置关系,并且这最多也只能得到3个或4个方向上的空间关系,其它方位的空间信息是不完整的。判断方向用指北针等辅助工具,在野外调查时,受地理位置的影响,本身就有困难,但当超过4个方位务必相对比较困难一些,并且要严格的定位每一株与其他之间的相对位置,这在很大的程度上直接增加了测算的难度。
除此方法之外,还有一种比较常用的方法是以参照树为圆心,一定的距离作为竞争半径,但是由于立地质量、环境、海拔等等都会导致竞争半径发生较大的变化。不稳定因素比较突出,所以很多时候在采用此种方法时都要先进行竞争距离确定,从而也导致调查任务的工作量加大。
本研究则是以对象木为中心,周围一圈的临近木为竞争木进行研究林木的空间竞争关系,及依据arcgis中产生的泰森多边形对主要林分中空间结构参数进行研究。
2.2 林分空间结构参数的计算
林分的空间结构参数分析常用“结构组法”。但是这种空间结构在一些空间信息上还会表现的不够全面,因此本文的研究方法在基于此结构单元的基础上,把3株变为围绕对象树种一周的林木,其主要靠现地调查及在arcgis中的泰森多边形来确定竞争木。从而计算并确定各个主要林分样地中的角尺度、混交度和大小比数的具体数值。
3 研究方法
3.1 样地的选取
本研究中的样地选取基于纳板河自然保护区海拔、山势地形、季风气候、森林植被、坡度、坡向、坡位林木树种、林木径阶分布情况等多方面的考虑,及在充分的外业调查的基础上进行比较筛选而得出的样地,最终挑选出16块该片区域中具有比较典型性代表的样地,每一个样地的设置均为50m×70m,面积为3500m2。此次选在纳板河自然保护区内的六种典型的林分类型中,分别是:热带雨林群落、热带季雨林群落、季风常绿阔叶林群落、山地雨林群落、热性竹林群落和落叶阔叶林群落。依据不同海拔高度及不同植被类型等其他因素等共同选择建立16块样地,并逐年进行样地监测和调查。
3.2 标准地因子测定
(1)区划方格;样地的设置为50m×70m,对于热带森林体系,其中物种较多,面积较大容易侧漏、重测错测等,故在原有样地的基础上再对50m×70m,进行细分,把每一块大样地分成35个10m×10m,拉线区划标记。
(2)每木检尺;在大样地内细分样地,运用相邻格子法逐一布置小样方,并分别对每个小样方开展树种确定及每木检尺等工作,记录每个小样方中直径超过5cm的林木。并及时记录相应的乔木的种名、胸径及其他。由于此次样地的布设地为纳板河国家级自然保护区,改保护区位于热带、亚热带地区,保护区内大部分植物在高温多雨的环境中都是生长比较迅速,从而导致其乔木的树高、乔木的东西南北冠幅及乔木的枝下高等极难测量,故此次测量并不针对乔木的树高、乔木的东西南北冠幅及乔木的枝下高进行测量。只调查和记录各树种的密度、盖度以及直径以下的幼苗、幼树的株数及其样地周围的环境情况等等。
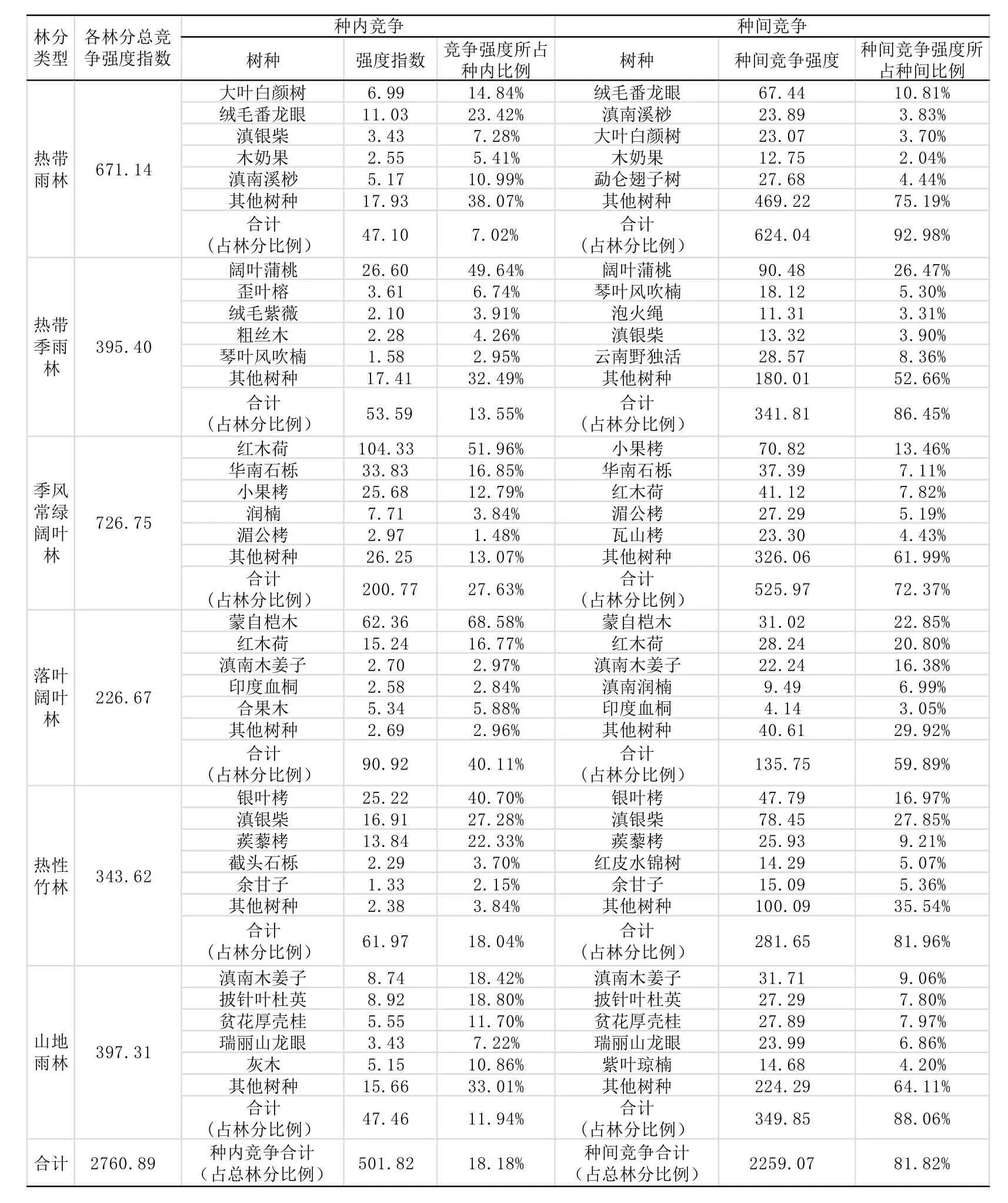
表1 不同森林植被类型的林分中种内及种间竞争
3.3 竞争强度计算
本文结合样地调查数据,依据六个林分空间竞争指数CI1、CI2、CI3、CI4、CI5及CI6,据惠刚盈[38]的研究中指出其中,CI1计算出的Hegyi竞争指数最能反映林分的空间关系,故此在本研究中只采用CI1,其余模型的计算在本研究中将不予计算。其CI1竞争指数的计算公式:
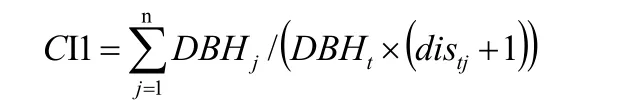
以上公式中:DBH为林木胸径(cm),t表示第t株对象木,j表示第j棵竞争木;distj表示林分样地中的对象木与竞争木之间的直线距离;n表示第n棵株对象木的竞争木株数。
4 研究结果
4.1 种内及种间竞争强度
林分中的林木在生长时,同种个体因有同样生态位需求,距离越近,数量越多,从而导致的竞争强度也就越大。同一片林分下的林木不仅仅影响其他林木,同时它也被周围林木所干扰影响。表4-1表示了不同森林植被类型的林分中种内及种间竞争较大的主要树种的竞争情况。
由上表可得,在林分类型上,此次所有林分样地的竞争指数为2760.89。季风常绿阔叶林的竞争强度指数为726.75;热带雨林的竞争强度指数为671.14;山地雨林的竞争强度指数为397.31;热带季雨林的竞争强度指数为395.40;热性竹林的竞争强度指数为343.62;落叶阔叶林的竞争强度指数为226.67。其中季风常绿阔叶林的总竞争指数表现的最高,依次为热带雨林、山地雨林、热带季雨林、热性竹林和落叶阔叶林。季风常绿阔叶林林分样地中386株;热带雨林林分样地中279株;山地雨林林分样地中246株;热带季雨林林分样地中302株;热性竹林林分样地中270株;落叶阔叶林林分样地中118株。各个林分强度竞争指数随着林分的密集程度的增加而增加,此外由于林分树种类型的不同、林分生境的不同也对林分竞争强度指数影响很大,热带季雨林中的阔叶蒲桃及热性竹林中的银叶栲均为各个林分的优势树种,但是该林分的这些优势树种生长表现均较差,特别是热性竹林中的银叶栲,平均高度只为6米。并且在热性竹林中优势树种银叶栲周围还伴生丛生竹林。丛生竹林不仅仅从空间上(阳光、二氧化碳、温度等),还有地下的养分(水份、无机盐等)都有资源竞争,但是本研究以乔木为主,并未涉及竹林在内,以致使得热性竹林林分整体的竞争指数相对较低。
从种内及种间看,同种个体因有相似的生态位需求,相对聚集分布,数量较多,竞争强度较大,不同种间则相对表现较弱。种内竞争指数为501.82。为总指数的18.18%;种间竞争指数为2259.07,为总竞争指数的81.82%。这是典型的热带、亚热带地区天然次生混交林的现状,由于林木种类过多,交错复杂,从而致使种间的竞争指数占主体,种内竞争仅仅占其中的较小部分。从种内及种间的竞争强度所占的比例来看,所有的种间竞争指数都比种内竞争指数大。落叶阔叶林林分样地的种内及种间竞争指数的变动明显小于其他各个林分种内及种间的竞争指数的变动。落叶阔叶林则种内种间较为相近,主要原因是因为落叶阔叶林的优势树种为蒙自桤木,这是由于该林分中蒙自桤木所占比例比较大,从而使得落叶阔叶林中种内竞争强度指数变大及种间的竞争强度指数变小,种内及种间的竞争强度指数相对较为接近。
从优势树种看,各个林分的优势树种都占据了该林分类型比较大的比例,无论是种内、种间或是整体而言都占据较大的比例。其中在种内竞争中,优势树种的竞争强度远大于其他组成树种,而在种间竞争中,热带雨林的优势树种绒毛番龙眼、热带季雨林的优势树种阔叶蒲桃、山地雨林的滇南木姜子及落叶阔叶林中的优势树种蒙自桤木的竞争强度也大于其他组成树种。季风常绿阔叶林中的红木荷和热性竹林中的银叶栲的竞争强度并非最大,就其主要的原因可能是红木荷和银叶栲在各自林分中的分布特点并不是均匀分布,其分布特点是以块状的形式散布在各个林分之中。
4.2 优势树种竞争强度比较
不同林分优势树种在很多方面上可以反映并代表该优势树种所处样地林分的一些生境的特点,能够直接简单的诠释该优势树种所处样地林分的相关属性,具有极其重要的代表作用。不仅仅如此,优势树种亦反映了在该林分下那种树种更适合生长繁殖,从而决定后期的林分中树种组成的变化。优势树种的变化直接反映出林分演替变化的结果,起到一定的指导作用。故了解各个林分中优势树种的变化对树种组成及林分演替具有重大的研究意义。
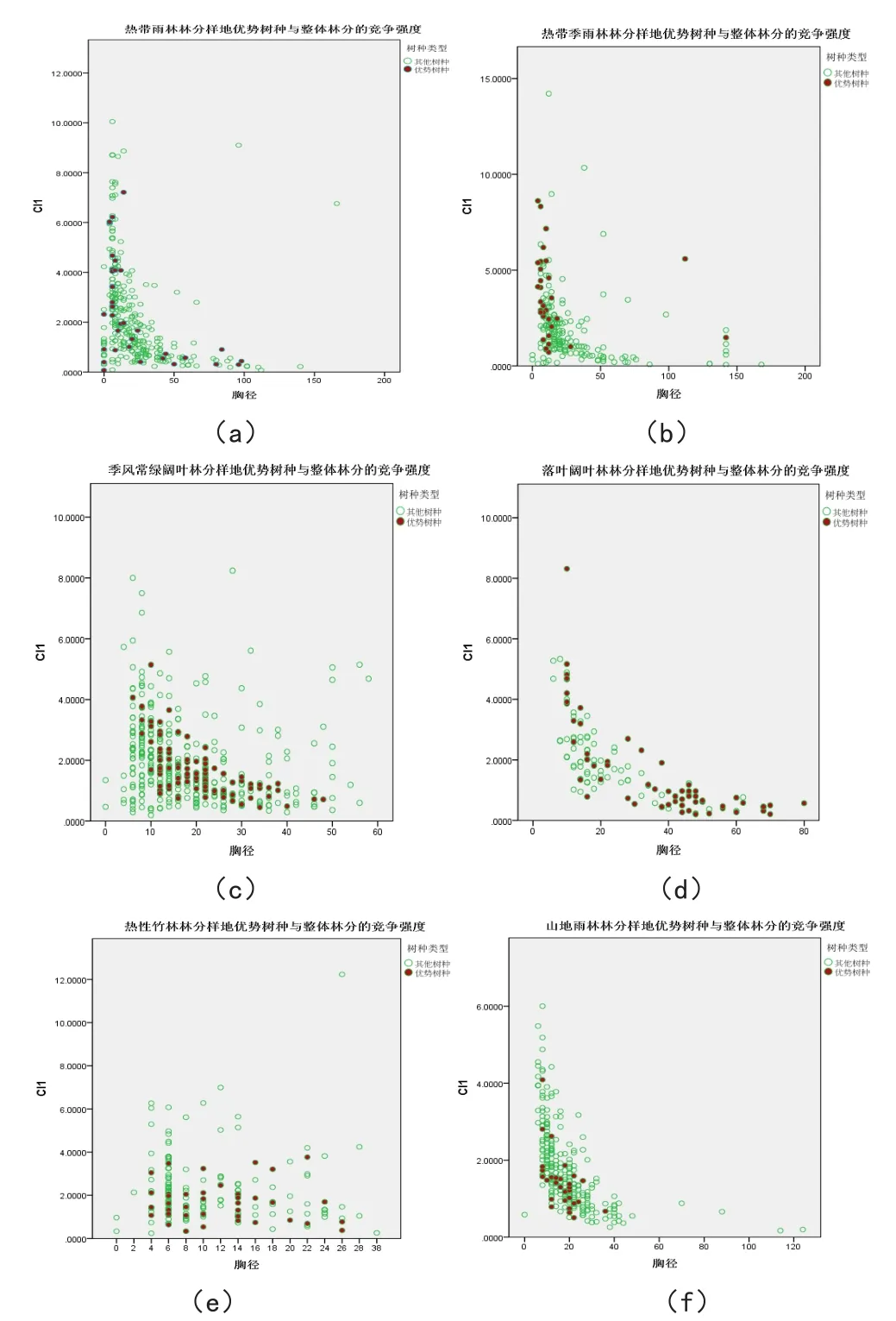
图1 优势树种在整体林分中的竞争
图1(a)热带雨林林分样地优势树种与整体林分的竞争强度;
图1(b)热带季雨林林分样地优势树种与整体林分的竞争强度;
图1(c)季风常绿阔叶林林分样地优势树种与整体林分的竞争强度;
图1(d)落叶阔叶林林分样地优势树种与整体林分的竞争强度;
图1(e)热性竹林林分样地优势树种与整体林分的竞争强度;
图1(f)山地雨林林分样地优势树种与整体林分的竞争强度。
由上图可以看出,优势树种和林分整体的竞争强度指数变化趋势是一样的。具有很强的代表性。其中图(a)热带雨林林分样地优势树种绒毛番龙眼与整体林分的竞争强度指数变化如出一辙,当林木胸径小于20cm时,随着径阶的增大竞争强度急剧下降。当对象木的胸径超过20cm时,下降趋势变得平缓,各径阶的竞争强度波动较小,且随着胸径的变大,各径阶的竞争强度趋于平稳不变;
图1(b)热带季雨林林分样地优势树种阔叶蒲桃与整体林分的竞争强度指数变化也较为接近,优势树种阔叶蒲桃主要集中在中小径阶部分,大径阶中并未有较多的阔叶蒲桃,这种情况说明阔叶蒲桃是后期近几年才新生的演替树种,按照这种生长趋势,在该片热带季雨林中,阔叶蒲桃的重要性将会显得越来越重要;
图1(c)季风常绿阔叶林林分样地优势树种红木荷与整体林分的竞争强度指数变化趋势很相近,林木各个胸径所对应的下降趋势整体而言都较为平缓,各径阶的竞争强度波动较小,且随着胸径的变大,各径阶的竞争强度趋于平稳不变,但是胸径从小到大的过程中,多会出现有零散的乔木的竞争强度高于整体水平,而优势树种红木荷的竞争强度指数变化相对于整体而言,变化不是很大,位于整体的最中间处,并处于一种比较集中的状态之中,相比之下表现的比较稳定、比较集中;
图1(d)落叶阔叶林林分样地优势树种蒙自桤木与整体林分的竞争强度指数变化趋势一样,当林木胸径不足20时,各径阶的竞争强度急剧下降,而当对象木的胸径超过20时,下降趋势变得平缓,各径阶的竞争强度波动较小,且随着胸径的变大,各径阶的竞争强度趋于平稳不变。但是优势树种蒙自桤木在中大径阶出表现得比较集中,而在小径阶处只有小部分的蒙自桤木,说明该林分中蒙自桤木处于一种衰亡的状态,之后可能会被其他物种所替代;
图1(e)热性竹林林分样地优势树种银叶栲与整体林分的竞争强度指数变化相近,林木各个胸径所对应的下降趋势整体而言都较为平缓,各径阶的竞争强度波动较小,且随着胸径的变大,各径阶的竞争强度趋于平稳不变,但是胸径从小到大的过程中,多会出现有零散的乔木的竞争强度高于整体水平,其原因主要是热性竹林中树木更新相对于其他林分较好,各个径阶的林木处于一种较为稳定的更新,从而各个径阶的竞争强度不会出现急剧下降的现象。优势树种蒙自桤木在整个林分中各个均匀相对比较均匀,处于一种较健康的发展模式;
图1(f)山地雨林林分样地优势树种滇南木姜子与整体林分的竞争强度。林木各个胸径所对应的下降趋势整体而言都较为平缓,各径阶的竞争强度波动较小,且随着胸径的变大,各径阶的竞争强度趋于平稳不变,但是胸径从小到大的过程中,多会出现有零散的乔木的竞争强度高于整体水平。但是优势树种滇南木姜子则主要集中在小径阶处,中大进阶处都没有滇南木姜子的存在。这反映出该优势树种滇南木姜子为近期刚刚在该山地雨林林分中生长繁殖的,并取得一定的优势占据该林分的主要位置。
猜你喜欢
阿来研究(2020年1期)2020-10-28 08:10:22
安徽农学通报(2020年17期)2020-10-09 10:46:23
防护林科技(2020年6期)2020-08-12 13:34:40
绿色科技(2019年6期)2019-04-12 05:38:42
绿色科技(2019年6期)2019-04-12 05:38:42
防护林科技(2016年8期)2016-08-29 01:28:17
新世纪水泥导报(2016年1期)2016-07-01 03:59:37
贵州科学(2016年2期)2016-06-13 00:58:58
华南农业大学学报(2016年3期)2016-06-01 09:27:59
中央社会主义学院学报(2016年2期)2016-05-04 04:18:28
