
辅助发酵菌株对发酵乳品质的影响
2019-07-24 08:01罗天淇李晓琬王世杰杨贞耐
乳业科学与技术 2019年3期
罗天淇,蔡 淼,张 敏,李晓琬,王世杰,朱 宏,杨贞耐,*
(1.北京工商大学,北京食品营养与人类健康高精尖创新中心,北京市食品添加剂工程技术研究中心,北京 100048;2.石家庄君乐宝乳业有限公司,河北 石家庄 050221)
酸乳是通过保加利亚乳杆菌和嗜热链球菌共同发酵牛乳生产的一种发酵乳制品,凭借其高营养价值和独特风味,深受广大消费者的喜爱[1-2]。近些年来,随着生活水平的提高,人们对于食品的营养和功能追求也越来越高[3],为了改善酸乳的风味、质地、外观等品质特性,增强酸乳的益生性,在常规酸乳发酵剂的基础上添加其他乳酸菌进行发酵已经成为一种流行的做法。研究表明,不同菌株的加入能够对发酵乳品质产生不同的影响。瑞士乳杆菌H9可以加快发酵乳的酸化和蛋白质水解,促进乙偶姻和苯甲醛的形成[4]。鼠李糖乳杆菌GG能够显著降低山羊乳酸乳中短链和中链脂肪酸的含量[5]。植物乳杆菌的加入能够引起发酵乳质地(黏附性)和挥发性风味化合物组成(乙醛)的明显变化[6]。产黏乳杆菌与传统酸乳发酵剂共同发酵会延长酸乳的发酵时间,提高酸乳的黏度与保水性,改善酸乳口感[7]。
本研究使用7 株从传统发酵乳制品中分离的常用乳酸菌作为辅助菌株,与常规酸乳发酵剂共同发酵,系统地研究比较这些菌株对发酵乳品质的影响,为进一步了解辅助发酵菌种的作用机制及功能性发酵乳产品研发提供理论依据。
1 材料与方法
1.1 材料与试剂
全脂乳粉 新西兰恒天然集团;发酵剂YO-MIX 300 LYO(保加利亚乳杆菌和嗜热链球菌) 丹麦Danisco公司;辅助菌株:包括乳酸乳球菌乳脂亚种(Lactococcus lactissubsp.cremoris,Lcr)、乳酸乳球菌乳酸亚种(Lactococcus lactissubsp.lactis,Ll)、干酪乳杆菌(Lactobacillus casei,Lca)、发酵乳杆菌(Lactobacillus fermentum,Lf)、鼠李糖乳杆菌(Lactobacillus rhamnosus,Lr)和2 株植物乳杆菌(Lactobacillus plantarum,Lp1、Lp2) 本实验室分离保藏。
1.2 仪器与设备
MLS-3781L-PC高压蒸汽灭菌锅 日本Sanyo公司;DHP-9032电热恒温培养箱 上海一恒科学仪器有限公司;iCinac乳品发酵监控仪 法国AMS Alliance公司;Rheolaser Master光学微流变仪 法国Formulaction公司;7890A-7000气相色谱-质谱联用(gas chromatography-mass spectrometry,GC-MS)仪美国Agilent公司;CR 21G Ⅲ高速冷冻离心机 日本Hitachi公司;Brookfield CT3质构仪 美国Brookfield公司;FE28数显pH计 Mettler-Toledo仪器(上海)有限公司。
1.3 方法
1.3.1 发酵乳的制备
使用全脂乳粉制备质量浓度12 g/100 mL的全脂复原乳,65 ℃条件下杀菌30 min,冷却至室温,接菌后37 ℃发酵至pH值为4.5,快速冷却并搅拌后置于4 ℃贮藏。其中,对照组(D)只加入商品发酵剂YO-MIX 300 LYO,实验组(D-Lr、D-Lf、D-Lca、D-Lp1、D-Lp2、D-Lcr、D-Ll)添加辅助菌株和商品发酵剂(1∶1),接菌量均为107CFU/mL。
1.3.2 发酵乳pH值、氧化还原电位的测定
采用iCinac乳品发酵监控仪对发酵过程中pH值、氧化还原电位的变化进行实时监测,每1 min记录1 次。采用pH计记录发酵乳贮藏期间的pH值变化。
1.3.3 发酵乳中活菌数的测定
参照GB 4789.35—2016《食品安全国家标准 食品微生物学检验 乳酸菌检验》测定发酵乳中乳酸菌的总菌数。
1.3.4 发酵乳微流变学参数测定
取接菌后的牛乳20 mL至微流变仪专用样品池中,将样品池插入微流变仪,运行实时测试程序。在37 ℃条件下实时监测发酵凝胶过程中的宏观黏度指数(micro viscosity index,MVI)、弹性指数(elasticity index,EI)及流动性指数(fluidity index,FI)随时间的变化情况,直至发酵乳pH值降至4.5,每隔3 min采集1 次数据。然后将样品放置于4 ℃冷藏21 d,定期取样,检测发酵乳微流变学参数的变化。
1.3.5 发酵乳持水力测定与质构分析
各组均准确称量10 g发酵乳样品,分别放于标号的离心管中并称质量,放入离心机中4 000 r/min离心20 min,去上清液,称质量。持水力按照下式计算。
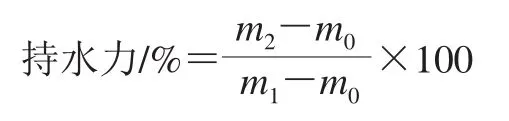
式中:m0为空离心管质量/g;m1为离心管加样品的质量/g;m2为样品离心除去上清液后的离心管加样品质量/g。
参考庞志花等[8]的方法,使用Brookfield CT3质构仪进行质地剖面分析(texture profile analysis,TPA),选用圆柱形TA10探头,目标值10.0 mm,测试速率0.5 mm/s。测定参数包括硬度、内聚性和胶着性。
1.3.6 发酵乳挥发性风味物质测定
固相微萃取(solid phase micro-extraction,SPME)法提取挥发性风味物质:萃取前将SPME纤维置于GC-MS进样口老化。取10 g发酵乳样品和1 μL内标物2-甲基-3-庚酮于30 mL萃取瓶中,内标物质量浓度为0.816 μg/μL,加盖密封,在40 ℃恒温水浴中平衡30 min后,将SPME萃取纤维通过瓶盖插入样品中的顶空部分,推出纤维,顶空吸附30 min后拔出;快速插入GC-MS进样口解吸5 min,进行GC-MS分析。
GC条件:采用DB-wax色谱柱(30 m×0.25 mm,0.25 μm)进行分析。升温程序:起始柱温40 ℃,保持3 min,以5 ℃/min升温至200 ℃,以10 ℃/min升至230 ℃,保持3 min,载气为氦气,流速1.2 mL/min,不分流进样。
MS条件:电子电离源,电子能量70 eV,传输线温度280 ℃,离子源温度230 ℃,四极杆温度150 ℃,质量扫描范围40~250(m/z)。
定性分析:化合物通过NIST 14质谱数据库和文献报道的保留指数(retention index,RI)来鉴定。
定量分析:使用2-甲基-3庚酮为内标物,采用内标法进行定量分析,求得各挥发性成分的含量。
1.4 数据处理
每个样品至少测定3 次,实验数据采用SPSS 17.0软件进行分析,P<0.05表示差异显著,使用Sigma Plot软件作图,风味物质用NIST 14谱库检索,并应用Canoco 4.5统计分析软件对风味数据进行主成分分析。
2 结果与分析
2.1 发酵乳pH值、氧化还原电位与活菌数变化
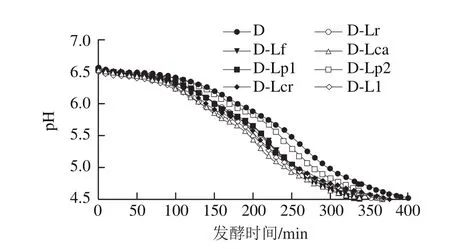
图1 发酵过程中发酵乳pH值的变化Fig. 1 Changes in pH value of fermented milk during fermentation
由图1可知,各组发酵乳的发酵pH值曲线变化相同,呈现先下降后趋于平稳的趋势,但到达发酵终点(pH=4.5)的时间存在一定差异,分别为407(D)、368(D-Lr)、351(D-Lf)、337(D-Lca)、340(D-Lp1)、376(D-Lp2)、386(D-Lcr)、376 min(D-Ll),说明辅助菌株的加入能够不同程度地缩短发酵时间,如Lca、Lp1能显著加快牛乳发酵,而Lcr则影响较小,这一差异可能与辅助菌株和常规酸乳发酵剂之间的相互作用有关。
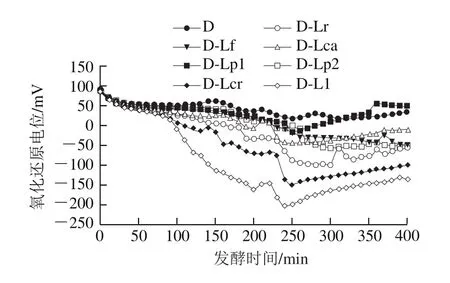
图2 发酵过程中发酵乳氧化还原电位的变化Fig. 2 Changes in oxidation-reduction potential of fermented milk during fermentation
氧化还原电位是乳酸菌的基本物理化学性质之一,氧化性强,氧化还原电位值较高;还原性强,氧化还原电位值较低[9]。由图2可知,各组发酵乳的氧化还原电位均呈先下降后缓慢提升的趋势,其中D、D-Lp1组变化最小,而添加乳酸乳球菌的D-Ll、D-Lcr组发酵乳氧化还原电位变化最大。Morandi等[10]研究709 株乳酸菌的还原活性后指出,粪肠球菌和乳酸乳球菌是还原活性最高的菌种,而嗜热链球菌的还原活性最低,与本研究结果基本吻合。
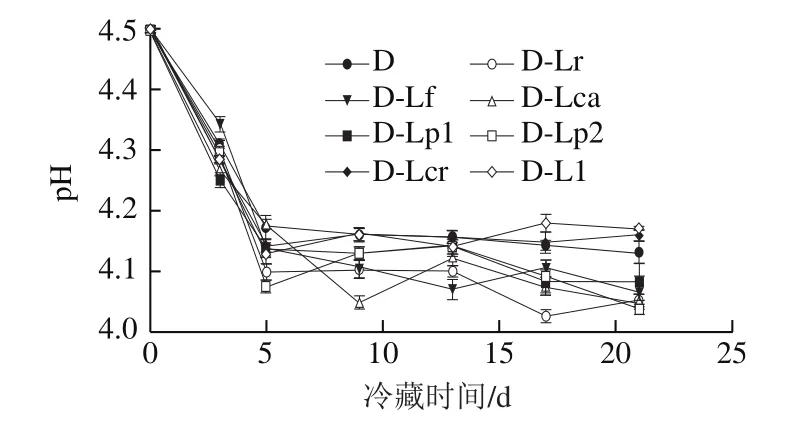
图3 4 ℃冷藏过程中发酵乳pH值的变化Fig. 3 Changes in pH value of fermented milk during 4 ℃ storage
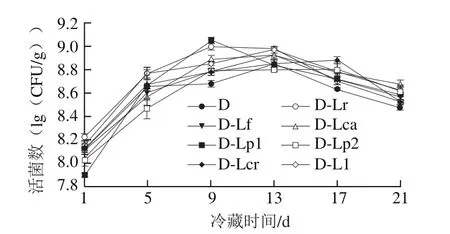
图4 4 ℃冷藏过程中发酵乳活菌数的变化Fig. 4 Changes in viable cell count of fermented milk during 4 ℃ storage
由图3~4可知,整体上冷藏期间发酵乳的pH值呈现出先快速下降后趋于稳定的趋势,但各组样品pH值存在一定差异,其中D-Ll和D-Lcr组pH值高于对照组,而D-Lp1、D-Lp2、D-Lca、D-Lf、D-Lr组pH值明显较低,说明这些辅助菌株的加入会影响发酵乳的后酸化过程。而各组发酵乳的活菌数在冷藏期间先升高后降低,同时对照组中的活菌数相对较低。
2.2 发酵乳发酵过程与冷藏期间的微流变学变化
微流变仪能够通过监测样品中粒子的运动,在不破坏样品的前提下研究样品流变特性[11]。由图5~10可知,各组发酵乳的EI、MVI和FI变化趋势相同,但是各组发酵乳的这些流变学特性之间存在一定差异。
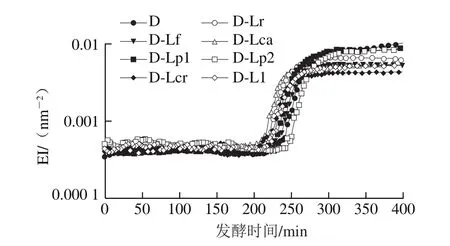
图5 发酵乳发酵过程中EI的变化Fig. 5 Changes in EI of fermented milk during fermentation
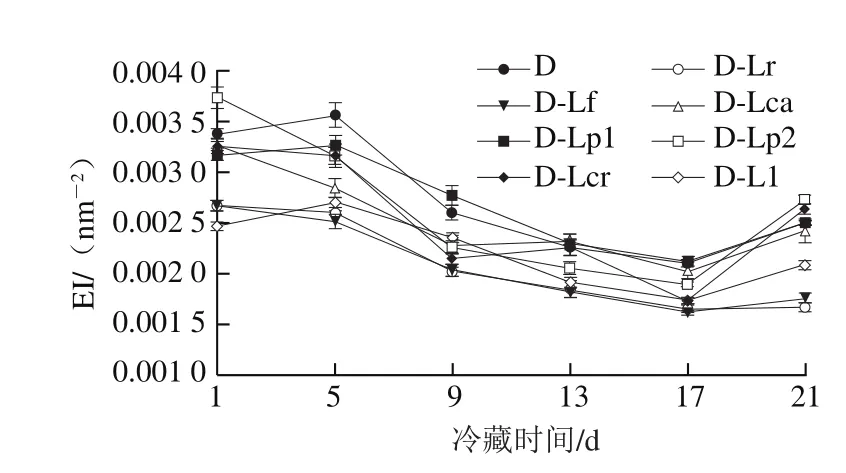
图6 4 ℃冷藏过程中发酵乳EI的变化Fig. 6 Changes in EI of fermented milk during 4 ℃ storage
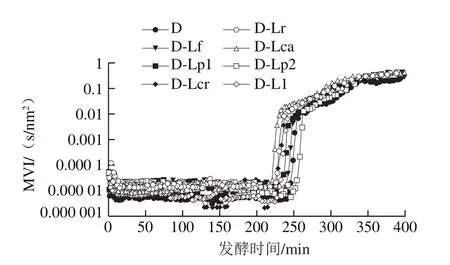
图7 发酵乳发酵过程中MVI的变化Fig. 7 Changes in MVI of fermented milk during fermentation
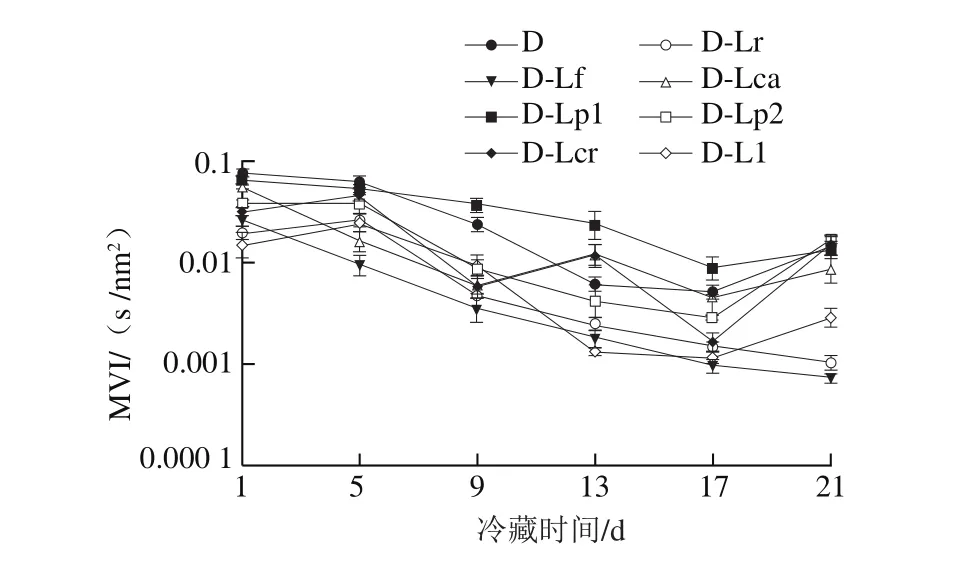
图8 4 ℃冷藏过程中发酵乳MVI的变化Fig. 8 Changes in MVI of fermented milk during 4 ℃ storage
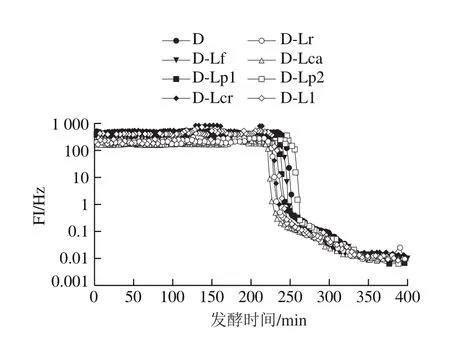
图9 发酵乳发酵过程中FI的变化Fig. 9 Changes in FI of fermented milk during fermentation
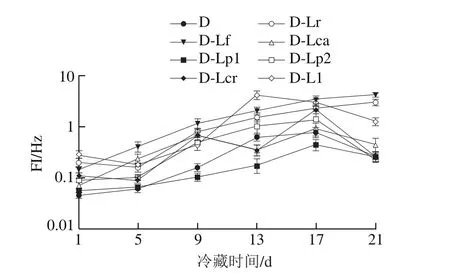
图10 4 ℃冷藏过程中发酵乳FI的变化Fig. 10 Changes in FI of fermented milk during 4 ℃ storage
凝胶EI、MVI、FI的变化可以分别表征发酵乳的弹性、黏性及流动性,EI、MVI越大,样品的黏弹性越强,而FI越大则样品流动性越强。结合图1的发酵pH值曲线可知,在发酵过程中(图5、7、9),发酵乳的EI、MVI、FI变化可以分为3 个阶段。在发酵前期,发酵乳pH值由起始值降至5.3左右,样品处于相对稳定的低黏度、低弹性、高流动性状态;当pH值进入5.3~5.0这一范围内时,发酵乳黏度、弹性快速增加,而流动性快速下降,这与酪蛋白引起的凝胶化有关[12];之后,发酵乳初步形成较为稳定的凝胶结构,黏性、弹性、流动性变化减缓,进入高黏弹性、低流动性阶段。发酵结束时,与其他组相比,对照组的MVI最低,EI和FI最高。4 ℃贮藏时(图6、8、10),由于温度的降低及搅拌的影响,发酵乳的MVI、EI在第1天出现一定程度的降低,FI升高。随后,各组发酵乳的MVI、EI逐渐降低,FI逐渐升高,流动性不断增强,这可能与后酸化引起的凝胶结构不稳定有关。其中,D-Lp1组发酵乳的MVI、EI最高,FI最低,具有良好的稳定性,结合实验室前期实验结果,推测可能与Lp1高产胞外多糖、促进凝胶结构稳定的特性有关[13]。
2.3 发酵乳冷藏期间持水力及质构特性变化

表1 发酵乳4 ℃冷藏期间的持水力和质构特性变化Table 1 Changes in water-holding capacity and texture of fermented milk during 4 ℃ storage
持水力是发酵乳品质高低的重要指标之一,它对延长发酵乳产品的货架期起着关键性作用。由表1可知,冷藏期间各组发酵乳的持水力整体上呈先升高后下降的趋势,同时实验所用辅助菌株的加入均能在一定程度上提高发酵乳的持水性能,有利于发酵乳凝胶结构的稳定性。
TPA可以模拟人类咀嚼食品,目前被广泛应用于固体、半固体食品的质构分析中[14]。内聚性能够反映样品内部分子间或各结构要素间结合作用的强弱,内聚性越大,结合作用越强[15],而胶着性反映使半固态样品破裂成吞咽时的分散状态所需的能量。TPA结果表明,Lf能够有效提高发酵乳的硬度,Lr、Lf、Lp1对发酵乳的内聚性、胶着性有增强作用。因此Lr、Lf、Lp1这3 株辅助菌株对发酵乳结构的稳定性具有积极作用。
2.4 发酵乳的风味物质分析
2.4.1 SPME-GC-MS检测结果
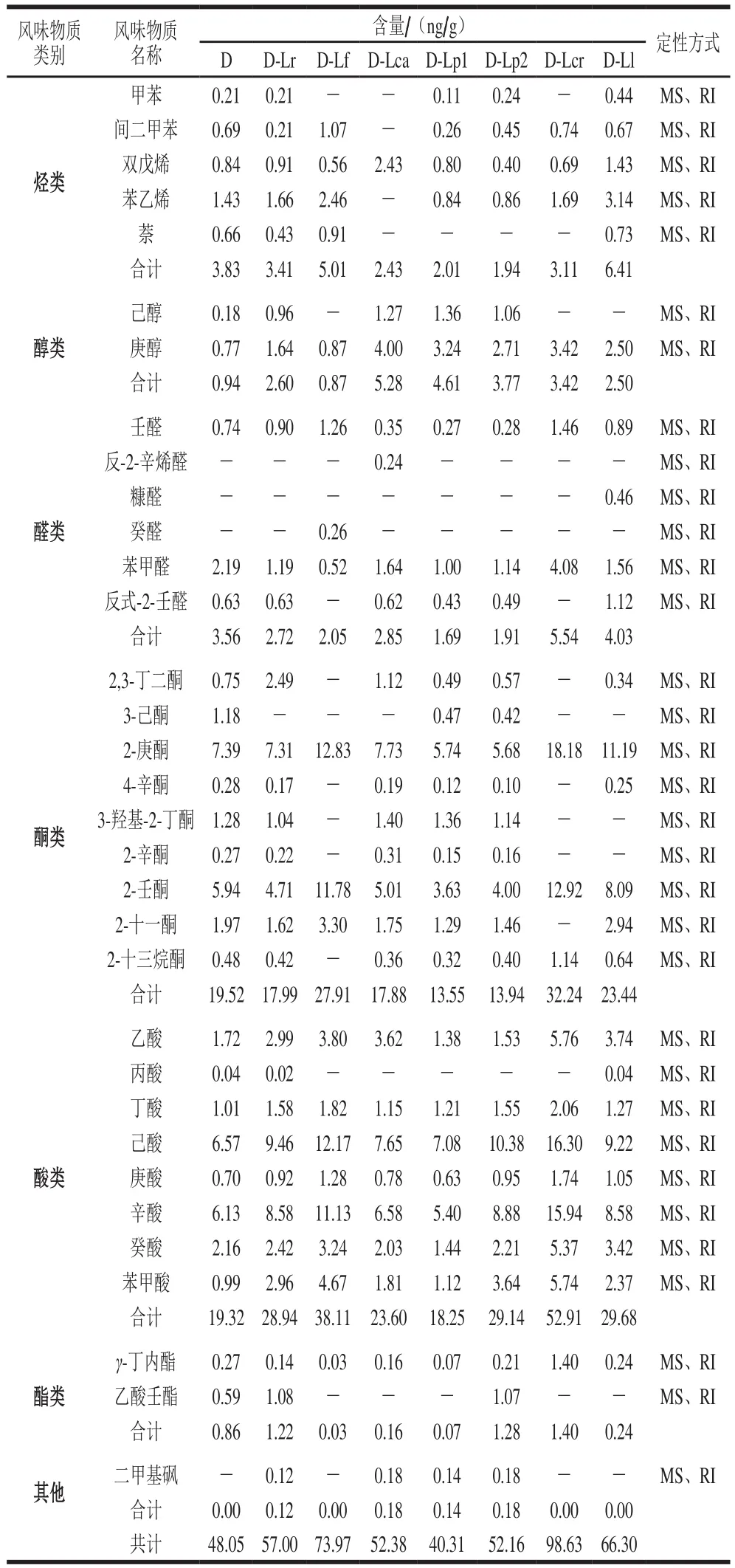
表2 发酵乳挥发性组分的定性及定量分析结果Table 2 Qualitative and quantitative analysis results of volatile compounds in fermented milk
由表2可知,经过SPME萃取,GC-MS检测分析后,从8 组酸乳样品中共检出挥发性物质33 种,包括烃类5 种、醇类2 种、醛类6 种、酮类9 种、酯类2 种、酸类8 种、其他化合物1 种。由于8 组样品发酵使用菌株的不同,造成发酵乳中挥发性风味物质组成、含量上的差异,形成不同的风味。
在风味研究中,烯烃和芳香烃类物质阈值较低,且具有特殊香气,对产品风味贡献较大;而烷烃类物质阈值较高,通常对样品风味的贡献不大。在检测出的5 种烃类物质中,甲苯具有水果甜味,苯乙烯能赋予食品甜味和花香味[16],萘具有类似樟脑丸的气味。通常认为在发酵乳制品中,烃类物质大多来源于原料乳自身,但由表2可知,加入不同发酵菌株后各组发酵乳样品的烃类物质含量存在显著差异(P<0.05),说明发酵乳中的烃类物质受发酵菌株的影响。
在8 组发酵乳样品中仅检出己醇和庚醇2 种醇类挥发性物质,且含量较低,其来源可能与甲基酮类物质的还原有关[17]。其中,己醇具有树脂和花的香味,在常规发酵剂发酵的D组酸乳中含量较低,而在分别添加发酵乳杆菌、乳酸乳球菌的D-Lf、D-Lcr、D-Ll 3 组样品中没有检出。同时郝晓娜等[18]发现,在乳酸乳球菌发酵制作干酪的过程中添加植物乳杆菌,可以额外产生己醇等风味物质,说明己醇在发酵乳中的产生可能具有菌株特异性。
醛类挥发性物质是发酵乳风味的重要组成部分,通常其风味阈值较低,但由于醛类化合物化学性质比较活泼,易发生还原反应,因此这类化合物在发酵乳制品中的含量通常都不高,在本研究8 组酸乳样品中的含量为2%~8%。在这些醛类物质中,壬醛带有鲜花、柑橘和脂肪的香气,但是含量超过某个阈值时会产生令人不愉快的气味[19];苯甲醛具有杏仁香,可以通过色氨酸降解生成;反式-2-壬醛有类似黄瓜、脂肪的香气。此外,反-2-辛烯醛只在D-Lca组检出,糠醛只在D-Ll组检出,癸醛只在D-Lf组检出,它们分别能赋予发酵乳坚果、鲜花、橙皮与牛油气味,这些特异性物质的存在可能能够参与益生菌发酵乳特征风味的形成。
酮类化合物是发酵乳中一类重要的风味物质,在各组样品中的含量为26%~40%。通常认为发酵乳中的酮类物质可以通过脂肪酸氧化、热降解、氨基酸降解和微生物代谢等多种途径形成,并且随着碳链的增长香气更加浓郁。在这些酮类物质中,2,3-丁二酮(双乙酰)、3-羟基-2-丁酮(乙偶姻)是乳酸菌代谢乳糖的产物,同时也是酸乳中极其重要的风味物质。双乙酰具有浓郁的奶油香味,相关研究表明,当酸乳中乙醛含量较低时,双乙酰能赋予酸乳独特的风味[20]。而乙偶姻是双乙酰的还原形式,和双乙酰一样有奶油香味,但香味远弱于双乙酰[21]。D-Lf、D-Lcr组中没有双乙酰检出,D-Lf、D-Lcr、D-Ll组中没有乙偶姻检出,说明这3 株菌对双乙酰、乙偶姻的生成存在抑制作用,可能与菌株间对相关底物的竞争作用及代谢通路有关。此外,2-庚酮、2-辛酮、2-壬酮、2-十一酮及2-十三烷酮等甲基酮来源于饱和脂肪酸的氧化水解[22],能够赋予发酵乳浓郁的香气,3-己酮具有葡萄香气。
酸类挥发性物质是发酵乳风味物质中的主要组成部分,在8 组样品中其含量分别为40.21%、50.77%、51.52%、45.07%、45.27%、55.86%、53.65%和44.77%,同时除了Lca、Lp1以外,其他5 株菌的加入均能有效提高发酵乳中的挥发性脂肪酸含量,并且添加乳酸乳球菌乳脂亚种的D-Lcr组酸乳的酸类物质含量显著高于其他组(P<0.05),与李良[23]关于乳酸乳球菌LL单独发酵可以提高稀奶油中脂肪酸和甲基酮含量的研究结果相似。通常认为,在发酵乳制品中酸类物质对整体风味的贡献主要表现在滋味上,但对气味也有一定影响。潘国卿等[24]认为,发酵乳制品中的短链脂肪酸是发酵乳奶油香味的主要组成成分。这些酸类物质中,乙酸具有醋酸味,丁酸、庚酸具有奶油味,己酸、癸酸具有油脂味,辛酸有微弱的水果酸气味。
酯类主要是通过脂肪酸水解、微生物代谢2 种途径产生,γ-丁内酯具有牛乳、奶油气味[25],乙酸壬酯具有香甜的水果气味。而二甲基砜具有硫磺气味,可能与原料乳杀菌时的美拉德反应及氨基酸降解有关。2.4.2 发酵乳风味物质组成的主成分分析
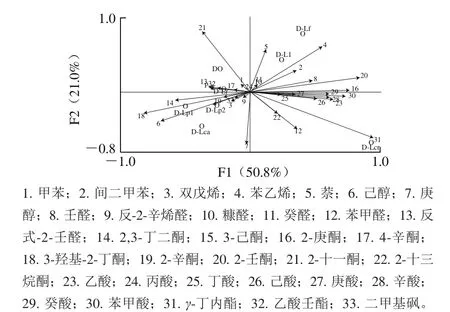
图11 发酵乳风味物质组成的主成分分析Fig. 11 Principal component analysis of fermented milk flavor compounds
为明确不同菌株与常规酸乳发酵剂复配对发酵乳风味的影响,利用主成分分析法考察挥发性风味物质与样品间的分布关系。由图11可知,F1和F2的贡献率分别为50.8%、21.0%,可以很好地解释不同菌株组合对酸乳风味的影响。各组样品在主成分分析图中的分布比较分散,说明不同菌株的加入使得常规发酵剂发酵酸乳的挥发性风味物质组成发生明显变化。
D-Lr、D-Lp1和D-Lp2 3 组发酵乳与D组发酵乳距离较近,仅在F2上存在一定差异,说明其对发酵乳风味的影响较小,可能与鼠李糖乳杆菌及植物乳杆菌在牛乳中生长较慢、在发酵过程中对发酵乳风味的贡献有限有关[26],造成这几组发酵乳风味差异的主要风味物质包括己醇(6)、2,3-丁二酮(14)和3-羟基-2-丁酮(18)。在这3 种物质的基础上,庚醇(7)、苯甲醛(12)和2-十三烷酮(22)引起D、D-Lca 2 组间的风味差异。而D-Lf、D-Ll组与D组在F1上存在显著差异,D-Lcr组与D组更是在F1、F2上均存在明显差异,同时乙酸(23)等酸类物质对这些差异的产生有较大贡献,说明Lf、Ll、Lcr这3 株菌与常规酸乳发酵剂共同发酵主要通过促进脂肪酸类挥发性物质的形成影响发酵乳的整体风味,并且对牛乳发酵过程中风味的形成有较大影响。
3 结 论
通过使用7 株不同的乳酸菌与常规酸乳发酵剂共同发酵,研究这些菌株辅助发酵时对发酵乳品质的影响。结果表明:辅助菌株的加入能够在不同程度上加快发酵过程中的产酸速率,与对照组相比可以缩短5.15%~17.20%的发酵时间,乳酸乳球菌Lcr、Ll在发酵过程中还会引起氧化还原电位的显著变化,同时发酵乳在冷藏期间的pH值也会受到影响;微流变学研究结果显示,在发酵过程中辅助菌株能够提高发酵乳的黏性,并降低弹性和流动性,而植物乳杆菌Lp1能够使发酵乳在贮藏期间具有高黏弹性及低流动性;这些菌株均能提高发酵乳的持水力,Lr、Lf、Lp1对发酵乳的内聚性、胶着性有增强作用;GC-MS和主成分分析结果显示,Lr、Lp1、Lp2对发酵乳风味影响较小,而Lca能够增加发酵乳中庚醇、苯甲醛、2-十三烷酮的含量,Lf、Ll、Lcr的加入会促进发酵乳中短链、中链脂肪酸等风味物质的形成。本研究结果揭示了不同辅助发酵菌株对酸乳发酵特性、微流变、质构和风味的影响,结果为高品质功能性发酵乳研发提供了参考。
猜你喜欢
食品科学(2022年12期)2022-07-07
中国土壤与肥料(2021年5期)2021-12-02
保鲜与加工(2021年1期)2021-02-06
陕西科技大学学报(2020年2期)2020-05-13
乳业科学与技术(2019年4期)2019-12-23
食品界(2018年10期)2018-11-23
肉类研究(2016年4期)2016-05-17
专用汽车(2016年5期)2016-03-01
专用汽车(2016年5期)2016-03-01
食品工业科技(2014年23期)2014-03-11
