
邛海河-湖系统春季浮游植物磷胁迫研究
2019-07-25 08:21王雪梅赵刘平胡金朝
湖北农业科学 2019年13期
王雪梅,赵刘平,胡金朝,杨 红
(西昌学院资源与环境学院,四川 西昌 615013)
富营养化已经成为目前世界上大多数淡水和近海生态系统所面临的主要水质问题,而氮、磷等营养物质的大量输入是引起富营养化的主要原因[1]。淡水水体中最主要的限制性营养元素是磷,因而浮游植物对磷的生物可利用性是引发富营养化的关键因子[2]。在磷的众多形态中,浮游植物仅能够直接利用正磷酸盐,但其含量在水体中往往不足总磷的5%,因此会很快导致浮游植物的磷胁迫。在磷胁迫状态下,藻类可以通过改变种类组分或分泌提高碱性磷酸酶(AP)活性,以催化水解不能被生物制剂吸收利用的非活性磷,因而碱性磷酸酶活性(APA)的大小可以用于指示水体中磷限制或磷胁迫的状态。
四川省西昌市邛海入选国家优良自然湖泊,因此对邛海的保护也越来越受到重视。但近年来由于城市发展和人工湿地建设,使其汇水河流和湖区的水文、水力特征都发生了改变,对其营养盐平衡和浮游植物也会产生相应的影响。因此,本试验以邛海河-湖系统为研究对象,通过群落水平碱性磷酸酶活性分析的方法,研究其水体中浮游植物磷胁迫的状况,探讨邛海河-湖系统环境因子与磷酸酶活性的关系,旨在为邛海富营养化的防治提供一定的理论依据。
1 材料与方法
1.1 研究区域概况
邛海位于四川省西昌市南7 km处,是攀西高原上最大的淡水湖,面积约31 km2,流域内南高北低,湖泊南北分布,湖水平均深14 m,水位变幅小,年均水温17.8℃左右。邛海的汇水河流北有高沧河,东有官坝河,南有鹅掌河,次一级的河流有青河、大沟河、踏沟河及海河等。除官坝河、鹅掌河外,其余多为季节性河流,以上河流汇入邛海后,由海河排泄。流域内支沟、冲沟密布,长度大于1 km的支沟众多,水系密度达 0.68 条 /km2[3]。
1.2 样品采集
经现场踏查后,选取了邛海周边有明显径流的6条河流设置采样点,包括鹅掌河、踏沟河、青河、大沟河、官坝河和海河,因湿地三期封闭整修无法进入,故湿地三期范围内的河流未进行调查。分别在每条河流的下游、入湖河口和邻近湖区设置监测点,其中海河在上游和出海口,各个监测点的位置布局如图1所示。于2016年4月12日使用有机玻璃采水器进行水样采集,每个监测点采集样品两份,采样点间距离不小于50 m,同时现场测定水体的温度、pH和透明度[4],并做好记录。采集样品送入实验室,并放于4℃冰箱中冷藏待分析。
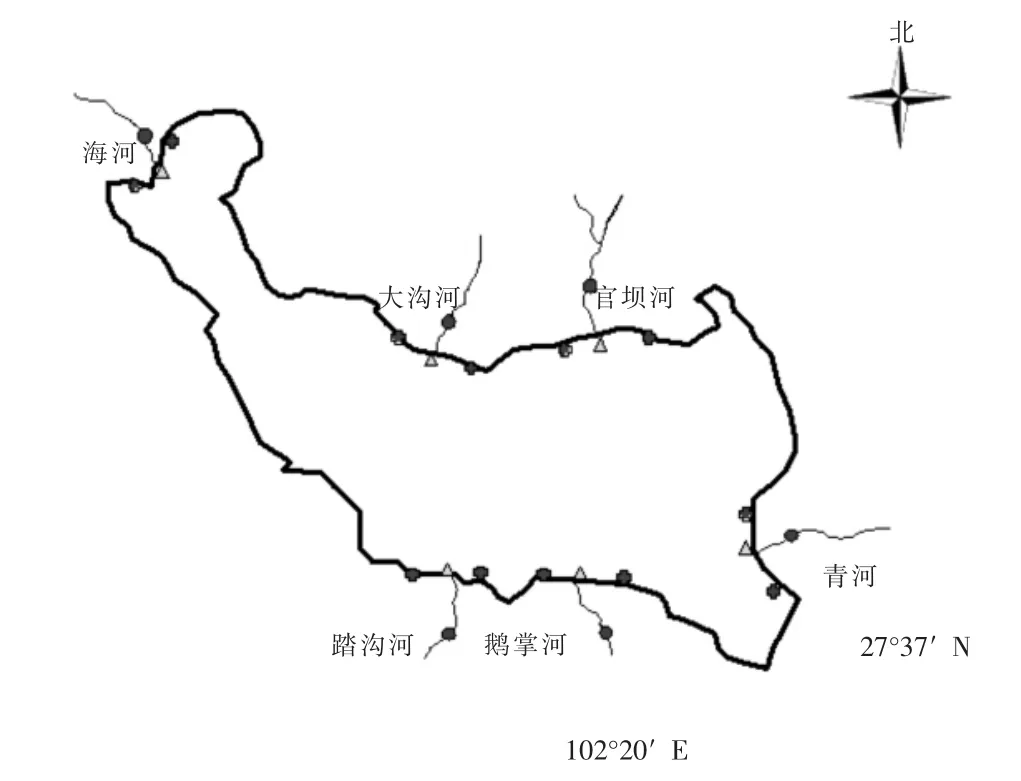
图1 邛海河-湖采样点布设
1.3 水样测定指标及方法
1.3.1 叶绿素 水体中叶绿素a(Chla)的浓度主要表征浮游植物的生物量,叶绿素b(Chlb)和叶绿素c(Chlc)的浓度可用于表征浮游植物组成。采用热乙醇-反复冻融法[5]提取叶绿素,利用分光光度计测定750、664、647、630 nm 波长处吸光度后计算各色素含量。
1.3.2 水体中碱性磷酸酶活性 水体中碱性磷酸酶可分为游离态(<0.22 μm)、微型浮游生物态(0.22~3 μm)、浮游植物态(>3 μm)3 种形态。 将水样经0.22 μm和3 μm醋酸纤维膜过滤划分为3种形态待测水样,采用对硝基苯磷酸二钠(PNPP)为底物,反应2 h后在410 nm波长处测其水解底物吸光度,计算碱性磷酸酶活性[6]。
1.3.3 水质因子 主要对与富营养化有关的水质因子进行测定,包括水中总氮(TN)、总磷(TP)和氨氮(NH4-N)浓度的测定。其中水中的总氮测定利用碱性过硫酸钾消解紫外分光光度法[7],总磷测定采用钼酸铵分光光度法[8],氨氮的测定采用纳氏试剂法[9]。
2 结果与分析
2.1 邛海河-湖系统浮游植物叶绿素浓度空间分布
邛海春季河-湖系统浮游植物叶绿素浓度空间分布特征如图2所示。从叶绿素a浓度来看,C1(青河邻近湖域)最高,为 6 μg/L,其次是 F3(海河河流)的5.9 μg/L、B3(踏沟河河流)的 5.8 μg/L、D2(大河沟河口)的 5.0 μg/L、D3(大河沟河流)的 4.7 μg/L,最少的是E2(官坝河河口),为 0.5 μg/L。 由此可见,青河、海河、踏沟河和大沟河的浮游植物生长较其他河流更快。
通过对叶绿素b/a和叶绿素c/a的计算可知,C2和C3两样点叶绿素b/a和叶绿素c/a显著高于其他样点(P<0.05),说明青河河口和邻近湖域水体中绿藻和其他藻类占优势。此外,不同河流间的其他叶绿素占比也存在差异,由此可知,不同河流藻类组成有所不同。
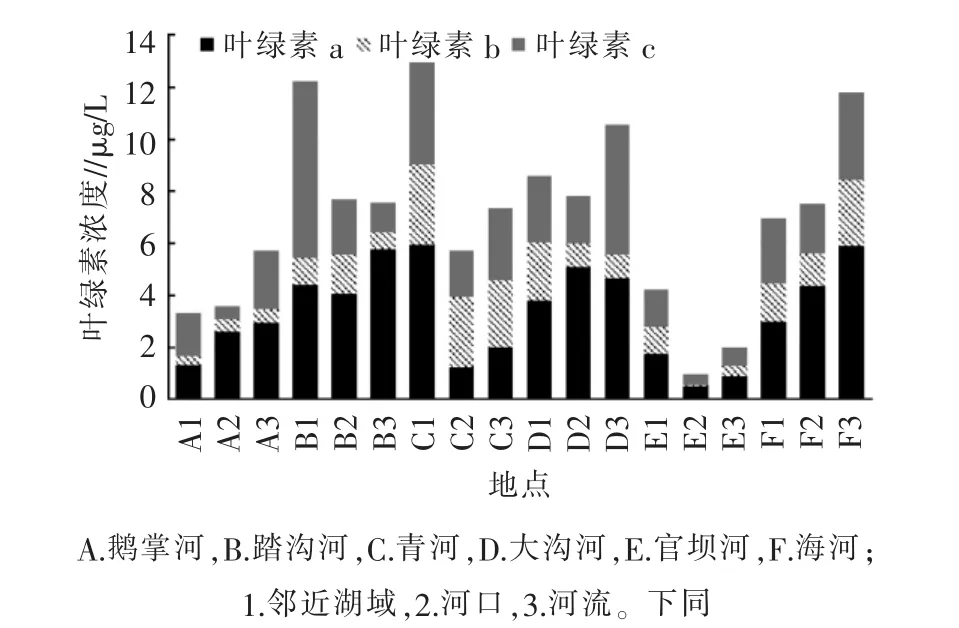
图2 邛海主要河流、河口和邻近湖域浮游植物叶绿素浓度
按河流不同位置区域进行比较可知,不同河流的叶绿素浓度变化趋势明显不同。鹅掌河从河流到邻近湖域叶绿素浓度逐渐下降,踏沟河叶绿素浓度从河流到河口下降,在邻近湖域又有所升高,但河流的叶绿素浓度最高;青河和官坝河的叶绿素浓度从河流到河口下降,但河口到邻近湖域急剧上升,邻近湖域>河流>河口;大河沟叶绿素浓度从河流到河口上升,但在邻近湖域浓度又下降且低于河流浓度;海河作为邛海的出水口,叶绿素浓度从河流到河口到临近湖域呈逐渐下降趋势。
综上可见,邛海春季河-湖系统浮游植物叶绿素浓度存在明显的空间异质性,可能是受不同河流的水文、水质影响[10]。
2.2 邛海河-湖系统碱性磷酸酶活性空间分布
邛海春季河-湖系统不同河流间海河的APA最高,其次是官坝河、青河,最低的是踏沟河,不同河流间的APA存在显著差异(P<0.05),其中海河和官坝河显著高于其他河流(P<0.05),其他河流间无显著差异(P>0.05)。
从同一河流不同位置的变化来看,鹅掌河APA从河流到河口有所增强,但变化很小,而从河口到邻近湖域急剧增加;踏沟河APA从河流到河口急剧上升,从河口到邻近湖域又急剧下降,河流和河口的APA无显著差异;青河APA从河流到河口增强,从河口到邻近湖域又急速下降,邻近湖域小于河流;大沟河APA从河流到河口急剧上升,从河口到邻近湖域又急剧减弱,且邻近湖域高于河流;官坝河APA从河流到邻近湖域逐步降低;海河从河流到河口到邻近湖域无显著差异(图3)。
从不同形态APA来看,总体上3种粒径对APA的贡献无显著差异(P>0.05),但不同样点有所不同,其中游离态(<0.22 μm)E2 的贡献最大,C2 的贡献最小;微型浮游生物态(0.22~0.30 μm)C3 的贡献最大,B1 的贡献最小;浮游生物态(>0.30 μm)B1 的贡献最大,A2的贡献最小。说明不同样点的碱性磷酸酶的主要来源存在差异。
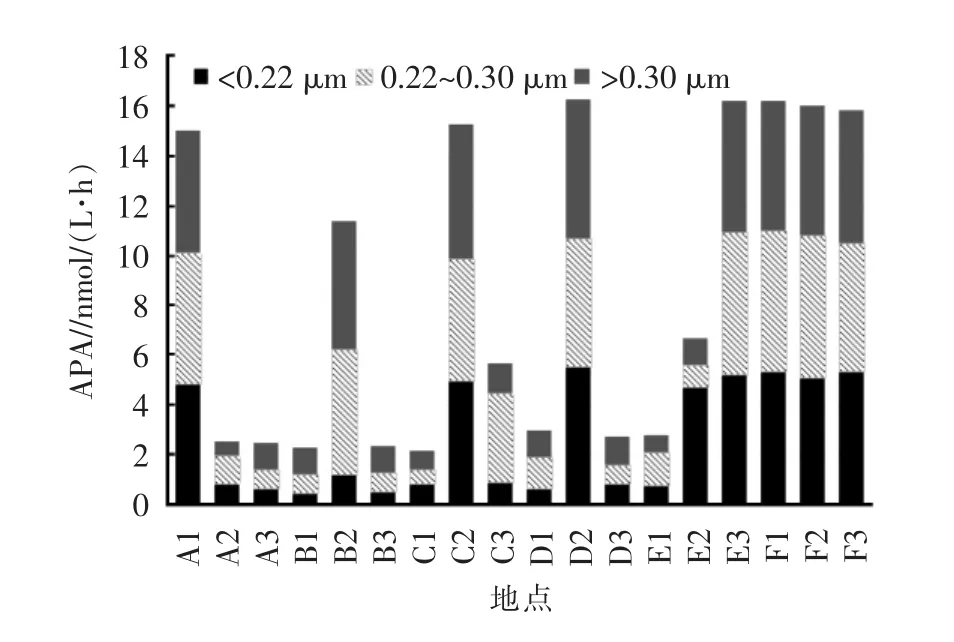
图3 邛海主要河流、河口和邻近湖域碱性磷酸酶活性
2.3 水质因子测定结果
邛海主要河流、河口和邻近湖域的水质因子测定结果见表1,因邛海应执行国家Ⅱ类水水质标准,可见6条河流从河流、河口到邻近湖域水温变化≤1℃,pH 6~9,水温和pH符合标准。鹅掌河河流、河口、邻近湖域总氮、总磷、氨氮含量均先减少后增加,河流(下游)总氮、总磷、最多,邻近湖域氨氮最多,河流总氮、氨氮含量超标,河口氨氮含量超标,邻近湖域总氮、氨氮含量超标。踏沟河河流、河口、邻近湖域总氮、总磷含量先减少后增加,氨氮含量先增加后减少,河流(下游)总磷、总氮含量最多,河口氨氮含量最多,河流总氮和氨氮含量超标,河口氨氮含量超标。青河河流、河口、邻近湖域总氮含量先增加后减少,总磷含量先减少后增加,氨氮含量逐渐增加,邻近湖域总磷、氨氮含量均最多,河口总氮含量最多,邻近湖域和河口的总氮和氨氮含量严重超标。大沟河河流、河口、邻近湖域总氮和氨氮含量逐渐减少,总磷含量逐渐增加,河流(下游)总氮、氨氮含量最多,邻近湖域总磷含量最多,河流、河口、邻近湖域的总氮含量和河流、河口的氨氮含量严重超标。官坝河河流、河口、邻近湖域总氮含量先增加后减少,氨氮含量先减少后增加,总磷含量逐渐减少,河流(下游)总磷含量最多,河口总氮含量最多,邻近湖域氨氮含量最多,河流、河口、邻近湖域的总氮和河流、邻近湖域的氨氮严重超标。海河河流、河口、邻近湖域总氮和总磷含量逐渐减少,氨氮含量先减少后增加,河流(上游)总氮、总磷和氨氮含量最多,河流、河口、邻近湖域总氮含量和河流、邻近湖域氨氮含量严重超标。
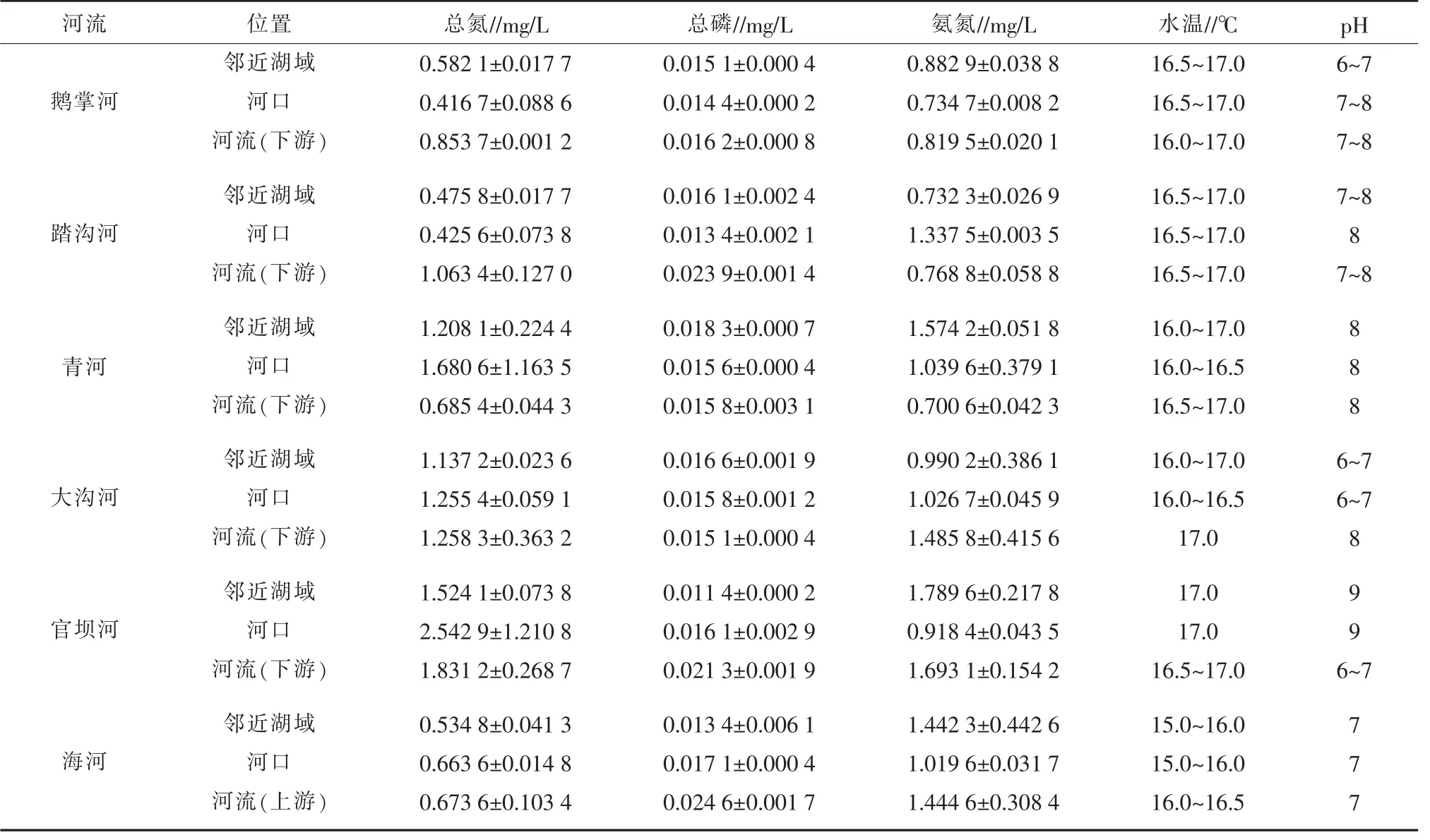
表1 邛海主要河流、河口和邻近湖域水质参数
2.4 邛海河-湖系统浮游植物磷胁迫
通过将总APA归一化为单位叶绿素活性(APAChla)进行对比分析,以消除生物量的影响。研究表明,APAChla可以作为水体磷是否缺乏状态的一种良好指示参数。由图4可见,E3(官坝河河流)APAchla最高,其次是 E2(官坝河河口)、C2(青河河口)、A1(鹅掌河邻近湖域),而C1(青河邻近湖域)APAchla最低。表明官坝河的河流受磷胁迫的程度最高,而官坝河附近主要为农田和居住区,结合水质测定结果中官坝河的总氮和氨氮含量严重超标,分析原因可能是由于该区域居住人口多,农业活动发达,人类生活和生产中大量废水排入河流导致营养盐的大量输入,但由于主要污水为含氮污水,使得氮磷比升高,而磷成为限制因子。青河邻近湖域受磷胁迫的程度最小,但其河口受磷胁迫的程度较其他位置高,主要因为青河河口为旅游区,商业活动发达,导致其污水集中排放,从水质指标可见该处总氮和氨氮含量超标,氮磷比增大导致该区域浮游植物磷胁迫,而青河邻近湖域是邛海水体透明度最高的区域,因此到邻近湖域由于水体扩散稀释作用及光照条件的改善使得浮游植物快速生长,解除了磷胁迫。踏沟河和大沟河在河口位置的APAchla也较河流和邻近湖域高,可能也是由于河口人类活动较频繁,污水排放影响了氮磷比。鹅掌河的APAChla最大值出现在邻近湖域,可能因为该区域底质主要为泥沙,河流水体到湖域后流速降低,水面变宽,大量泥沙携带营养盐沉淀,导致该区域的浮游植物磷胁迫。海河作为邛海的出海河流,从邻近湖域、河口河流(上游)APAChla不断降低,主要因为海河周边商业发达,大量的污水排入,严重时还会出现污水倒灌的情况,所以邻近湖域的磷胁迫程度更高。
此外,研究表明,高APA尤其是高比例的游离态APA(粒级<0.22 μm)能反映过去一段时间浮游植物群落的磷胁迫或限制状态[11]。本研究中河流和邻近湖域的游离态APA与叶绿素a呈正相关(P<0.01),表明游离态的APA主要是由浮游植物产生,河流和邻近湖域浮游植物长时间受磷胁迫;而河口的游离态 APA 与叶绿素呈负相关(r=-0.65,P>0.05),不能由此判断其磷胁迫状态。
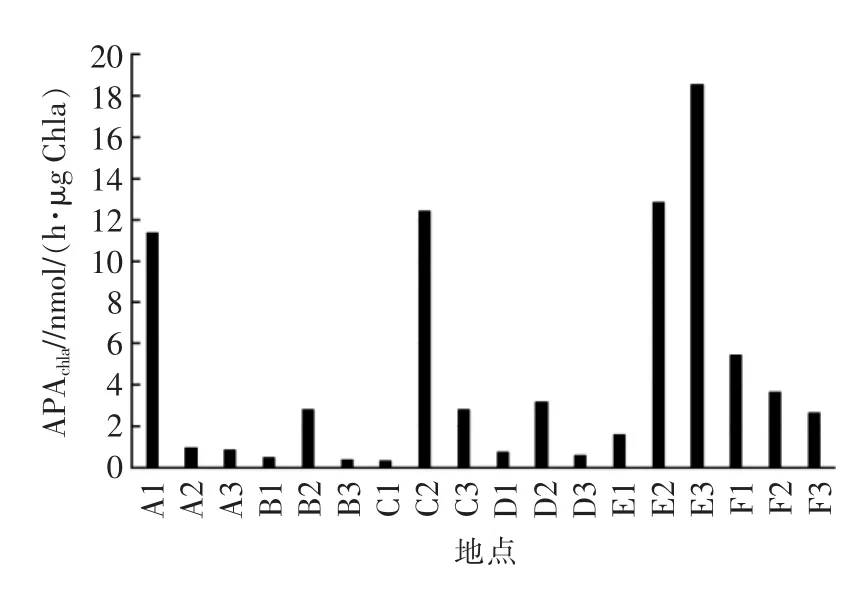
图4 邛海主要河流、河口和邻近湖域单位叶绿素碱性磷酸酶活性
2.5 水质因子对碱性磷酸酶活性的影响
对邛海河-海系统不同区域单位叶绿素碱性磷酸酶活性与水质因子的相关性进行分析,结果(表2)表明,APAchla与总氮、总磷、氨氮含量及氮磷比在邻近湖域均呈负相关,在河流均呈正相关,而在河口APAchla与氨氮含量呈负相关,与其他3个水质因子呈正相关。研究表明,磷胁迫不仅取决于磷含量,还取决于浮游植物对磷的需求[12],邻近湖域APAchla与氮磷比呈负相关,说明氮的输入相对于磷过量时,也会导致磷的相对缺乏,过高的氮磷比也会导致潜在的磷胁迫与限制;而在河口APAchla与氮磷比呈显著正相关,说明碱性磷酸酶在一定范围内受氮磷比的控制;河流中APAchla与氮磷比虽然呈正相关,但不显著,说明该区域还有其他影响因子起作用。
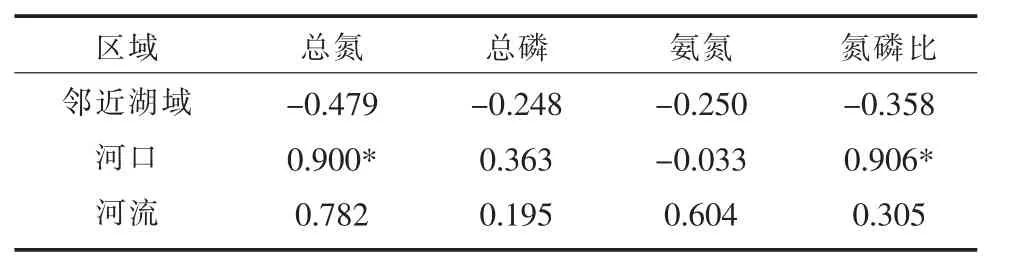
表2 邛海河-海系统不同区域单位叶绿素碱性磷酸酶活性与水质因子的皮尔逊相关系数
对邛海主要河流单位叶绿素碱性磷酸酶活性与水质因子的相关性分析见表3。鹅掌河APAchla与氨氮、总磷含量呈正相关,与氮磷比、总氮含量呈负相关,其中与氨氮含量相关性最强;踏沟河除了氨氮以外,其他环境因子都与 APAchla呈负相关,其中APAchla与氨氮含量的相关性最强,与氮磷比的相关性最弱;青河APAchla与总氮含量、氮磷比呈正相关,与其他两个环境因子呈负相关;大沟河APAchla与总氮、总磷含量及氮磷比均呈正相关,与氨氮含量呈负相关,其中与氨氮含量的相关性最强,与总磷含量的相关性最弱;官坝河APAchla与总氮、总磷含量呈正相关,与其他两个因子呈负相关;海河APAchla与总氮、总磷含量呈负相关,与其他两个因子呈正相关。但所有河流APAchla与水质因子的相关性都不显著,因为各河流除水质因子外还有其他因子对浮游植物影响较大,不同河流由于周边人类活动情况不同所以其相关性也不同。因此不同河流在控制富营养化时应采取不同的措施,可参考河流水质因子与单位叶绿素APA的相关性选择水质的控制指标。
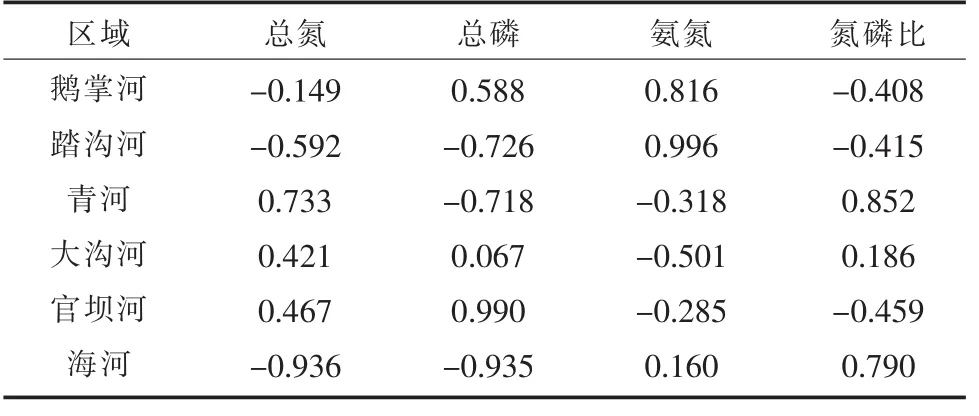
表3 邛海主要河流单位叶绿素碱性磷酸酶活性与水质因子的皮尔逊相关系数
3 结论
通过对邛海河-湖系统春季浮游植物叶绿素浓度空间分布、碱性磷酸酶活性及水质因子的分析得到以下结论。
1)邛海春季河-湖系统浮游植物叶绿素浓度和碱性磷酸酶活性存在明显的空间异质性,进而导致不同河流和不同区域存在不同程度的磷胁迫。官坝河河流的胁迫程度最高,其次是官坝河河口、青河河口、鹅掌河邻近湖域,而青河邻近湖域的胁迫程度最低。
2)邛海春季河-湖系统中河流和邻近湖域的游离态碱性磷酸酶与叶绿素a含量呈正相关(P<0.01),河流和邻近湖域浮游植物长时间受磷胁迫,而河口的游离态碱性磷酸酶与叶绿素呈负相关 (r=-0.65,P>0.05),不能由此判断其磷胁迫状态。
3)邛海春季河-湖系统中邻近湖域的单位叶绿素碱性磷酸酶与氮磷比呈负相关,说明氮的输入相对于磷过量时,也会导致磷的相对缺乏,过高的氮磷比也会导致潜在的磷胁迫与限制;而在河口单位叶绿素碱性磷酸酶与氮磷比呈显著正相关,说明碱性磷酸酶在一定范围内受氮磷比的控制;河流单位叶绿素碱性磷酸酶与氮磷比虽然呈正相关,但不显著,说明该区域还有其他影响因子起作用。
4)邛海春季河-湖系统不同河流水质因子与单位叶绿素碱性磷酸酶存在相关性但不显著,说明各河流除水质因子外还有其他因子对浮游植物影响较大。因此不同河流在控制富营养化时应采取不同的措施,可参考河流水质因子与单位叶绿素碱性磷酸酶的相关性选择水质的控制指标。
猜你喜欢
东坡赤壁诗词(2020年3期)2020-07-04
少年文艺·开心阅读作文(2018年7期)2018-08-12
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
广西林业科学(2016年1期)2016-03-20
医学研究杂志(2015年8期)2015-06-22
岷峨诗稿(2014年4期)2014-11-15
中国医学科学院学报(2013年6期)2013-03-11
