
不同基质条件下食品防腐剂对冷藏河鲀鱼片腐败菌的抑菌差异
2019-07-30 02:56:12马永生李智博杨朋霏丁吉佟赵前程
中国食品学报 2019年6期
李 萌 马永生 李智博 杨朋霏 丁吉佟 赵前程
(大连海洋大学食品科学与工程学院 辽宁大连116003)
水产品因富含蛋白质等营养物质和高水分,在加工、 贮藏过程中品质会逐渐降低并易发生腐败变质现象。 引起水产品腐败的主要原因是外源性腐败菌的生长繁殖, 采用食品防腐剂抑制腐败菌是水产品保鲜的有效手段之一[1]。食品防腐剂按照化学性质分为酸性防腐剂如山梨酸及其钾盐,酯型防腐剂如尼泊金酯类, 无机盐防腐剂如亚硫酸盐,以及生物防腐剂如乳酸链球菌素(Nisin)等。由于无机盐防腐剂容易分解后残留二氧化硫,所以目前我国的食品法规中仅将其列入特殊防腐剂且不能在肉制品中使用[2]。 国内外的研究表明,不同化学性质的食品防腐剂如山梨酸钾、 尼泊金乙酯和Nisin,均能够有效抑制水产品腐败菌的生长繁殖,延长水产品的货架期[3-5]。
不同细菌对防腐剂的耐受性具有显著性差异[6]。 在实际的保鲜应用中,常用多种防腐剂复配进行水产品保鲜处理。有研究表明,食品防腐剂在不同的食品基质条件下对同一种腐败菌也会呈现出抑菌活性的差异[7]。测定不同基质条件下防腐剂对腐败菌的抑菌效果对筛选复配防腐剂尤为重要。
养殖红鳍东方鲀(Tackifugu Rubripes)味道鲜美,营养价值较高,深受消费者的喜爱。 随着国内外市场需求增长, 对河鲀鱼的鲜度和食用品质也提出更高的要求。目前,有利用电解水或(和)壳聚糖涂膜技术开展冷藏河鲀鱼片保鲜工作的研究报道[8-10]。 随着河鲀鱼市场的开放,产品种类不断增加, 然而针对河鲀鱼片的复配保鲜剂尚未得到深入的研究, 食品防腐剂和基质条件对冷藏河鲀鱼片腐败菌的抑菌效果的研究鲜见报道。 本文以养殖红鳍东方鲀鱼片中分离获得的腐败菌为对象,分析腐败菌种类和致腐能力, 并比较不同基质条件下山梨酸钾、尼泊金乙酯和Nisin 3 种不同类型的防腐剂对腐败菌的抑菌效果, 以期为冷藏红鳍东方鲀鱼片的防腐保鲜应用提供理论数据。
1 材料与方法
1.1 材料与培养基
河鲀鱼片,由大连天正实业有限公司提供,将鲜活养殖红鳍东方鲀按GB/T 27624-2011 《养殖红鳍东方鲀鲜、冻品加工操作规范》的规定加工处理成鱼片,放于食品级托盘后,进行封膜包装,低温保温箱运送至大连海洋大学实验室低温贮藏。
营养肉汤、 营养琼脂均购于北京陆桥技术有限责任公司。液体、固体培养基均由成品制剂与蒸馏水按规定比例配制,121 ℃,15 min 高压灭菌。
1.2 试剂与仪器
即用PCR 扩增试剂盒(Taq),生工生物工程(上海)股份有限公司;山梨酸钾,宁波王龙科技股份有限公司;尼泊金乙酯,浙江圣效化学品有限公司;乳酸链球菌素,河南豫兴生物科技有限公司。
多功能酶标仪,美国伯腾仪器有限公司;洁净工作台,苏州安泰空气技术有限公司;压力蒸汽灭菌锅,上海三申医疗器械有限公司;电热恒温鼓风干燥箱,上海跃进医疗器械厂;无菌均质器,宁波新芝生物科技有限公司;PCR 扩增仪, 英国Techne 公司;凝胶成像仪,美国uvp 公司。
1.3 方法
1.3.1 冷藏河鲀鱼片腐败菌的分离和纯化 托盘包装的河鲀鱼片在0~4 ℃冰箱保藏至第13 天时色泽发黄,肉质无弹性发黏,纹理模糊,有明显的酸臭味,感官判断为腐败末期,此时无菌取样10 g鱼片至无菌均质袋内,加入90 mL 无菌生理盐水,均质后,用无菌生理盐水进行10 倍稀释,选取合适的3 个稀释梯度,取100 μL 稀释液在营养琼脂平板上涂布。 平板翻转倒置于恒温培养箱内,30℃培养72 h。 从平板中挑选菌落形态差异较大的细菌,重复至少3 次平板划线,得到纯化后的腐败菌单菌落。
1.3.2 冷藏河鲀鱼片腐败菌分子生物学鉴定采用沸水浴法提取细菌DNA, 构建PCR 扩增体系,利用如下PCR 扩增条件进行扩增:94 ℃变性5 min,进行35 个循环(94 ℃30 s,55 ℃30 s,72 ℃90 s),72 ℃延伸10 min。取9 μL PCR 扩增产物进行1%琼脂糖凝胶电泳, 若PCR 扩增条带单一明亮,则将PCR 扩增产物送至生工生物工程(上海)股份有限公司进行测序工作;16s rDNA 序列利用NCBI 中的Blast 进行序列比对, 获知相似性99%以上的菌株种属后, 利用NCBI 和RDP 数据库下载相似菌株或模式菌株序列, 使用MEGA 6 软件, 用邻接法(Neighbor-Joining 法)Kimura twoparmeter 计算方法构建菌株的系统发育树。
1.3.3 冷藏河鲀鱼片腐败菌致腐能力分析 将冷藏河鲀鱼片用无菌生理盐水冲洗, 无菌条件下沥干,浸泡至含105CFU/mL 腐败菌的生理盐水中10 s,空白组浸泡无菌生理盐水,取出沥干后,放于无菌袋中在冷藏条件下保存, 每隔1 d 测定样品的挥发性盐基氮(TVBN)和三甲胺含量(TMA)。
1.3.4 不同基质对腐败菌生长量的影响
1)液体培养基和液体鱼肉汁基质的制备:采用营养肉汤培养基,按比例添加至蒸馏水,加热溶解后,调整pH 至6.4,121 ℃,15 min 高压灭菌;取河鲀鱼片100 g 加入到400 mL 蒸馏水中,组织捣碎机捣碎,煮沸、过滤,121 ℃高压灭菌15 min,即获得无菌鱼肉汁(pH 为6.4);
2)测定不同基质对腐败菌生长量的影响:将900 μL 液体培养基或液体鱼肉汁,100 μL 106CFU/mL 的菌液加入到无菌48 孔板中,放置于25℃培养箱内,测定培养0 和24 h 时595 nm 波长下的吸光度值,计算各组的ΔOD595nm值(ΔOD595nm=培养后OD595nm-培养前OD595nm), 评估培养基质对腐败菌生长的影响。
1.3.5 不同基质下防腐剂对腐败菌的差异性分析参考潘晓倩等[11]的方法,略作修改。 将800 μL液体培养基或液体鱼肉汁,100 μL 106CFU/mL 的菌液加入到无菌48 孔板中,分别加入100 μL 不同浓度的防腐剂,空白组加入100 μL 无菌生理盐水,将48 孔板放置于25 ℃培养箱内,测定培养0和24 h 时595 nm 波长下的吸光度值, 计算各组的ΔOD595nm值,并按下列公式(1),表征防腐剂的抗菌率。
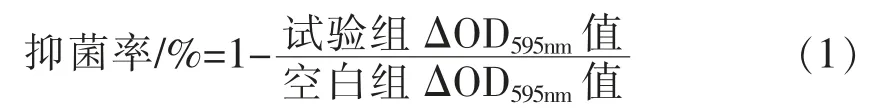
1.4 数据处理
统计学分析应用SPSS 12.0 软件,用平均值±标准差的形式表示数据结果,其中P<0.05 为显著性差异。 利用OriginPro 8.5 软件进行作图。
2 结果
2.1 冷藏河鲀鱼片腐败菌的分离鉴定
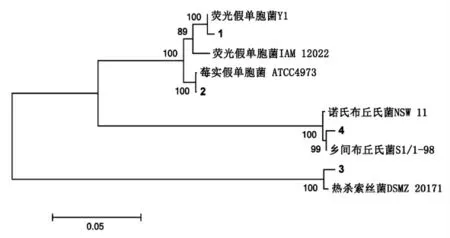
图1 基于16s rDNA 序列同源性的4 株腐败菌系统发育树Fig.1 Phylogenetic tree of PCR-amplified bacteria 16s rDNA gene phylotypes from four spoilage bacteria
冷藏河鲀鱼片腐败末期, 在稀释度为105的营养琼脂平板上,根据肉眼观察菌落形态、菌落颜色、隆起状态、边缘和表面状态,筛选出存在差异的4 株菌落,进行分离纯化,并分别标记为1,2,3和4 号, 经革兰氏染色判定1,2 和4 号细菌为革兰氏阴性细菌,3 号为革兰氏阳性细菌。
利用16s rDNA 测序技术, 将4 株腐败菌的16s rDNA 序列与相似菌株构建进化树,结果如图1 所示, 腐败菌菌株1,2,3 和4 号分别为荧光假单胞菌(Pseudomonas fluorescens)、莓实假单胞菌(Pseudomonas fragi)、 热杀索丝菌(Brochothrix thermosphacta)和乡间布丘氏菌(Buttiauxella gaviniae), 在Genbank 中的登录号分别为KY465667,KY465668,KY465669 和KY465670。
2.2 冷藏河鲀鱼片腐败菌致腐能力分析
冷藏河鲀鱼片分别接种腐败菌后, 以其在贮藏过程中TVBN 值和TMA 值的增长趋势来衡量4 株腐败菌的致腐能力。 如图2a 所示,在冷藏的第3 天,接种荧光假单胞菌和莓实假单胞菌的河鲀鱼片TVBN 值分别为 (8.95±0.06)mg/100 g 和(8.31±0.09)mg/100 g 显著性高于空白组 (6.51±0.13)mg/100g,而接种热杀索丝菌和乡间布丘氏菌的河鲀鱼片在第5 天的TVBN 值分别达到(8.27±0.03)mg/100g 和(8.15±0.05)mg/100 g 显著性高于空白组TVBN 值(7.00±0.16)mg/100 g(P<0.05)。
由图2b 可知,接种腐败菌的河鲀鱼片贮藏第1 天的TMA 值均显著性高于空白组,随着贮藏时间的延长,各组TMA 值逐渐升高,在第5 天时,接种荧光假单胞菌、莓实假单胞菌、热杀索丝菌和乡间布丘氏菌的河鲀鱼片TMA 值已分别达到(2.20±0.00),(1.86±0.06),(1.71±0.01)和(1.54±0.02)mg/100 g 显著性高于空白组(1.16±0.02)mg/100 g(P<0.05)。
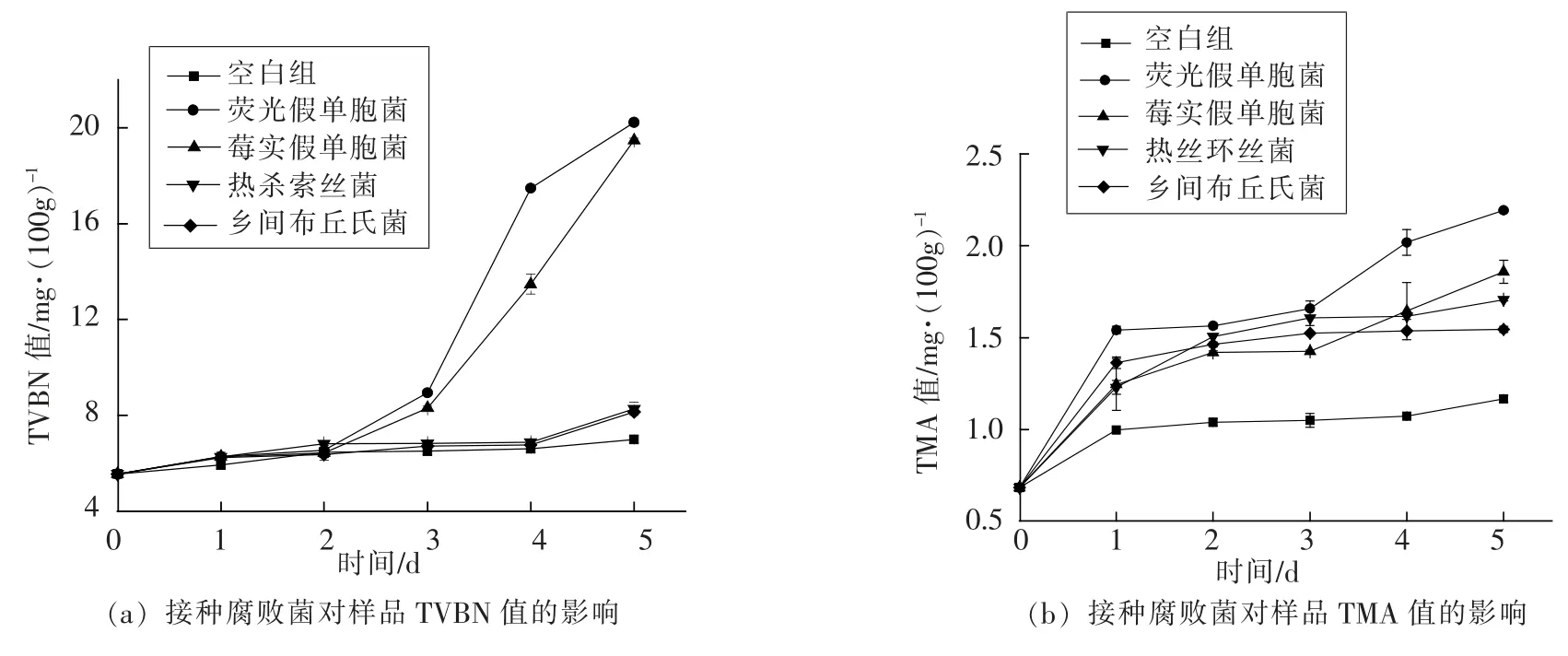
图2 冷藏河鲀鱼片4 株腐败菌的致腐能力Fig.2 Spoilage capability of four kinds of spoilage bacteria in chilled puffer fish
2.3 基质条件对4 株腐败菌生长量的影响
由图3 可知, 源于冷藏河鲀鱼片的腐败菌荧光假单胞菌、 莓实假单胞菌和热索环丝菌在液体培养基基质条件下的ΔOD590nm分别为0.89±0.03,1.16±0.01 和0.33±0.01,均显著高于在鱼肉汁基质中各自的ΔOD590nm(0.82±0.01,0.81±0.04 和0.14±0.01)(P<0.05)。 2 种培养基质对乡间布丘氏菌的生长量并无显著性的影响(P>0.05)。
2.4 不同基质条件下山梨酸钾对4 株腐败菌的抑菌活性
山梨酸钾溶液对荧光假单胞菌的抑菌效果如图4a 所示,在2 种基质条件下,7.5 mg/mL 的山梨酸钾溶液有效抑制荧光假单胞菌的生长繁殖,但抑菌率随着山梨酸钾浓度的降低而减少;0.47~3.75 mg/mL 的山梨酸钾溶液在鱼肉汁基质中对荧光假单胞菌的抑菌率显著性高于在液体培养基基质条件中(P<0.05)。 如图4b 所示,0.47~7.5 mg/mL的山梨酸钾溶液对莓实假单胞菌的抑菌率在鱼肉汁条件下均显著性高于液体培养基条件下(P<0.05)。

图3 不同基质条件对4 株腐败菌生长量的影响Fig.3 The growth of four kinds of spoilage bacteria under different matrix
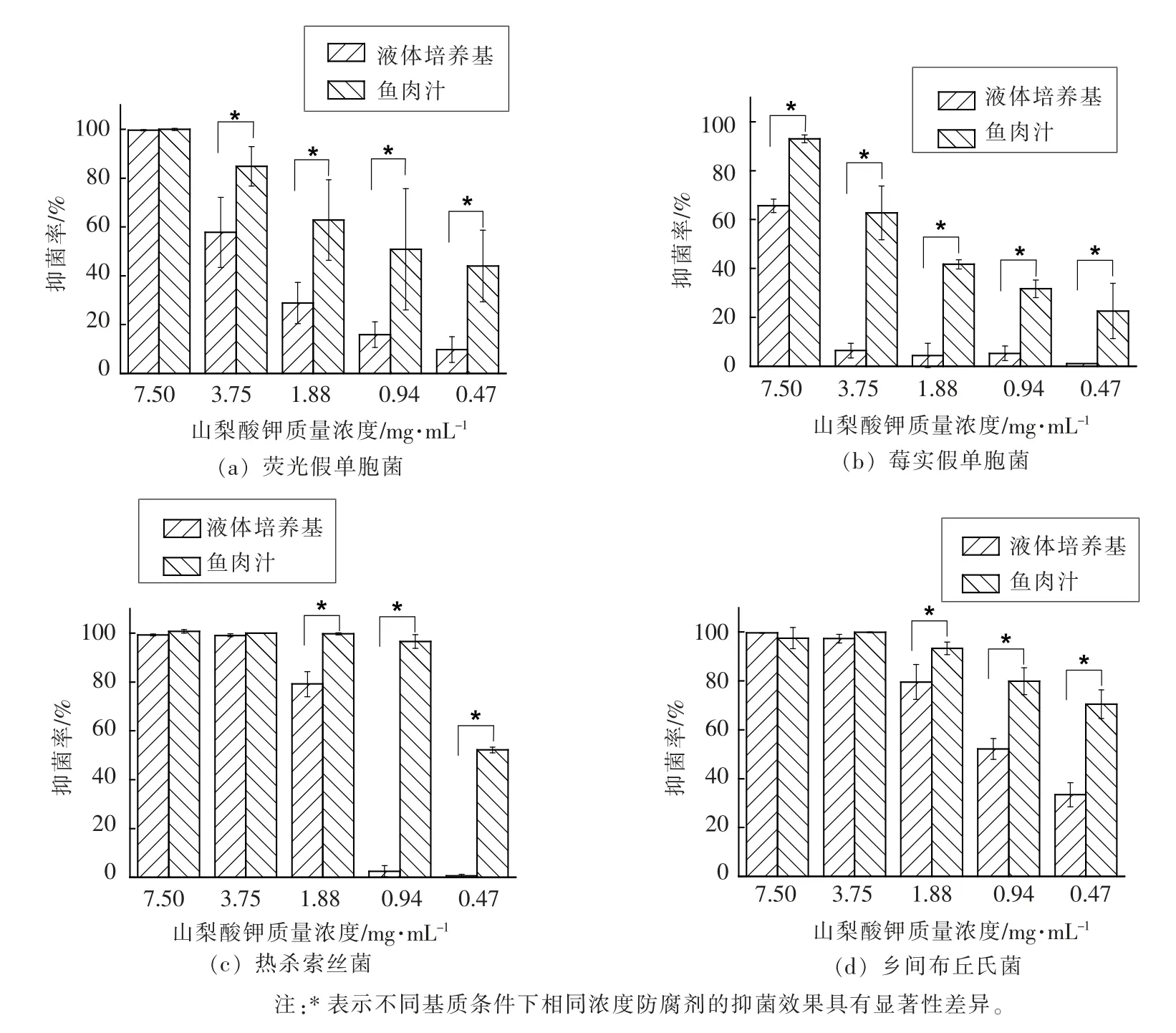
图4 不同基质条件下不同浓度山梨酸钾溶液对4 株腐败菌的抑菌活性Fig.4 Antibacterial activities of potassium sorbate with different concentration against four kinds of spoilage bacteria under different matrix
如图4c,图4d 所示,3.75 mg/mL 和7.5 mg/mL的山梨酸钾溶液能够完全抑制液体培养基和鱼肉汁基质中热杀索丝菌和乡间布丘氏菌的生长繁殖,但0.47~1.88 mg/mL 的山梨酸钾溶液对液体培养基中的热杀索丝菌和乡间布丘氏菌抑制效果要显著差于鱼肉汁基质条件(P<0.05)。
2.5 不同基质条件下尼泊金乙酯对4 株腐败菌的抑菌活性
尼泊金乙酯对冷藏河鲀鱼片腐败菌的抑制效果如图5 所示,0.05~0.80 mg/mL 的尼泊金乙酯均能够有效抑制荧光假单胞菌(图5a)、莓实假单胞菌(图5b)、热杀索丝菌(图5c)和乡间布丘氏菌(图5d)的生长繁殖。 液体培养基和鱼片基质未对尼泊金乙酯活性产生显著性的影响(P>0.05)。
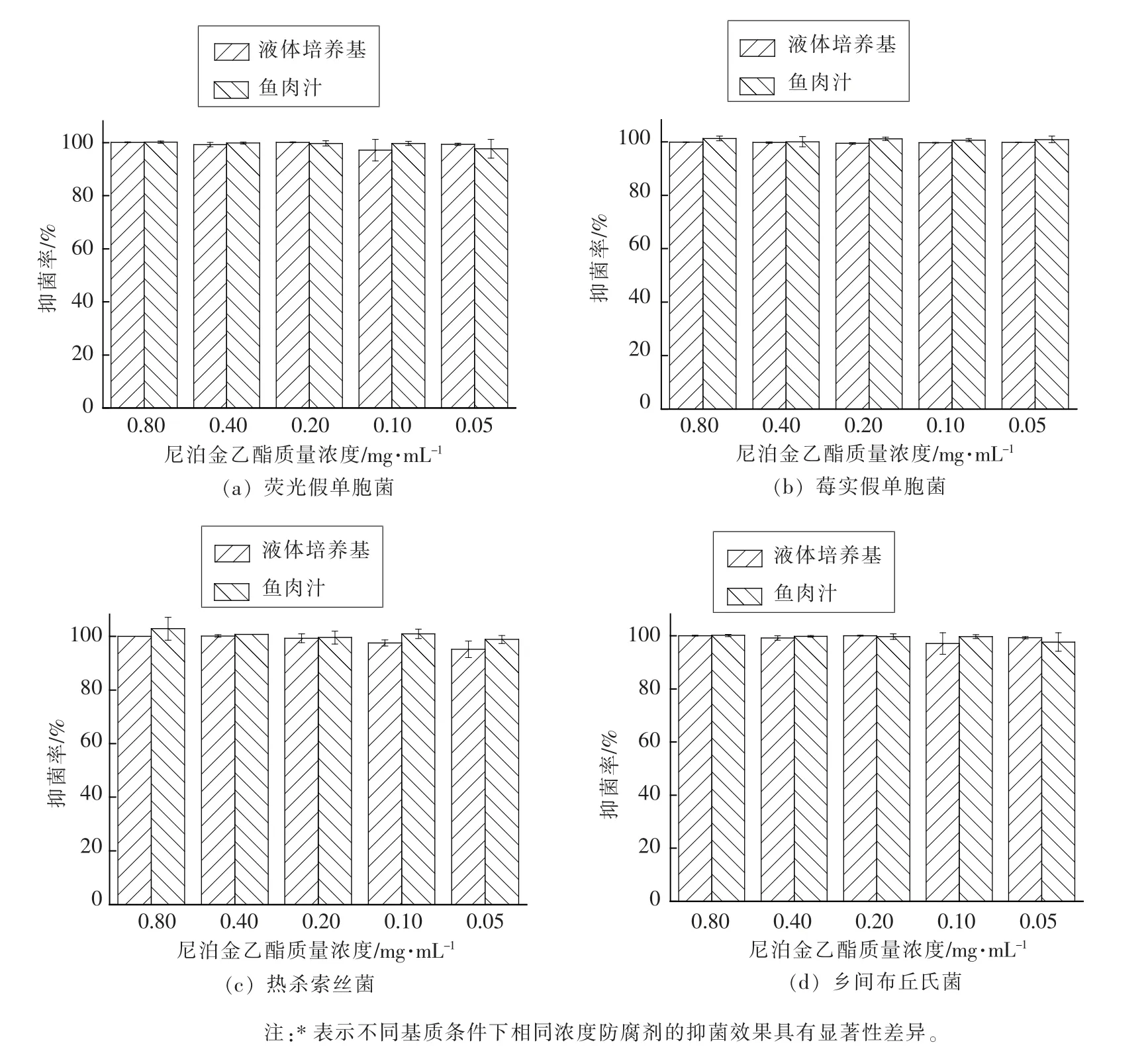
图5 不同基质条件下不同浓度尼泊金乙酯对4 株腐败菌的抑菌活性Fig.5 Antibacterial activities of ethyl hydroxybenzoate with different concentration against four kinds of spoilage bacteria under different matrix
2.6 不同基质条件下Nisin 溶液对4 株腐败菌的抑菌活性
不同浓度的Nisin 溶液对革兰氏阳性细菌热杀索丝菌有抑菌效果, 然而不能抑制革兰氏阴性细菌荧光假单胞菌、 莓实假单胞菌和乡间布丘氏菌的生长繁殖,因此本文仅列出Nisin 对热杀索丝菌的抑菌结果。 由图6 可知, 随着浓度的降低,Nisin 对液体培养基中的热杀索丝菌抑菌率逐渐降低,但0.015 mg/mL 的Nisin 溶液对鱼肉汁中的热杀索丝菌抑菌率仍高达97%。0.015~0.25 mg/mL的Nisin 溶液对鱼肉汁中热杀索丝菌抑菌率要显著优于液体培养基基质(P<0.05)。
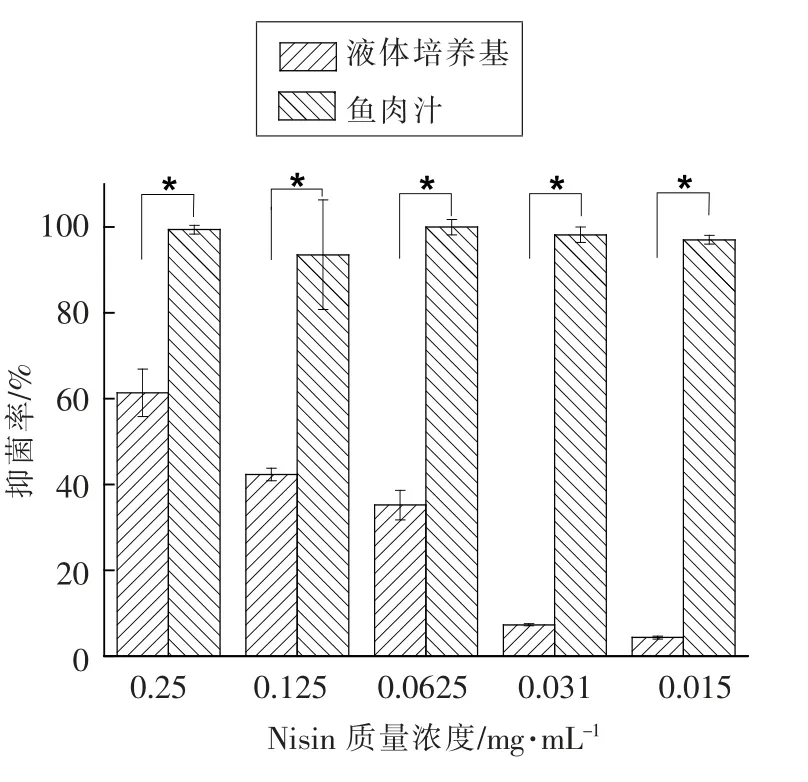
图6 不同基质条件下不同浓度Nisin 溶液对热杀索丝菌的抑菌活性Fig.6 Antibacterial activities of Nisin with different concentration against Brochothrix thermosphacta under different matrix
3 讨论
在不同贮藏条件下, 导致水产品腐败变质的腐败菌种类不同[12]。 在有氧冷藏条件下,假单胞菌属,特别是荧光假单胞菌、莓实假单胞菌和恶臭假单胞菌(P. putida),在贮藏过程中会逐渐占据优势地位并代谢产生感官不可接受的代谢产物,导致生鲜水产品腐败变质[13-14];而嗜冷型热索环丝菌容易在水产品中生长繁殖, 并在有氧等适宜条件下成为优势细菌,造成产品腐败变质[15];也有研究表明, 乡间布丘氏菌为导致有氧冷藏草鱼肉腐败变质的优势腐败菌[16]。 与前人的研究一致,本研究中分离获得的荧光假单胞菌、莓实假单胞菌、热索环丝菌和乡间布丘氏菌4 株腐败菌均能够加速腐败进程, 是导致冷藏河鲀鱼片产品腐败变质的关键腐败菌。
由于通过食品防腐剂抑制腐败菌的生长繁殖,能够对水产品进行有效的防腐保鲜,因此本文选取不同类型的防腐剂对冷藏河鲀鱼片的4 株腐败菌进行抑菌试验分析。 酸性防腐剂山梨酸钾抑菌活性易受环境pH 的影响[17],为排除基质中pH对抑菌效果的影响, 本研究中将液体培养基和鱼肉汁的pH 调节一致, 但2 种基质下山梨酸钾的抑菌效果仍具有显著差异, 即使对2 种基质下生长量相同的乡间布丘氏菌, 山梨酸钾在鱼肉汁中的抑菌活性优于液体培养基基质, 这说明除pH和细菌生长量因素外, 山梨酸钾的活性也易受抑菌环境基质成分组成和结构的影响。
尼泊金乙酯在液体培养基条件下对模式腐败菌藤黄微球菌质量(Mcirococcus luteus)、枯草芽孢杆菌(Bacillus subtilis)和大肠杆菌(Escherichia coli)的最小抑菌质量浓度分别为0.8,0.1 和0.1 mg/mL[18];对于化妆品中分离的铜绿假单胞菌(Pseudomonas aeruginosa)和大肠杆菌最低抑菌质量浓度均为2.5 mg/mL[19];然而,本研究中更低质量浓度的尼泊金乙酯溶液(0.05 mg/mL)对液体培养基和鱼肉汁基质中的4 株源于冷藏河鲀鱼片的腐败菌均能有效抑制。 这反映出不同菌株对尼泊金乙酯的耐受差异性较大, 也表明冷藏河鲀鱼片腐败菌的生长繁殖更易被尼泊金乙酯抑制。
生物防腐剂Nisin 在水产品保鲜中具有较好抑菌效果,但是它的抑菌谱有局限性,对革兰氏阴性细菌或真菌无抑制作用[20]。 这与本研究中的结果一致,Nisin 对荧光假单胞菌、 莓实假单胞菌和乡间布丘氏菌均无显著的抑制效果, 但能够有效抑制热杀索丝菌的生长繁殖。 Nisin 在食品基质中稳定性差,与脂肪、蛋白质等成分容易发生反应,导致Nisin 抑菌效力降低[21-22]。 本研究中Nisin 对液体培养基和河鲀鱼肉汁基质中的热杀索丝菌具有显著性的抑菌活性差异, 一方面可能是由于热杀索丝菌在鱼肉汁中的生长量要显著小于液体培养基基质中, 另一方面由于液体培养基中的蛋白胨和牛肉浸膏等蛋白质成分对Nisin 的吸附影响,导致Nisin 对液体培养基中的腐败菌抑菌活性下降。
4 结论
导致冷藏河鲀鱼片腐败变质的腐败菌有荧光假单胞菌、莓实假单胞菌、热杀索丝菌和乡间布丘氏菌。 本文选取的尼泊金乙酯和山梨酸钾防腐剂均能较好地抑制4 株腐败菌的生长繁殖,但Nisin仅对热杀索丝菌具有抑菌效果。 液体培养基和鱼肉汁基质条件下的抑菌结果表明,0.05 mg/mL 尼泊金乙酯能够有效抑制腐败菌的生长繁殖, 且不易受基质环境的影响;然而,山梨酸钾和Nisin 的抑菌活性受基质环境的影响, 对鱼肉汁中腐败菌的抑菌率显著性高于液体培养基条件。因此,尼泊金乙酯和山梨酸钾具有抑制冷藏河鲀鱼片产品中腐败菌生长繁殖的能力, 后续结合冷藏河鲀鱼片产品的基质条件, 可选取合适的浓度和比例用于制备复配保鲜剂延长产品的货架期。
猜你喜欢
食品与发酵工业(2022年4期)2022-02-25 08:45:36
化工生产与技术(2021年1期)2021-03-25 01:15:10
食品与生物技术学报(2021年6期)2021-01-15 22:39:59
食品与生物技术学报(2020年3期)2020-05-19 02:03:50
饮食与健康·下旬刊(2018年3期)2018-04-11 09:29:14
环球时报(2018-03-02)2018-03-02 05:35:05
海南热带海洋学院学报(2015年5期)2015-03-14 09:50:59
吉首大学学报(自然科学版)(2015年4期)2015-02-13 07:39:42
中华皮肤科杂志(2014年4期)2014-12-19 12:55:46
中国人兽共患病学报(2014年9期)2014-04-02 06:55:30
