
我国水稻田稗属杂草种类分布特点:以9个省级行政区73个样点调查数据为例
2019-07-29 02:30:28陈国奇唐伟李俊陆永良董立尧
中国水稻科学 2019年4期
陈国奇 唐伟 李俊 陆永良 董立尧,*
我国水稻田稗属杂草种类分布特点:以9个省级行政区73个样点调查数据为例
陈国奇1, 2,#唐伟3,#李俊4陆永良3,*董立尧4,*
(1江苏省作物遗传生理重点实验室/江苏省作物栽培生理重点实验室/扬州大学 农学院,江苏 扬州 225009;2江苏省粮食作物现代产业技术协同创新中心/扬州大学,江苏 扬州 225009;3中国水稻研究所 水稻生物学国家重点实验室,杭州 310006;4南京农业大学 植物保护学院/农作物生物灾害综合治理教育部重点实验室, 南京 210095;#共同第一作者;*通讯联系人,E-mail: dly@njau.edu.cn; luyongliang@caas.cn)
【】明确我国水稻田发生危害的稗属(spp.)杂草种类分布特点。2015年至2018年间的9月至11月,在东北、华东、华南、西北、西南水稻产区依据《中国植物志》英文修订版的分类鉴定标准,于水稻收割前采用七级目测法调查了73个地点共525块稻田稗属杂草的发生情况。共发现8种稗属杂草,其中无芒稗(var.)的发生频度最大(55.43%),其次是硬稃稗(,47.28%)、稗原变种(var.,40.40%)、西来稗(var.,24.09%);长芒稗()、水田稗()、孔雀稗()、光头稗()的频度较低。东北地区稻田稗属杂草以水田稗为单优势种;华东地区稻田稗属杂草呈现共优势的格局,硬稃稗、无芒稗危害最重,西来稗、稗原变种发生量也较大;华南地区稻田以稗原变种为单优势种;西北地区稻田无芒稗和稗原变种为优势种;西南地区稻田无芒稗占优势,稗原变种危害频繁。在这8种水稻田常见稗属杂草中,无芒稗的生态位宽度值最大(35.2),其次是硬稃稗(23.6)、西来稗(18.2)和稗原变种(16.3)。无芒稗、硬稃稗、西来稗相互之间的生态位重叠值较高,稗原变种与孔雀稗、光头稗、长芒稗的重叠值也较高。我国不同地区具有不同的稻田稗属杂草种类分布格局,其背后的机理亟待研究阐明。
稻田;稗属杂草;田间调查;优势种;生态位
稗属(spp.)杂草是水稻田危害严重的一类C4杂草,稗属杂草与水稻亲缘关系较近,不仅形态相似,在生态习性方面也非常相似[1],与水稻伴生会形成激烈的资源竞争,并可导致水稻严重减产,甚至绝收[2, 3]。稗属杂草防控是水稻田草害治理的重点,也是水稻田除草剂研发的最重要靶标杂草。我国水稻田登记使用的杀禾本科杂草除草剂中的绝大多数品种在登记试验和产品使用说明书中都涉及稗属杂草,如丙草胺、乙草胺、丁草胺、苯噻酰草胺、噁草酮、二氯喹啉酸、五氟磺草胺、精噁唑禾草灵、噁唑酰草胺、双草醚、嘧啶肟草醚、西草净、扑草净、异噁草松、乙氧氟草醚、禾草敌、禾草丹、氟酮磺草胺、双唑草腈、莎稗磷、敌稗、氯氟吡啶酯、双环磺草酮等(http://www.china pesticide.gov.cn/hysj/index.jhtml)。然而,不同种类的稗属杂草对水稻田生境的适应特性不尽相同,对水稻生产造成的危害也不相同,甚至差别极大[4-10]。因此,在水稻田稗草治理中,明确水稻田稗草种类组成情况兼具有理论研究价值和指导防控实践的意义。
稗属不同杂草之间形态高度相似[11-13],在我国“稗草”一词存在歧义,一种意思是指稗属杂草,另一种意思是单指稗()这一种。《中国植物志》中文版对稗属杂草进行了详细的描述并给出了检索表。2006年,《中国杂草志》英文修订版(http://foc.eflora.cn/)第22卷发布,对我国稗属植物进行了进一步修订,并给出了较为明确、清晰的检索分类依据(http://foc.eflora.cn/content.aspx? TaxonId =111218),为稗属杂草研究奠定了分类知识基础。在我国,多数关于水稻田杂草群落调查的文献只提及稗草复合群,而未细分具体的种类,少数文献提及了不同的稗属杂草种类但没有相关的田间发生量数据,进而难以有效支持水稻田稗属杂草种类演替和种群生态学等方面的研究。2015年至2018年间,我们在东北、西北、西南、华东、华南多个水稻产区于水稻收割前展开田间稗属杂草危害的调查,明确我国水稻田发生危害的稗属杂草种类分布特点,以期为我国水稻田稗属杂草治理对策和稗属杂草生物学、生态学研究提供重要的基础数据和参考。
1 材料与方法
1.1 稗属杂草种类鉴别
《中国杂草志》英文修订版的编研工作对《中国植物志》进行了全面的修订,历时25年,于2013年由中国、美国、英国、法国世界各地植物学家合作完成,是目前世界上最大、水平最高的英文版植物志(http://foc.eflora.cn/)。因此,本研究完全参照《中国杂草志》英文修订版资料进行稗属杂草的种类鉴别。
根据《中国植物志》英文修订版,我国稗属植物共有8个种,其中6种为常见杂草,包括稗()、硬稃稗(var.)、水田稗()、光头稗()、孔雀稗()、长芒稗()。此外,稗又有6个亚种均为杂草,包括稗原变种(var.)、无芒稗(var.)、西来稗(var.)、小旱稗(var.)、短芒稗(var.)、细叶旱稗(var.)。修订版《中国植物志》将旧版(http://frps.eflora.cn/)中的旱稗()并入稗原变种中;将水稗()并入水田稗中。
因此,《中国植物志》英文修订版确认的稗属杂草共有11个种或亚种。这11种稗属杂草中,常见于水稻田的稗属杂草包括8种:硬稃稗、水田稗、稗原变种、无芒稗、西来稗、光头稗、孔雀稗、长芒稗。这8种我国水稻田常见稗属杂草形态相似,主要通过小穗性状进行区分(图1)。参照《中国植物志》英文修订版,我国水稻田常见的8种稗属杂草的主要区分方法如下(表1):
1)硬稃稗:第一外稃(下外稃)腹面外凸,革质,较硬,发亮(其他稗属杂草第一外稃腹面扁平,草质,较软,不反光)。
2)水田稗:小穗大,长3.8~6.0 mm(同一花序上大的小穗长可超过5 mm),叶鞘鞘口有时有毛,秆直立,分蘖紧密簇生(其他稗属杂草小穗较小,2~4 mm,秆常披散,疏松成丛)。
A-硬稃稗小穗背面(左)和腹面(右);B-稗原变种小穗背面(左)和腹面(右);C-水田稗幼苗(红圈示鞘口有毛);D-硬稃稗幼苗(红圈示鞘口无毛)。1-第一颖片;2-第二颖片(紧贴其内的为第二外稃);3-第一外稃; A、B中红色的比例尺表示1 cm。
A, Back (left) and ventral (right) spikelet of; B, Back (left) and ventral (right) spikelet ofvar.; C, Hairs on abaxial side at junction of sheath and blade of; and D, Glabrous on abaxial side at junction of sheath and blade of. 1, Lower glume; 2, Upper glume (appressed inside it is the upper lemma); 3, Lower lemma. The scales in red in A and B suggest 1 cm.

图1 稗属杂草主要的种类鉴别特点
Fig. 1. Main identifying keys ofspecies.

表1 我国水稻田常见的8种稗属杂草形态区分
A,var.; B,; C,; D,; E,; F,var.; G,var.; H,., The lower cuspidate to awned; RS, Racemes with some secondary branching. The same as below.
3)光头稗:圆锥花序主轴通常无疣基长毛,总状花序长1~2 cm,小穗常成4行排列于穗轴的一侧,小穗无芒,长2~3 mm。
4)孔雀稗:圆锥花序分枝(总状花序)具有明显的小分枝,小穗长2.0~3.5 mm,第一外稃顶端具1.0~1.5 cm长的芒。
5)长芒稗:圆锥花序暗紫色,主轴疏被疣基长毛,小穗常带紫色,长3~4 mm,第一外稃顶端具1.5~5.0 cm长的芒。
6)稗原变种:圆锥花序主轴粗糙或具疣基长刺毛,小穗长3~4 mm,同一株上小穗第一外稃的芒长短不一,长0.5~1.5 cm(甚至达3 cm)。
7)无芒稗:圆锥花序直立,分枝硬挺,分枝具有明显的小分枝,小穗长3~4 mm,无芒或具有不超过5 mm的尖头。
8)西来稗:圆锥花序直立,分枝上无小分枝,小穗长3~4 mm,无芒或具有不超过5 mm的尖头。
1.2 调查区域和调查方法
2015年至2018年间的9月-11月,我们在江苏、安徽、上海、浙江、黑龙江、广东、贵州、宁夏、新疆于各地水稻收割前开展稻田稗属杂草种类调查。调查前先与较为熟悉的当地农科院、植保站、农药经销商等联系,确定水稻田稗属杂草发生较重的地点作为调查点并落实当地向导,具体调查点主要为当地水稻田稗属杂草发生较重的区域。每个调查点选定稗属杂草较多的5块或10块水稻田开展田间调查,每块田取667 m2左右的调查面积。总共在9个省级行政区调查了73个点共525块水稻田,包括黑龙江3个调查点30块田,江苏20个调查点160块田,安徽15个调查点115块,上海4个调查点40块,浙江9个调查点70块,广东6个调查点30块,宁夏4个调查点20块,新疆6个调查点30块,贵州6个调查点30块。
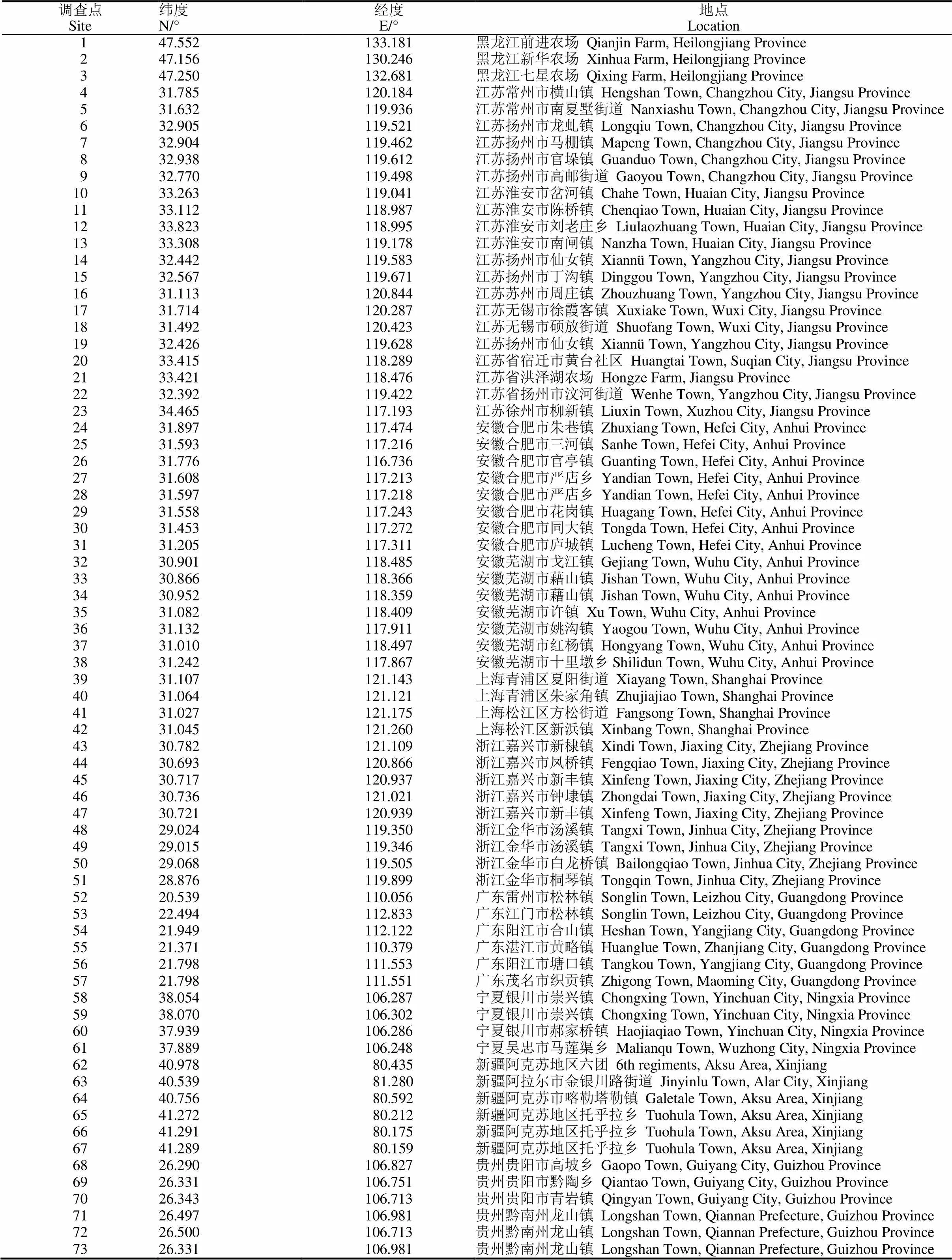
表2 本研究73个调查点的位置信息
根据《中国杂草志》英文修订版对田间观察到的每种稗属杂草进行种类鉴定,并采用七级目测法记录每一块水稻田稗属杂草的优势度值[14, 15]。田埂上和田边的稗属杂草不计。
1.3 数据统计分析
所有数据展示格式均为平均值±标准误。基于8种稗属杂草在全部调查田块的出现次数,计算其频度(出现田块数/525×100%)。对照七级目测法确定每种稗属杂草在每块调查田块中的优势度级别,计算每种稗属杂草在每个调查点的综合值[15-17],综合值=(该优势度等级出现田块数×该级代表值)/(田块总数×该优势度等级最高代表值)×100%)。建立“调查点-稗属杂草综合值”数据矩阵。将调查所开展的9个省级行政区分为5个地区进行比较分析,包括东北(黑龙江)、华东(江苏、安徽、上海、浙江)、华南(广东)、西北(宁夏、新疆)、西南(贵州)。基于“调查点-稗属杂草综合值”数据矩阵,采用Levins方法计算各种稗属杂草的生态位宽度值及两两之间的生态位重叠值[18-20]。
2 结果与分析
2.1 水稻田稗属杂草发生频度
依据《中国杂草志》英文修订版的分类标准,在9个省级行政区的525块水稻田中记录到硬稃稗、水田稗、稗原变种、无芒稗、西来稗、光头稗、孔雀稗、长芒稗等8种我国水稻田常见稗属杂草(表3)。在所调查525块稗属杂草发生的水稻田中,无芒稗的发生频度最大(55.43%),其次是硬稃稗(47.28%)、稗原变种(40.40%)、西来稗(24.09%),长芒稗、水田稗、孔雀稗、光头稗的频度较低(表3)。在东北地区的黑龙江,所有调查的水稻田中均有水田稗,稗原变种、长芒稗、无芒稗也较常见。在华东地区的江苏、安徽、上海、浙江,无芒稗、硬稃稗、稗原变种、西来稗较为常见,长芒稗、孔雀稗频度较低,光头稗偶尔发生。在华南地区的广东,所有调查稻田均出现稗原变种,无芒稗的频度超过80%,孔雀稗、硬稃稗和长芒稗偶见于稻田。在西北地区的宁夏和新疆,无芒稗、稗原变种最为常见,水田稗、硬稃稗偶发,孔雀稗在宁夏稻田较常见。在西南的贵州,所调查的30块水稻田只发现了无芒稗和稗原变种。
2.2 水稻田稗属杂草的发生量
在所调查的73个点525块稻田中,无芒稗的综合值(11.4)最高(表4),其次是硬稃稗(9.5)、稗原变种(7.3)、西来稗(5.5)、水田稗(3.1)、孔雀稗(1.0)、长芒稗(0.7)、光头稗(0.2)。东北地区水田稗呈单优势,其发生的综合值接近72。华东地区硬稃稗综合值(14.3)最大,无芒稗的综合值(12.3)较硬稃稗略低,西来稗和稗原变种的综合值也较大。华南地区稗原变种为优势稗属杂草,无芒稗和孔雀稗在少数田块重度危害。西北地区无芒稗综合值(9.0)最大,其次是稗原变种,水田稗和孔雀稗有时发生量较大。西南地区以无芒稗为主,稗原变种的发生量也较大。
2.3 水稻田稗属杂草的生态位宽度和生态位重叠
生态位宽度反映所研究物种在所调查区域内的分布和适应能力,分布越广、发生量越大则生态位宽度值越大(表4)。在这8种水稻田常见稗属杂草中,无芒稗的生态位宽度值最大(35.2),其次是硬稃稗(23.6)、西来稗(18.2)和稗原变种(16.3),长芒稗、水田稗、光头稗和孔雀稗在所调查稻田的生态位宽度值均较小。
表5 调查水稻田中8种稗属杂草的生态位重叠值

Table 5. Pairwise niche overlaps among the eight Echinochloa species on surveyed rice fields.
生态位重叠值反映物种两两之间的分布相似性情况,重叠值越大反映两两之间的分布特点越接近。稗原变种与孔雀稗的重叠值最大,为0.99(表5);稗原变种与光头稗、长芒稗的重叠值超过0.30;无芒稗、硬稃稗、西来稗相互之间的重叠值也较高(0.33~0.45);此外,硬稃稗与长芒稗的重叠值也较高,为0.27。
3 讨论
本研究基于9个省级行政区73个点的调查数据表明,我国不同地区具有不同的稻田稗属杂草种类分布格局。东北地区稻田稗属杂草以水田稗为单优势;华东地区稻田稗属杂草呈现共优势的格局,硬稃稗、无芒稗危害最重,西来稗、稗原变种发生量也较大;华南地区稻田以稗原变种为单优势;西北地区稻田无芒稗和稗原变种为优势;西南地区稻田无芒稗占优势,稗原变种危害频繁。
在我国,水田稗仅在东北地区稻田发生严重危害,这可能与该种对东北地区稻田温度较低和一年一熟制的栽培模式密切相关。水田稗原产于亚洲,是亚洲、美洲和欧洲水稻田的主要杂草[21-23]。Boddy等[22]研究了水田稗的种子萌发生物学特性,发现水田稗种子萌发的基础温度是9.5°C,最适宜萌发温度是31°C,总体而言适宜萌发温度低于稗。在东北地区,田间水稻通常在5月份移栽,9月份收割,这期间温度有利于水田稗的萌发和生长。有研究比较了8个水田稗种群和4个稗草()种群的生物学特性[10],发现水田稗种群的出苗速率显著快于稗草种群,出苗整齐性高于稗草种群,这与文献[24]报道的水田稗种子休眠较稗草种子休眠期短,出苗整齐性高一致,因此更适宜一年一季的栽培模式。西北地区的部分调查田块(10%)也发现水田稗危害,西北地区水稻田也为一年一季模式,但总体而言,水田稗危害低于东北地区稻田,具体原因有待进一步研究。而在华东、华南和西南地区,一年多熟制的轮作模式下,翻耕、播种、灌溉等农艺措施对土壤扰动较大,因此,不利于水田稗种子在土壤中的保存,因而有效阻碍了水田稗的危害。
硬稃稗生物学、生态学亟待更多的深入研究。硬稃稗是亚洲地区危害严重的恶性杂草,对水稻田生境高度适宜[3]。黑暗/光照下15℃/25℃、20℃/30℃、25℃/35℃均较适宜硬稃稗种子萌发,淹水0~6 cm条件下其种子也能正常萌发[25]。本研究发现硬稃稗仅在华东地区稻田危害严重,在华南和西北地区稻田偶见,其中的原因未知。华东地区稻田多为稻麦轮作田,而华南地区多为稻-菜轮作田,西北地区稻田为一年一熟制。因此,硬稃稗仅在华东地区危害严重可能与其对于稻麦轮作系统的高度适应性相关。硬稃稗在水稻田的成灾机制和防控方法仍需进一步研究。此外,有研究在安徽发现了抗除草剂噁嗪草酮和丁草胺的硬稃稗种群[26],这对稻田硬稃稗的防治构成了新的挑战。
本研究发现无芒稗在8种稻田常见稗属杂草中具有最大的适应辐,表现在远高于其他种类的生态位宽度值(表3),并且在所调查各个水稻种植区域均具有较高的发生量。无芒稗植株分蘖多且披散,圆锥花序具有二级分枝,种子量大[1],并且对稻田灌水和保水具有极强的适应性[27],在水稻田具有极强的竞争能力和适应性[28],在直播和移栽水稻田均为恶性杂草[29],甚至巴西的水稻田也深受其害[30]。此外,我国东北和华东地区均发现抗除草剂的无芒稗种群[26, 31, 32],为无芒稗的防控进一步增加了难度。
本研究发现所调查的稻田中,稗原变种的生态位宽度也较大,但除华南的调查点之外其他地区稗原变种的平均发生量不是最高的,在东北调查点的平均发生量远低于水田稗,在华东调查点的平均发生量低于硬稃稗、无芒稗和西来稗,在西北和西南调查点的发生量低于无芒稗。这可能与稻田除草剂开发时的靶标检测有关。稗是水稻田危害最重的恶性杂草之一,在全世界广泛报道[1, 2, 23],也是水稻田除草剂开发中最重要的靶标杂草之一,水稻田针对禾本科杂草的除草剂开发过程中,多数都会以稗草为供试对象,而其中最多的是稗原变种。
本次调查的525块水稻田中,光头稗的发生频度仅2.36%,表明该种对调查区域水稻田的适应能力有限。光头稗主要分布在南北纬30°以内的热带、亚热带区域,主要危害旱地作物,尤其是免耕旱地作物,在南亚地区一些旱直播稻田和移栽稻田也会发生严重危害[33]。光头稗生长迅速并且能在短期内结实,多数研究表明光头稗种子几乎没有休眠,种子在20℃~34℃条件即可萌发出苗,然而,埋土2 cm即可使光头稗种子出苗率低于20%。本次调查的东北、西北地区稻田气候对光头稗并不适宜,而华东、华南、西南地区稻田均为水稻与其他作物轮作,水稻播种前均会采取翻耕或者旋耕操作,因而,不利于光头稗在稻田发生。本研究也发现部分田间孔雀稗和长芒稗形成优势,但总体而言发生危害程度较低。孔雀稗喜深水田,深泥水田中常出现优势群丛[1]。长芒稗也喜湿,对10 cm以下的水层具有突出的适应性,是一些湿地中的恶性杂草,然而该种在水稻田发生恶性危害的现象并不常见。目前关于孔雀稗和长芒稗的生物学、生态学研究资料不足,稻田孔雀稗和长芒稗的成灾机制也需进一步开展研究。
国内已有一些文献报道我国稻田稗属杂草的分类及分布研究结果[34-36]。本研究揭示了我国不同地区73个调查点水稻田稗属杂草的种类分布特点。然而,到目前为止,我们仍然无法基于稗属杂草种类的准确鉴定来指导稻田稗属杂草的化学防控策略。一方面,尚没有系统性的研究明确不同种类的稗属杂草对同一种除草剂的敏感性差别;另一方面,同一种稗属杂草不同种群之间对同一种除草剂的敏感性差别极大[32, 37]。并且,我们在调查中也发现,多种稗属杂草共生于同一块水稻田的情况较为普遍。因此,为指导特定地区稻田稗属杂草的化学防控,基于当地稻田的稗属杂草种群持续进行对除草剂抗性监测十分必要。
当前,农药减量使用、保护农区生态环境已经成为我国的国家意志,稻田稗属杂草防控宜采取包括多种类型措施的综合防控技术[38],因此,研究不同稗属杂草的生物学、生态学特点,进而结合水稻栽培系统提出针对性的综合防控策略十分必要且紧迫。
谢辞:感谢上海师范大学郭水良教授、南京农业大学强胜教授及Bernal Valverde客座教授在稗属杂草分类中提供的帮助;感谢广东省农业科学院植保所田兴山研究员、冯莉研究员及郭文磊博士,黑龙江省农业科学院植保所黄春艳研究员,黑龙江省农垦总局植保植检站关成宏研究员,贵州省农业科学院植保所何永福研究员,安徽省农业科学院植保所张勇博士,周凤艳博士,芜湖市农业技术中心植保站汤凡清等老师,上海市青浦区农技推广服务中心金燕研究员,张琳老师,扬州市江都区植保站张友明研究员,盐城市阜宁县农业委员会郭丽华老师,淮安市农业科学院李茹研究员,付佑胜主任等,以及嘉兴市、金华市植保站和陶氏公司等众多单位诸多同志在田间调查采样前后提供的建议、向导和帮助。
[1] 李扬汉. 中国杂草志. 北京: 中国农业出版社, 1998: 1214-1221.
Li Y H. Weed Flora of China. Beijing: China Agriculture Press, 1998: 1214-1221. (in Chinese)
[2] Holm L G, Plucknett D L, Pancho J V, Herberger J P. The World’s Worst Weeds: Distribution and Biology. Honolulu: University Press of Hawaii, 1977.
[3] Rao A N, Johnson D E, Sivaprasad B, Ladha J K, Mortimer A M. Weed management in direct-seeded rice, in Advances in Agronomy. L.S. Donald: Academic Press, 2007: 153-255.
[4] Zhang Z, Gu T, Zhao B, Yang X, Peng Q, Li Y, Bai L. Effects of commonvarieties on grain yield and grain quality of rice, 2017, 203: 163-172.
[5] 张自常, 李永丰, 张彬, 杨霞. 稗属杂草对水稻生长发育和产量的影响应用生态学报, 2014, 25(11): 3177-3184.
Zhang Z C, Li Y F, Zhang B, Yang X. Influence of weeds inon growth and yield of rice., 2014, 25(11): 3177-3184. (in Chinese with English abstract)
[6] Ruiz-Santaella J P, Pujadas A, De Prado R. Botanical identification of several Echinochloa biotypes collected in Spanish rice fields with control failures of cyhalofop-butyl, 2003, 68(4a): 421-424.
[7] Aminpanah H. Competition between rice (L.) and barnyardgrass ((L.) P. Beauv.) or rice barnyardgrass (Vasing.), 2012, 29: 395-403.
[8] Kovach D A, Widrlechner M P, Brenner D M. Variation in seed dormancy inand the development of a standard protocol for germination testing, 2010, 38(3): 559-571.
[9] De Cauwer B, Rombaut R, Bulcke R, Reheul D. Differential sensitivity ofandto 4-hydroxyphenyl pyruvate dioxygenase- and acetolactate synthase-inhibiting herbicides in maize, 2012, 52(6): 500-509.
[10] Damalas C A, Dhima K V, Eleftherohorinos I G. Morphological and physiological variation among species of the genusin northern Greece, 2008, 56(3): 416-423.
[11] Chen G, Zhang W, Fang J, Dong L. Identification of massive molecular markers inusing a restriction-site associated DNA approach, 2017, 39(5): 287-293.
[12] Altop E K, Mennan H. Genetic and morphologic diversity ofpopulations from different origins, 2011, 39(1): 93-102.
[13] Tabacchi M, Mantegazza R, Spada A, Ferrero A. Morphological traits and molecular markers for classification ofspecies from Italian rice fields, 2006, 54(6): 1086-1093.
[14] Chen G Q, He Y H, Qiang S. Increasing seriousness of plant invasions in croplands of eastern China in relation to changing farming practices: A case study, 2013, 8(9): e 74136.
[15] Qiang S. Multivariate analysis, description, and ecological interpretation of weed vegetation in the summer crop fields of Anhui Province, China, 2005, 47(10): 1193-1210.
[16] 强胜, 王启雨, 谢定三. 安徽省霍邱县夏收作物田杂草群落的数量分析植物资源与环境, 1994, 3(2): 39-44.
Qiang S, Wang Q Y, Xie D S. Quantitative analysis of weed communities of summer crops in Huoqiu county of Anhui Province., 1994, 3(2): 39-44. (in Chinese with English abstract)
[17] 张峥, 戴伟民, 章超斌, 强胜. 江苏沿江地区杂草稻的生物学特性及危害调查. 中国农业科学, 2012, 45(14): 2856-2866.
Zhang Z, Dai W M, Zhang C B, Qiang S. Investigation of the biological characteristics and harmfulness of weedy rice (L. f.) occurred in the regions along the Yangtze River of Jiangsu Province., 2012, 45(14): 2856-2866. (in Chinese with English abstract)
[18] 陈国奇, 郭水良, 印丽萍. 外来入侵种植物学性状和环境因子间关系的典范对应分析. 浙江大学学报: 农业与生命科学版, 2008, 34(5): 571-577.
Chen G Q, Guo S L, Yin L P. Canonical correspondence analysis of relationship between botanical characters of exotic weeds and their environmental factors., 2008, 34(5): 571-577. (in Chinese with English abstract)
[19] Barbini S A, Sabadin D E, Lucifora L O. Comparative analysis of feeding habits and dietary niche breadth in skates: the importance of body size, snout length, and depth, 2018, 28(3): 625-636.
[20] 张孝然, 蒲真, 黄治昊, 周鑫, 邢韶华. 大黄花虾脊兰生境特征及生存群落物种生态位分析植物科学学报, 2017, 35(6): 799-806.
Zhang X R, Pu Z, Huang Z H, Zhou X, Xing S H. Habitat characteristics and niche analysis ofDecne. Survival community.2017, 35(6): 799-806. (in Chinese with English abstract)
[21] Mennan H, Kaya-Altop E. Molecular techniques for discrimination of late watergrass () and early watergrass () species in Turkish rice production, 2012, 60(4): 525-530.
[22] Boddy L G, Bradford K J, Fischer A J. Population-based threshold models describe weed germination and emergence patterns across varying temperature, moisture and oxygen conditions, 2012, 49(6): 1225-1236.
[23] Holm L G, Pancho J V, Herberger J P. A Geographical atlas of World Weeds. New York: John Wiley and Sons, 1979.
[24] Barrett S C H, Wilson B F. Colonizing ability in thecomplex (barnyard grass): I. Variation in life history, 1983, 61(2): 556-562.
[25] Opena J L, Chauhan B S, Baltazar A M. Seed germination ecology ofand its implication for management in rice (L.), 2014, 9(3): e92261.
[26] 王琼, 陈国奇, 姜英, 王庆亚, 姚振威, 董立尧. 水稻田稗属杂草(spp.)对稻田常用除草剂的敏感性南京农业大学学报, 2015, 38(5): 804-809.
Wang Q, Chen G Q, Jiang Y, Wang Q Y, Yao Z W, Dong L Y. Sensitivity ofspecies to frequently used herbicides in paddy rice field., 2015, 38(5): 804-809. (in Chinese with English abstract)
[27] 徐正浩, 王一平, 余柳青, Mortimer A M. 水稻移栽密度和早期水管理对无芒稗生态控制效果的影响生态学杂志, 2004, 23(4): 41-43.
Xu Z H, Wang Y P, Yu L Q, Mortimer A M. Effects of rice transplanting density and early irrigation management on barnyardgrass ecological control., 2004, 23(4): 41-43. (in Chinese with English abstract)
[28] 徐正浩, 何勇, 王一平, 俞谷松. 不同水层和密度条件下化感作用水稻对无芒稗的干扰控制作用应用生态学报, 2004, 15(9): 1580-1584.
Xu Z H, He Y, Wang Y P, Yu G S. Interference of allelopathic rice cultivars on barnyardgrass under different water irrigation and rice plant density.,2004, 15(9): 1580-1584. (in Chinese with English abstract)
[29] 徐正浩, 谢国雄, 周宇杰, 高屾. 三种栽植方式下不同株型和化感特性水稻对无芒稗的干扰控制作用作物学报, 2013, 39(3): 537-548.
Xu Z H, Xie G X, Zhou Y J, Gao S. Interference of rice with different morphological types and allelopathy on barnyardgrass under three planting patterns.,2013, 39(3): 537-548. (in Chinese with English abstract)
[30] Bonow J F L, Lamego F P, Andres A, Avila L A, Telo G M, Egewarth K. Resistance ofvar.to imazapyr plus imazapic herbicide and alternative control in irrigated rice, 2018, 36: e018168627.
[31] 董海, 蒋爱丽, 李林生, 杨皓, 刘晓舟, 李颖. 辽宁省无芒稗对二氯喹啉酸的抗药性研究北方水稻, 2007(6): 36-39.
Dong H, Jiang A L, Li L S, Yang H, Liu X Z, Li Y. Studies on resistance of(L.) Beauv. var.(Pursh) Peterm to Quinclorac in Liaoning., 2007(6): 36-39. (in Chinese with English abstract)
[32] 温广月, 钱振官, 李涛, 沈国辉, 刘益君. 上海地区稻田无芒稗对丁草胺抗药性的测定杂草科学, 2015, 33(3): 34-36.
Wen G Y, Qian Z G, Li T, Shen G H, Liu Y J. Identification ofvar.resistant to butachlor from paddy fields in Shanghai., 2015, 33(3): 34-36. (in Chinese with English abstract)
[33] Peerzada A M, Bajwa A A, Ali H H, Chauhan B S. Biology, impact, and management of(L.) Link, 2016, 83: 56-66.
[34] 陆永良, 刘德好, 余柳青, 刘都才, 郭水良. 中国主要农区稻田稗草分类与多样性研究. 植物科学学报, 2014, 32(5): 435-445.
Lu Y L, Liu D H, Yu L Q, Liu D C, Guo S L. Classification and diversity ofin paddy fields of main agricultural regions in China., 2014, 32(5): 435-445. (in Chinese with English abstract)
[35] 周平, 郭水良, 印丽萍. 稗属植物的分类与系统学研究概况. 杂草科学, 2013, 31(1): 1-4.
Zhou P, Guo S L, Yin L P. Status of research on taxonomy and systematics of the genus., 2013, 31(1): 1-4. (in Chinese with English abstract)
[36] 陆永良, 刘德好, 郭水良, 余柳青. 基于ISSR标记的中国稻田稗属植物分类. 浙江农业学报, 2014, 26(5): 1309-1314.
Lu Y L, Liu D H, Guo S L. Yu L Q. Classification ofspecies in Chinese paddy fields based on ISSR markers., 2014, 26(5): 1309-1314. (in Chinese with English abstract)
[37] Chen G, Wang Q, Yao Z, Zhu L, Dong L. Penoxsulam- resistant barnyardgrass () in rice fields in China, 2016, 16(1): 16-23.
[38] 董立尧, 王红春, 陈国奇, 付佑胜, 徐江艳, 张玉华, 于佳星, 刘庆虎. 直播稻田杂草防控技术. 北京:中国农业出版社, 2016: 111-133.
Dong L Y, Wang H C, Chen G Q, Fu Y S, Xu J Y, Zhang Y H, Yu J X, Liu Q H. Weed Management Strategies on Direct Seedling Rice Fields. Beijing: China Agriculture Press, 2016: 111-133. (in Chinese)
Distribution Characteristics ofSpecies in Rice Fields in China: A Case Survey on 73 Sites from Nine Provincial Administrative Regions
CHEN Guoqi1, 2,#, TANG Wei3,#, LI Jun4, LU Yongliang3,*, DONG Liyao4,*
(Jiangsu Key Laboratory of Crop Genetics and Physiology/ Jiangsu Key Laboratory of Crop Cultivation and Physiology/,,; Jiangsu Co-Innovation Center for Modern Production Technology of Grain Crops/,,; State Key Laboratory of Rice Biology,,,;College of Plant Protection/Key Laboratory of Integrated Management of Crop Disease and Pests,,,,;;Corresponding author,:;)
【】The purpose is to disclose the distributon patterns ofspecies in rice fields in China. 【】In periods from September to November between 2015 and 2018, we conducted field surveys on the occurrence ofspecies at 73 locations (totally 525 rice fields). 【】Totally eightspecies were observed in the 525 fields surveyed. Among these fields,var.showed the highest frequency (55.43%), followed by(47.28%),var.(40.40%),var.(20.94%); and,,,showed low frequency. On rice fields surveyed in Northeast China,was mono-dominant amongspecies on rice fields; in East China,andvar.were the most serious among this genus,var.andvar.also showed high occurrences; in South China,var.was mono-dominant; in Northwest China,var.andvar.were dominant; and in Southwest China,var.was dominant andvar.also showed high occurrence. Among the eightspecies observed,var.showed the highest niche breadth (35.2), followed by(23.6),var.(18.2) andvar.(16.3). Niche overlaps among,var.andvar.were high, as well as those betweenvar.and,or, respectively.【】Frequency ofspecies varied with areas, and the relative mechanisms entail further studies.
rice fields;species; field survey; dominant species; ecological niche
S451; S511
A
1001-7216(2019)04-0368-09
10.16819/j.1001-7216.2019.8137
2018-12-14;
2019-02-11。
国家重点研发计划资助项目(2018YFD0300802);江苏高校优势学科建设工程资助项目(PAPD)。
猜你喜欢
风流一代·青春(2022年7期)2022-07-22 22:57:11
意林·作文素材(2021年15期)2021-10-08 13:54:26
学生天地(2019年33期)2019-08-25 08:56:24
初中生世界·九年级(2016年12期)2017-01-06 14:43:17
幼儿智力世界(2016年2期)2016-05-30 10:48:04
幼儿智力世界(2016年1期)2016-05-30 10:48:04
西南农业学报(2016年5期)2016-05-17 05:42:32
小学生导刊(高年级)(2016年4期)2016-04-08 21:45:45
现代农业(2015年5期)2015-02-28 18:40:49
创新作文(1-2年级)(2014年10期)2015-01-13 02:38:09
