
低温寡照影响番茄幼苗根系有机酸代谢和养分吸收*
2019-08-14 13:30:32李煜姗杨再强1
中国农业气象 2019年8期
李煜姗,李 平,杨再强1,,汪 甫
低温寡照影响番茄幼苗根系有机酸代谢和养分吸收*
李煜姗1,2,李 平3,杨再强1,3,汪 甫2
(1.南京信息工程大学气象灾害预报预警与评估协同创新中心,南京 210044;2.江西省气象服务中心,南昌 330096; 3.江苏省农业气象重点实验室,南京 210044)
低温寡照气象灾害严重制约设施番茄产量和品质,本研究拟从番茄根系有机酸代谢和养分吸收的角度,探究低温寡照影响番茄生长的潜在机制。通过人工控制试验,设置不同低温(最高温/最低温:12℃/2℃、14℃/4℃、16℃/6℃、18℃/8℃)和弱光(200、400μmol·m−2·s−1)的交互处理,研究低温寡照处理2、4、6、8、10d后苗期番茄根系活力、氮磷钾含量、植株干重以及根系分泌低分子量有机酸(LMWOAs)的动态变化。结果表明:低温寡照显著抑制番茄根系活力和氮、磷、钾吸收,抑制根、茎叶干重增加,温度越低抑制作用越强;最低温(12℃/2℃)和最弱光(200μmol·m−2·s−1)处理下,番茄根系活力仅为对照的7.70%~22.1%,根系氮、磷、钾的净吸收量分别为对照的3.75%~18.1%、1.28%~27.1%和19.1%~35.5%,根系、茎叶干重分别为对照的23.4%~55.9%和42.6%~66.5%。低温寡照胁迫下番茄根系分泌低分子量有机酸的总量显著降低,土壤pH值升高,其中草酸的分泌量最大,下降幅度最明显。表明低温寡照对番茄生长的抑制作用可能与根系活力下降,草酸分泌减少和养分吸收降低有关,推测在低温寡照胁迫初期施加适量草酸或氮、磷、钾复合肥料或许可提高番茄的抗灾能力。
低温寡照;番茄;根系活力;土壤养分;有机酸
低温寡照是一种复合型气象灾害,一般发生在每年10月−翌年3月。番茄生长的最适温度和光合有效辐射(PAR)分别为15~25℃和550~650μmol·m−2·s−1[1],低温寡照势必会影响番茄植株生长。邹雨伽等[2]发现,昼/夜温12/2℃,PAR 200μmol·m−2·s−1低温寡照处理10d后花期番茄株高和茎粗的增长量较正常植株分别下降70.1%和94.9%。同时,番茄果实发育和品质形成也会受到影响,江梦圆等[3]发现花期番茄在昼/夜温12/2℃,PAR 200μmol·m−2·s−1低温寡照处理10d,成熟期果实体积较正常果实减小49.5%,果实内维生素C、可溶性固形物和糖酸等营养物质含量均显著下降[4]。低温寡照灾害严重制约了设施番茄产业的经济效益,但由于国内多数温室不具备控温、控光功能,目前仍难以通过调整温度和光强来缓解低温寡照胁迫[5]。因此,迫切需要了解低温寡照抑制番茄生长和果实形成的内在机制,进而提出合理的应急方案。
关于低温寡照对设施作物的致灾机制,国内外研究主要聚焦于低温寡照对叶片光合作用和氧化胁迫等方面的影响。低温寡照会抑制番茄叶片的光合作用,进而减少植株干物质累积[6]。低温寡照对番茄叶片光合作用的抑制主要体现在3个方面:(1)改变番茄叶片形态,Nieuwhof等[7]发现昼/夜温19℃/6℃,弱光照胁迫下番茄叶片的相对生长率和叶面积比率较正常植株显著下降,不利于叶片进行光合作用;(2)降低番茄叶片叶绿素含量,如高冠等[8]发现昼/夜温12℃/2℃,PAR 200μmol·m−2·s−1低温寡照胁迫下花期番茄叶片叶绿素a和叶绿素b均下降了36.0%左右;(3)弱化叶片对光能的利用效率,张淑杰等[9]研究指出低温寡照胁迫下,花期番茄叶片最大净光合速率、PSII最大光能转化效率、光化学淬灭系数均显著降低,导致光合作用受阻。另外,低温寡照还会刺激植物叶片产生活性氧,降低抗氧化酶活性,积累丙二醛,造成叶片的氧化损伤[10]。基于低温寡照抑制番茄叶片光合作用并造成氧化胁迫的致灾机制,Zhang等[11]提出通过转基因手段,将拟南芥中基因[C-repeat(CRT)and dehydration- responsive element(DRE)binding factor 1]引入番茄,加强低温寡照胁迫下叶片的光合作用和抗氧化能力,可有效促进番茄植株生长。但由于目前转基因蔬菜在国内仍未被公众接受,利用转基因手段加强番茄抵抗低温寡照能力的应对方案还难以实施。因此,仅仅关注番茄叶片对低温寡照的响应机制难以形成简单易行、可推广的应对措施,而番茄根系对低温寡照的响应目前还缺乏研究。
植物根系对外界环境变化较为敏感,在遭受胁迫后根系活力及根系代谢过程也会发生变化[12−14]。根系活力与植物吸收土壤水分和养分的能力有关,根系活力越弱,植物吸收水分和养分的能力越差。在根系代谢过程中,植物会向环境中分泌一些代谢物质,其中低分子量有机酸(LMWOAs)是植物根系分泌物的主要成分[15],它有利于难溶性养分的活化,促进植物养分吸收[16−17],还能提高根系活力,促进植物根系的生长发育[18]。逆境条件下,根系活力和低分子量有机酸代谢的变化均可能影响植物对土壤养分的吸收[19],这可能也是低温寡照抑制番茄生长的重要原因。
因此,本研究拟通过低温寡照胁迫试验了解番茄根系活力、低分子量有机酸代谢及氮磷钾等养分吸收的变化,通过分析上述指标与植株生长的关系,从番茄根系角度探究低温寡照对番茄的致灾机制,以期提出一种应对低温寡照胁迫的农艺方案,为缓解设施番茄低温寡照灾害提供创新思路。
1 材料与方法
1.1 试验设计
试验于2016年9月−2017年6月在南京信息工程大学农业气象试验站进行,以“金粉5号”为供试番茄品种。于Venlo可控温室进行育苗,待两叶一心时,选择生长健壮、长势一致的幼苗定植于PVC塑料花盆(口径25cm,高20cm)中,每盆种植2株。供试土壤为黄棕壤,基本理化性质为:pH值6.75,有机质含量11.6g·kg−1,有效氮69.4mg·kg−1,有效磷32.0mg·kg−1,有效钾180mg·kg−1。每盆装干土3kg,均匀拌入基肥:尿素0.78g、氯化钾11.1g和一水磷酸钙0.64g。
待番茄幼苗生长至第4片真叶展开、株高达15cm以上时将其移至人工气候箱(TPG-1260 Australian)内,进行为期10d的低温寡照处理。设置4个低温水平,即18℃/8℃(最高/最低温度)、16℃/6℃、14℃/4℃、12℃/2℃,模拟自然温度变化特征,当日5:00−14:00从最低温均匀上升至最高温,当日14:00−次日5:00均匀下降至最低温,变温过程由程序自动控制,具体参照朱丽云等[4]的设计;光合有效辐射(PAR)设置200、400μmol·m−2·s−1两个弱光水平;以28℃/18℃、PAR 1000μmol·m−2·s−1为对照,人工气候箱内空气湿度均设为75%,温度误差控制在±0.5℃范围内。低温寡照交互共8个处理,具体见表1,每个处理15盆。

表1 试验设计
1.2 样品采集和分析
分别于低温寡照胁迫后的第2、4、6、8、10天采集各处理根际土壤和番茄植株样品,各处理每个采样时期采集3盆样品,采样方法为破坏性采样,具体操作为:采样前2d不浇水或少量浇水,避免土壤过湿,将塑料盆用剪刀从侧边剪开,取出完整土体,将外围非根际土去除,利用抖根法将番茄根际微域范围内的土壤取出,视为根际土[20],分析根际土壤中低分子量有机酸含量;每盆取出2株完整番茄植株,1株用于测定根系和地上部干重、根冠比、根系氮磷钾浓度,另1株用于测定根系活力。
(1)根冠比
番茄植株按根系和茎叶分开,洗净后擦干。放入烘箱中105℃杀青30min后,75℃烘干至恒重,记录干重。根冠比的计算式为
根冠比=DW根系/DW茎叶(1)
式中,DW根系、DW茎叶分别为根系、茎叶干重(g)。
(2)根际土壤pH值及低分子量有机酸
根际土壤pH值(土水比为1:2.5)通过pH计(USA)测定[21]。
根际土壤中低分子量有机酸利用0.1M NaOH溶液和饱和NaCl溶液浸提,乙酸乙酯萃取,旋转蒸发后采用高效液相色谱法(LC-600,南京)进行测定[22],测定的低分子量有机酸种类包括草酸、甲酸、苹果酸、丙酸、琥珀酸和乙酸。
(3)根系活力及根系氮磷钾浓度
根系活力指根系的生长状况和代谢水平,可反映植物根系吸收水分和养分的能力,采用TTC法测定[23]。
根系氮磷钾的测定:将烘干后的根系用不锈钢磨样机磨成粉末,采用H2SO4−H2O2进行消解,消解液中全氮及全磷采用酶标仪法测定,全钾则采用火焰光度计法测定[21]。
低温寡照处理期内番茄根系氮、磷、钾净吸收量计算式为
Tnet=Tn−Tn−2(2)
式中,n为第2、4、6、8、10天,Tn为第n天时番茄根系氮、磷、钾总量,Tn−2为第n−2天时番茄根系氮、磷、钾总量,Tnet为第n−2天至第n天的氮、磷、钾净吸收量。
1.3 数据处理
数据分析及图表绘制采用SPSS16.0及Excel2010,液相色谱数据用伍豪色谱数据工作站进行处理。采用Duncan检验法(a=0.05)进行显著性分析。
2 结果与分析
2.1 低温寡照影响番茄根系活力
根系活力是反应根系生长和代谢水平状况的综合性指标,可直接影响植物地上部的生长和营养状况,根系活力高表示植物生长状况良好、代谢旺盛,反之则说明植物生长不良。低温寡照胁迫下番茄幼苗根系活力的变化如图1所示。由图可见,CK的根系活力在观测期内基本不变,保持在3000μg·g−1·h−1左右。而低温寡照处理下,观测期内根系活力显著低于CK(L2T4处理中第6、8天除外),且随胁迫时间呈先升高后降低趋势,在第6天达到峰值。在光照强度相同时,番茄根系活力与温度呈正比;在温度相同时,400μmol·m−2·s−1光照强度下根系活力在前8d高于200μmol·m−2·s−1处理,第10天二者差异变小。L1T1处理对根系活力的抑制最为明显,其根系活力在232~660μg·g−1·h−1范围内,为同期CK处理的7.7%~22.1%。说明低温寡照会抑制番茄根系活力,不利于根系代谢和养分吸收。
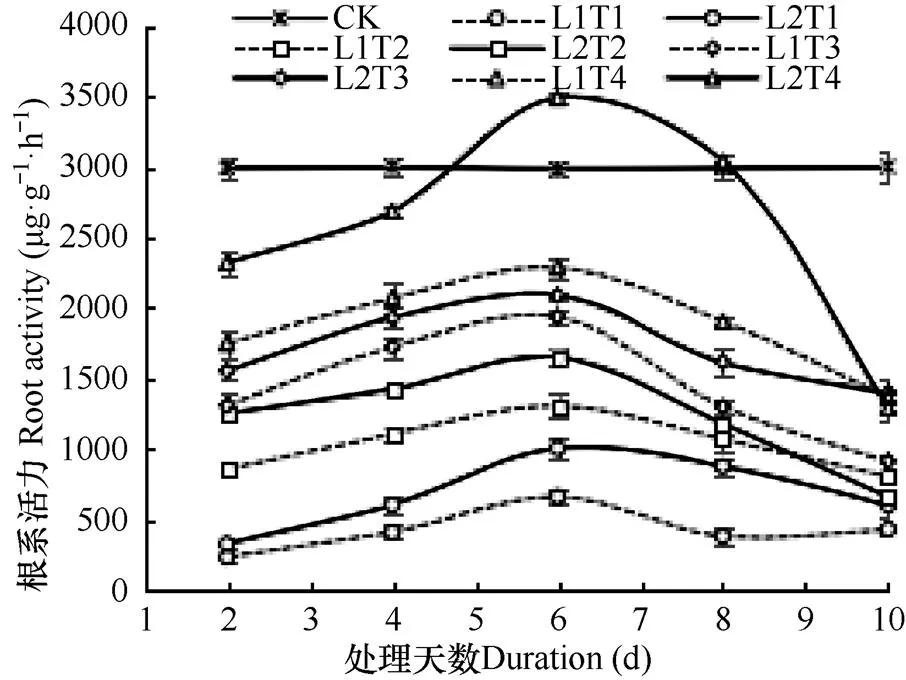
图1 各低温寡照胁迫处理10d期间番茄幼苗根系活力的动态变化
2.2 低温寡照影响番茄根系对氮磷钾的吸收
图2显示,观测期内对照(CK)处理单株番茄根系对氮的净吸收量保持在0.53~1.37mg水平,且随着生育进程先快速升高后轻微下降。低温寡照处理中,每次观测时根系对氮的净吸收量均明显低于CK,且随着温度和光照的降低,根系对氮的净吸收量也不断降低,在胁迫第6天时表现最明显;观测期内,L1T1和L2T1(温度最低处理)根系对氮的净吸收量仅为同期CK处理的3.75%~18.1%和12.1%~37.1%。可见,低温寡照明显抑制根系对土壤氮素的吸收。

图2 各低温寡照处理不同时段单株番茄幼苗根系对氮(a)、磷(b)、钾(c)的净吸收量
正常条件下(CK),观测期单株番茄根系对磷的净吸收量保持在0.025~0.120mg水平,前4d吸收量较低,后期较高。低温寡照处理时,根系对磷的净吸收量的变化规律与氮相似,L1T1和L2T1(温度最低处理)处理根系对磷的净吸收量仅为同期CK处理的1.28%~27.1%和19.1%~35.5%。可见,低温寡照明显抑制根系对土壤磷素的吸收。
正常条件下(CK),观测期单株番茄根系对钾的净吸收量保持在0.441~0.755mg水平,随着生育进程缓慢上升。低温寡照处理时,根系对钾的净吸收量的变化规律与氮、磷相似,L1T1和L2T1(温度最低处理)处理根系对钾的净吸收量仅为同期CK处理的2.15%~7.41%和7.02%~22.4%。可见,低温寡照明显抑制根系对土壤钾素的吸收,且抑制作用强于氮磷。
2.3 低温寡照影响番茄植株干物质累积
图3显示,观测期内对照(CK)处理单株番茄根系干重在0.070~0.245g范围内,且随着生育进程不断上升。低温寡照处理中,每次观测时干重均明显低于CK,且温度越低根系干重越低,光照越低根系干重也越低,随着生育进程,各处理与CK的差异不断增大;观测期内,L1T1和L2T1(温度最低处理)根系干重为同期CK处理的23.4%~55.9%和33.8%~73.3%。可见,低温寡照明显抑制根系生长。
正常条件下(CK),观测期单株番茄茎叶干重在0.756~1.686g范围内,且随着生育进程不断上升。低温寡照处理中,番茄茎叶干重的变化规律与根系相似,L1T1和L2T1(温度最低处理)处理茎叶干重为同期CK处理的42.6%~66.5%和63.1%~87.4%。可见,低温寡照明显抑制茎叶生长,但弱于对根系的抑制作用。
正常条件下(CK),观测期番茄根冠比在0.093~0.145范围内,随着生育进程缓慢上升。低温寡照处理中,每次观测时根冠比均明显低于CK,随着生育进程,各处理与CK的差异不断增大;各低温寡照处理间根冠比差异较小,但L1T1和L2T1(温度最低处理)处理根冠比仍最低,为同期CK处理的49.5%~84.1%和53.6%~85.8%。可见,低温寡照对番茄根冠比也有一定的抑制作用。
2.4 低温寡照影响番茄根系有机酸的分泌
由表2可见,CK处理根际土壤中,6种低分子量有机酸中草酸含量最高,占80.2%~88.3%,试验期间,草酸含量保持在34.6~58.9μg·g−1水平,且随生育进程草酸含量不断上升。低温寡照处理中,每次观测时草酸含量均明显低于CK,且温度越低土壤中的草酸含量越低,在观测期间(第6天)草酸含量达到峰值时该特点最明显,L1T1和L2T1(温度最低处理)处理土壤中的草酸含量仅为同期CK处理的6.31%~10.4%和7.38%~11.6%。可见,低温寡照明显抑制根系对草酸的分泌。

图3 各低温寡照胁迫处理10d期间单株番茄幼苗根系(a)、茎叶干重(b)和根冠比(c)的变化

表2 不同处理观测期间根际土壤低分子量有机酸浓度(μg·g−1)
注:数据均为平均值±标准差,nd表示未检测到。
Note: The data is mean±SD, nd indicates no detection.
正常情况下(CK),根区琥珀酸含量位列低分子量有机酸中第二,在3.79~4.61μg·g−1范围,随生育进程变化趋势不明显。在12℃/2℃以上时,低温寡照对土壤中琥珀酸含量的影响不大,与CK间差异不显著,当温度下降到12℃/2℃时土壤中观测不到琥珀酸,说明此时根系停止分泌琥珀酸或分泌量过少难以检测到。
正常情况下(CK),根区乙酸含量位列低分子量有机酸中第三,在1.23~3.37μg·g−1范围,随生育进程变化趋势不明显。当温度为18℃/8℃时,低温寡照处理显著提高乙酸含量,L1T4和L2T4处理土壤中乙酸含量为同期CK处理的2.69~4.70倍和1.78~13.3倍,而当温度降至18℃/8℃以下时土壤中观测不到乙酸,说明此时根系停止分泌乙酸或分泌量过少难以检测到。
正常情况下(CK),根区丙酸含量位列低分子量有机酸中第四,在0.44~0.94μg·g−1范围,随生育进程变化趋势不明显。低温寡照处理中,每次观测时丙酸含量均明显高于CK,且整体表现为温度越低土壤中丙酸含量越高,在观测期间(第6天)丙酸含量达到峰值时该特点最明显,L1T1和L2T1(温度最低处理)处理土壤中丙酸含量为同期CK处理的3.66~5.11倍和2.64~9.25倍。可见,低温寡照明显促进根系对丙酸的分泌。
正常情况下(CK),根区甲酸和苹果酸含量较低,分别在0.55~0.68μg·g−1和0.17~0.35μg·g−1范围,随生育进程变化趋势不明显。在12℃/2℃以上时,低温寡照处理对甲酸和苹果酸含量的影响不明显,当温度降至12℃/2℃时,在第4天观测期后,甲酸和苹果酸含量显著上升,说明此时低温寡照明显促进根系对甲酸和苹果酸的分泌。
由图4可见,对照(CK)处理根际土壤中总有机酸浓度为43.1~68.5μg·g−1,且随生育进程有机酸分泌量不断上升。低温寡照处理中,每次观测时总有机酸浓度均明显低于CK,温度越低总有机酸浓度越低,且观测期内呈先升高后降低趋势,在第6天达到峰值。在观测期内,L1T1和L2T1(温度最低处理)处理土壤总有机酸浓度为同期CK处理的12.0%~16.9%和14.9%~22.7%。可见,低温寡照会明显抑制根系对低分子量有机酸的分泌。

图4 各低温寡照胁迫处理10d期间根际土壤低分子量有机酸总浓度的动态变化
2.5 低温寡照影响根际微域范围内土壤酸碱度
根际微域范围内土壤pH值的变化规律与总有机酸的变化规律正好相反(图5),对照(CK)处理根际范围土壤pH值最低,为5.34~5.62,且随着生育期推进而不断降低。低温寡照处理中,每次观测时土壤pH值均高于CK,温度越低土壤pH值越高,随生育进程呈先降低后升高的趋势,在第6天达到最低值。观测期内,L1T1和L2T1(温度最低处理)处理土壤pH值比同期CK处理高13.2%~19.1%和12.3%~18.5%。可见,低温寡照会使番茄根际土壤pH值明显升高。

图5 各低温寡照胁迫处理10d期间根际土壤pH值的动态变化
3 结论与讨论
3.1 讨论
低温寡照不利于植物叶片的光合作用,显著影响植物生长[24−26]。除此之外,本研究发现低温寡照对番茄幼苗生长的抑制作用还与根系活力的降低有关。根系活力反映了根系的代谢能力,它的变化直接影响植株生长和抗逆性[27]。番茄根系活力对温度和光照的变化较敏感[13−14]。本研究发现低温寡照胁迫下,番茄根系活力受到明显抑制。但在胁迫初期根系活力有所增加,这可能是由于番茄幼苗为适应环境胁迫而作出的应激反应,但这种反应随着胁迫时间的延长会不断减弱[28]。根系活力会影响根系养分吸收,而低温寡照胁迫导致根系活力下降,可能是造成番茄植株生长受阻的主要原因之一。
低温寡照胁迫下番茄根系对土壤氮、磷、钾等养分吸收减弱也是造成番茄生长受阻的重要原因。本研究发现低温寡照显著抑制了根系对氮、磷、钾的吸收,这对番茄幼苗生长极为不利。除了抑制根系活力,低温寡照还会通过破坏根系韧皮部和木质部形态[29],限制根系对养分的吸收,进而影响番茄幼苗根系和茎叶的干物质累积[30−31]。Bingham等[32]研究发现,低温对番茄根系生长的抑制作用大于地上部,导致根冠比下降,本研究结果进一步证实了这一点。发育不良的根系不仅不利于养分和水分吸收,更会阻碍养分和水分向地上部运输[33],因此,低温寡照胁迫下,根冠比下降进一步抑制了番茄地上部生长。
外界环境(温度、光照等)的变化还将对植物的根系代谢过程造成影响,低分子量有机酸(LMWOAs)是植物根系通过代谢过程向土壤分泌的一类常见小分子有机物。研究表明,LMWOAs对降低土壤pH值,活化根际微域土壤矿质养分有重要作用[34]。根际土壤中LMWOAs主要来源于根系分泌、微生物分泌以及凋落物的降解[35−36],并会被微生物分解,处于动态变化中。但Gunina等[37]指出,在较短时间内(10d),微生物利用的有机酸仅占土壤总有机酸的0.1%~0.2%。而本研究中番茄幼苗只接受了10d的低温寡照处理,因此不考虑微生物对LMWOAs的分解作用。本研究发现草酸是番茄根系分泌物中主要的有机酸,这与Yang等[14,38]的研究结果一致。低温寡照胁迫显著降低了LMWOAs尤其是草酸的分泌。LMWOAs主要来源于线粒体中的三羧酸循环和乙醛酸循环的中间产物[39],易受植物呼吸作用和光合作用影响。在低温寡照条件下,番茄的光合速率降低,光合作用受到抑制,必然导致三羧酸循环和乙醛酸循环受阻[40],造成中间产物减少,进而使得LMWOAs分泌减少。LMWOAs分泌减少会导致根际土壤pH值升高,进而抑制土壤中氮、磷等养分的有效性[41−42],弱化植物对养分的吸收。研究发现,除了对根际矿质养分的活化作用,草酸本身对植物和微生物生长也有利[43−45]。因此,低温寡照处理下,草酸分泌量减少将对番茄幼苗生长产生诸多不利影响。另外,有研究表明丙酸会抑制灯笼椒的生长[46],而低温寡照胁迫下丙酸分泌增多,可能是造成番茄植株生长受抑制的另一个潜在原因。
3.2 结论
低温寡照胁迫显著抑制番茄根系活力,阻碍根系吸收氮磷钾,抑制根系分泌低分子量有机酸尤其是草酸,并提高了根际土壤pH值,这可能是低温寡照抑制番茄生长的重要原因。基于本研究结果推测,在低温寡照发生初期向土壤施加适量草酸和氮、磷、钾复合肥可能会缓解低温寡照对番茄的不利影响,但仍需要进一步通过试验加以验证。
[1] 高援献.番茄茄子栽培技术[M].北京:中国盲文出版社,1999: 10-11.
Gao Y X.The cultivation techniques of tomato and eggplant [M].Beijing:China Braille Press,1999:10-11.(in Chinese)
[2] 邹雨伽,高冠,杨再强,等.低温寡照对番茄花期植株生长及干物质分配的影响[J].江苏农业科学,2016,44(12):178-184.
Zou Y J,Gao G,Yang Z Q,et al.Effect of low temperature and weak light at flowering stage on plant growth and dry weight distribution[J].Jiangsu Agricultural Science,2016,44(12): 178-184.(in Chinese)
[3] 江梦圆,杨再强,王明田,等.花期低温寡照对番茄植株生长及果实发育的影响[J].江苏农业科学,2018,46(7):125-131.
Jiang M Y,Yang Z Q,Wang M T,et al.Effect of low temperature and weak light at flowering stage on plant growth and fruit development[J].Jiangsu Agricultural Science,2018, 46(7):125-131.(in Chinese)
[4] 朱丽云,杨再强,李军,等.花期低温寡照对番茄开花坐果特性及果实品质的影响[J].中国农业气象,2017,38(7):456-465.
Zhu L Y,Yang Z Q,Li J,et al.Effect of low temperature and weak light at flowering stage on flower-fruit characteristics of tomato[J].Chinese Journal of Agrometeorology, 2017,38(7): 456-465.(in Chinese)
[5] 魏瑞江.日光温室低温寡照灾害指标[J].气象科技,2003, 31(1):50-53.
Wei R J.The disaster grades of low temperature and spare sunlight in greenhouse[J].Mereological Science and Technology, 2003,31(1):50-53.(in Chinese)
[6] 于红,黎贞发,罗新兰,等.低温寡照对日光温室番茄幼苗生长的影响[J].北方园艺,2011,(24):56-60.
Yu H,Li Z F,Luo X L,et al.Effect of low temperature and less sunlight on the growth of tomato seedling in solar greenhouse [J].Northern Horticulture,2011,(24):56-60.(in Chinese)
[7] Nieuwhof M,Garretsen F,van Oeveren J C.Growth analysis of tomato genotypes grown under low energy conditions[J]. Netherlands Journal of Agricultural Science,1991,39(3): 191-196.
[8] 高冠,邹雨伽,杨再强,等.低温寡照胁迫对设施番茄花期叶片衰老特性的影响[J].北方园艺,2016,(5):49-55.
Gao G,Zou Y J,Yang Z Q,et al.Effect of low temperature and low irradiation stress on senescence of greenhouse tomato during flowering period[J].Netherlands Journal of Agricultural Science,2016,(5):49-55.(in Chinese)
[9] 张淑杰,杨再强,陈艳秋,等.低温、弱光、高湿胁迫对日光温室番茄花期生理生化指标的影响[J].生态学杂志,2014,33(11): 2995-3001.
Zhang S J,Yang Z Q,Chen Y Q,et al. Effects of low temperature,weak light and high humidity stresses on the physiological and biochemical indicators of greenhouse tomato during flowering period[J].Chinese Journal of Ecology, 2014,33(11):2995-3001.(in Chinese)
[10] Asada K.The water-water cycle in chloroplasts:scavenging of active oxygens and dissipation of excess photons [J].Annual Review of Plant Biology,1999,50(50):601-639.
[11] Zhang Y J,Yang J S,Guo S J,et al.Over-expression of the Arabidopsis CBF1 gene improves resistance of tomato leaves to low temperature under low irradiance[J].Plant Biology,2010,13(2011):362-367.
[12] Dresbøll D B,Thorup-Kristensen K.Spatial variation in root system activity of tomato (L.) in response to short and long-term waterlogging as determined by 15 N uptake[J].Plant & Soil,2012,357(1-2):161-172.
[13] Adil M,Abbasi B H,Khan T.Interactive effects of melatonin and light on growth parameters and biochemical markers in adventitious roots ofL[J].Plant Cell Tissue & Organ Culture,2015,123(2):405-412.
[14] Yang Z Q,Li Y S,Li P,et al.Effect of difference between day and night temperature on tomato(Mill.)root activity and low molecular weight organic acid secretion[J].Soil Science & Plant Nutrition,2016,62(5-6): 423-431.
[15] Wu Y Y,Xing D K.Effect of bicarbonate treatment on photosynthetic assimilation of inorganic carbon in two plant species of Moraceae[J].Photosynthetica,2012,50(4):587-594.
[16] 黄建凤,吴昊.植物根系分泌的有机酸及其作用[J].现代农业科技,2008,(20):323-324.
Huang J F,Wu H.Organic acids secreted by plant roots and their functions[J].Modern Agricultural Technology,2008,(20): 323-324.(in Chinese)
[17] Huang Q Y,Zhao Z H,Chen W L.Effects of several low-molecular weight organic acids and phosphate on the adsorption of acid phosphatase by soil colloids and minerals[J].Chemosphere,2003,52(3):571-579.
[18] 王文波.凋落物源有机酸对暗棕壤磷的活化及对水曲柳生理和生长的影响[D].哈尔滨:东北林业大学,2010:79-81.
Wang W B.Organic acids from forest litters and the effects on P release from dark brown forest soil and physiological characteristics and growth of.seedings [D].Haerbin:Northeast Forestry University,2010:79-81.(in Chinese)
[19] Yan Q Y,Duan Z Q,Mao J D,et al.Effects of root-zone temperature and N,P,and K supplies on nutrient uptake of cucumber(L.) seedlings in hydroponics [J].Soil Science and Plant Nutrition,2012,58:707-717.
[20] Chen M C,Wang M K,Chiu C Y,et al.Determination of low molecular weight dicarboxylic acids and organic functional groups in rhizosphere and bulk soils of Tsuga and Yushania in a temperate rain forest[J].Plant and Soil,2001,231(1): 37-44.
[21] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000:12-14,308-315.
Lu R K.Analytical methods of agricultural chemistry in soil[M].Beijing:China Agricultural Science and Technology Press,Beijing,2000:12-14,308-315.(in Chinese)
[22] 李煜姗,杨再强,李平,等.高效液相色谱法测定设施番茄土壤低分子量有机酸的色谱条件研究[J].土壤通报,2016, 47(1):73-78.
Li Y S,Yang Z Q,Li P,et al.Study on chromatographic condition in determination of low molecular weight organic acids in tomato-planted soil under greenhouse with HPLC[J].Chinese Journal of Soil Science,2016,47(1):73-78. (in Chinese)
[23] Knievel D P.Procedure for estimating ratio of live to dead root dry-matter in root core samples[J].Crop Science,1973, 13(1):124-126.
[24] Dongsansuk A,Lütz C,Neuner G.Effects of temperature and irradiance on quantum yield of PSII photochemistry and xanthophyll cycle in a tropical and a temperate species [J].Photosynthetica,2013,51(1):13-21.
[25] Fujimoto M,Nishihara G N,Terada R.The effect of irradiance and temperature on the photosynthesis of two agarophytesand(Gelidiales) from Kagoshima,Japan[J].Fisheries Science,2014,80(4):695-703.
[26] 朱雨晴,薛晓萍.遮阴及复光对花果期番茄叶片光合特性的影响[J].中国农业气象,2019,40(2):126-134.
Zhu Y Q,Xue X P.Effect of shading and light restoration on photosynthetic characteristics of tomato leaves during flowering and fruit period[J].Chinese Journal of Agrometeorology, 2019,40(2):126-134.(in Chinese)
[27] Ruf M,Brunner I.Vitality of tree fine roots:reevaluation of the tetrazolium test[J].Tree Physiology,2003,23(4):257-263.
[28] Ellingson K A,Rajapakse N C,Riley M B.Phytochemical profile changes of tomatoes in response to altered light environments[J].Hortscience,2004,39(4):759.
[29] Ou L J,Wei G,Zhang Z Q,et al.Effects of low temperature and low irradiance on the physiological characteristics and related gene expression of different pepper species[J]. Photosynthetica,2015,53(1):85-94.
[30] Khuankaew T,Tanabata S,Yamamoto M,et al.Temperature affects N and C assimilation and translocation inGagnep[J].Journal of Horticultural Science & Biotechnology,2014,89(3):287-292.
[31] Ylivainio K,Peltovuori T.Phosphorus acquisition by barley (L.) at suboptimal soil temperature [J].Agricultural Food Science,2012,21(4): 453-461.
[32] Bingham I J,Cumbus I P.Influence of root temperature on the potassium requirements of young tomato plants[J].Plant & Soil,1991,133(2):227-237.
[33] 吴立群,蔡志欢,张桂莲,等.低温对不同耐冷性水稻品种秧苗生理特性及根尖解剖结构的影响[J].中国农业气象, 2018,39(12):805-813.
Wu L Q,Cai Z H,Zhang G L,et al.Effects of low temperature on physiological characteristics of rice seedlings with different cold tolerance an anatomical structure of root tip[J].Chinese Journal of Agrometeorology,2018,39(12): 805-813.(in Chinese)
[34] Vítková M,Komárek M,Tejnecký V,et al.Interactions of nano-oxides with low-molecular-weight organic acids in a contaminated soil[J].Journal Hazardous Materials,2015, 293:7-14.
[35] Bahn M,Schmitt M,Siegwolf R,et al.Does photosynthesis affect grassland soil-respired CO2and its carbon isotope composition on a diurnal timescale[J].New Phytologist,2008, 182(2):451-460.
[36] Mencuccini M,Hölttä T.The significance of phloem transport for the speed with which canopy photosynthesis and belowground respiration are linked[J].New Phytologist,2010, 185(1):189-203.
[37] Gunina A,Dippold M A,Glaser B,et al.Fate of low molecular weight organic substances in an arable soil:from microbial uptake to utilisation and stabilization[J].Soil Biology & Biochemistry,2014,77:304-313.
[38] Ström L,Olsson T,Tyler G.Differences between calcifuge and acidifuge plants in root exudation of low-molecular organic acids[J].Plant and Soil,1994,167(2):239-245.
[39] López-Bucio J,Nieto-Jacobo M F,Ramı́Rez-Rodrı́Guez V,et al.Organic acid metabolism in plants: from adaptive physiology to transgenic varieties for cultivation in extreme soils[J].Plant Science,2000,160(1):1-13.
[40] Mangiapia M,Scott K.From CO2to cell:energetic expense of creating biomass using the Calvin-Benson-Bassham and reductive citric acid cycles based on genome data[J].Fems Microbiology Letters,2016,363(7):fnw054.
[41] 张星,刘杏认,林国林,等.生物炭和秸秆对华北农田表层土壤矿质氮和pH值的影响[J].中国农业气象,2016,37(2): 131-142.
Zhang X,Liu X R,Lin G L,et al. Effects of biochar and straw return on mineral nitrogen and pH of the surface soil in farmland of the north China plain[J].Chinese Journal of Agrometeorology,2016,37(2):131-142.(in Chinese)
[42] 张皓禹,黄志华,王娟,等.不同酸化剂对石灰性土壤pH值、磷有效性的影响[J].中国土壤与肥料,2019,(1):145-150.
Zhang H Y,Huang Z H,Wang J,et al. Effects of different acidifiers on pH and phosphorus availability in calcareous soil[J].Chinese Journal of Soil and Fertilizer,2019,(1): 145-150.(in Chinese)
[43] Pan F J,Liang Y M,Zhang W,et al.Enhanced nitrogen availability in karst ecosystems by oxalic acid release in the rhizosphere[J].Frontiers in Plant Science,2016,7:687.
[44] Wang Y Z,Whalen J K,Chen X,et al.Mechanisms for altering phosphorus sorption characteristics induced by low- molecular-weight organic acids[J].Canadian Journal of Soil Science,2016,96(3):289-298.
[45] Wani M A.Oxalic acid effect on potassium release from typical rice soils of Kashmir[J].Communications in Soil Science & Plant Analysis,2012,43(8):1136-1148.
[46] 徐慧敏.低分子有机物质对辣椒生长发育过程的效应研究[D].杨凌:西北农林科技大学,2009:20-29.
Xu H M.Studies on the effects of low molecular organic matter on the growth and development process of pepper[D].Yangling:Northwest A&F University,2009:20-29. (in Chinese)
Low Temperature and Low Irradiation Affected the Metabolism of Low-Molecular- Weight Organic Acids and Nutrients Uptake in Tomato Seedling Root
LI Yu-shan1,2, LI Ping3, YANG Zai-qiang1,3, WANG Fu2
(1.Collaborative Innovation Center on Forecast and Evaluation of Meteorological Disasters, Nanjing 210044, China; 2.Meteorological Service Center of Jiangxi, Nanchang 330096; 3.Jiangsu Key Laboratory of Agricultural Meteorology, Nanjing 210044)
Combined low temperature and low irradiation(LTLI) conditions seriously affect the yield and quality of tomato, the present study aimed to clarify the underlying mechanisms through investigating the changes of organic acids metabolism and nutrients uptake in tomato root. A pot experiment in artificially controlled environments was conducted to study the dynamic changes of root activity, nitrogen(N), phosphorus(P), and potassium(K) uptake by the root, dry weights of the root and shoot, and the secretion of low-molecular-weight organic acids(LMWOAs) by the root, under different LTLI conditions (maximum/minimum temperature: 12/2℃, 14/4℃, 16/6℃, 18/8℃; weak irradiation: 200, 400μmol·m−2·s−1). Results showed that the LTLI strongly inhibited the root activity, net absorption of N, P and K, and dry weights of the root and shoot, and the effects were enhanced with the decrease of temperature. Under the condition of 12/2℃ and 200μmol·m−2·s−1, the root activities were 7.70%−22.1% of the control, net absorption of N, P and K in the root were 3.75%−18.1%, 1.28%−27.1%, and 19.1%−35.5% of the control, respectively, dry weights of the root and shoot were 23.4%−55.9% and 42.6%−66.5% of the control, respectively. The LTLI also inhibited the secretion of LMWOAs and increased the soil pH, oxalic acid was the main acid which decreased most obviously. This study indicates that the inhibition of the LTLI on the growth of tomato seedling was related to the decrease of root activity, secretion of oxalic acid, and the nutrients uptake. Therefore, suitable application of oxalic acid or complex fertilizer (including N, P, K) at the initial stage of the LTLI occurrence, may enhance the resistance ability of tomato seedling to the LTLI.
Low temperature low irradiation; Tomato; Root activity; Soil nutrient; Organic acid
10.3969/j.issn.1000-6362.2019.08.004
李煜姗,李平,杨再强,等.低温寡照影响番茄幼苗根系有机酸代谢和养分吸收[J].中国农业气象,2019,40(8):512-522
2019−01−09
。E-mail:yzq@nuist.edu.cn
国家自然科学基金(41475107;41775104);“十二五”国家支撑计划(2014BAD10B07)
李煜姗(1993−),女,瑶族,硕士,助理工程师,主要从事气象灾害与气象服务研究。E-mail:liyu shan529@163.com
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02 01:06:26
落叶果树(2021年6期)2021-02-12 01:28:54
文苑(2020年12期)2020-04-13 00:54:10
家庭医学(下半月)(2019年9期)2019-10-12 08:03:56
海峡姐妹(2019年1期)2019-03-23 02:42:46
中成药(2018年5期)2018-06-06 03:12:15
天然产物研究与开发(2018年3期)2018-05-07 06:38:35
中国蔬菜(2016年8期)2017-01-15 14:23:38
中国茶叶加工(2015年3期)2015-02-27 07:55:30
河南城建学院学报(2014年4期)2014-02-27 07:08:52
