一年生钟萼木生长性状遗传分化与优良个体早期选择研究
2019-07-22 13:16郭祥泉钱国钦吴载璋涂年旺郭祥堆
亚热带植物科学 2019年2期
郭祥泉,钱国钦,吴载璋,涂年旺,吴 炜,郭祥堆
(1.福建省三明市林业局,福建 三明 365000;2.福建省世界银行贷款造林项目办公室,福建 福州 350001;3.福建省永安市林业局,福建 永安 366000;4.福建省永安林业(集团)股份有限公司,福建 永安 366000;5.永安市第六中学,福建 永安 366000)
多数林木树种为了能更好地适应不同自然环境,其繁育机制逐渐进化成杂交繁育,以杂合型个体生存。杂合型个体杂交子代遵循基因重组规律和数量性状遗传的基因积加效应,分化为抗旱、抗病、耐瘠薄、不同速生性、抗风以及其他抗逆性等多样性遗传特性,使物种能够适应不同生境,提高物种生存机会[1—3]。钟萼木Bretschneidera sinensis为异花授粉树种,杂交子代的胸径与树高生长性状表现多样性遗传,生长性状在生长早期表现出显著分化[3]。Ying 等[4]报道美洲黑杨10 年生与3 年生树高生长相关性(r=0.61~0.96)。梁一池[5]报道杉木早期树高生长与其后各龄树高生长存在显著或极显著相关,且随着年龄比值的增大,相关关系愈密切。高与胸径生长的早期选择具有显著的遗传相关性,因此在林木幼树期进行超级苗筛选,能获得较好遗传品质个体,杂种优势个体适宜苗木期筛选[1,6—9]。事实上,自然选择本身就是早期选择过程,在天然更新环境中,种子萌芽与幼苗生长过程,小苗需经过病虫害、干旱、水渍等逆境选择和个体竞争,选择适宜生长的个体。故天然更新保存下来的个体是早期选择后的适宜生长个体,特别是自然更新较难的物种,所保存下来的个体本身就是一种能适应环境生长的个体[9—10]。作者对4 年生钟萼木进行优良个体选择,所筛选优良个体后续表现出良好生长效果[11],一年生优良个体选择未见报导;对钟萼木一年生苗木群体高与地径生长性状遗传多样性研究表明,群体高与地径性状生长权重表现为多样性正态分布,优良个体与弱势个体年生长量相差数倍,表现显著分化。生长性状分布表明,杂种优势个体属于小概率群体,进行一年生早期筛选具有科学的理论依据[3]。利用天然自由授粉子代进行优良个体选择在遗传育种中属于混合选择方法,是林木良种选育初期选择常用方法[1]。本研究根据钟萼木生长性状遗传多样性与早期生长性状分化特性,应用t 检验选择法对该树种的高与地径生长性状进行早期选择,筛选优良基因型个体,从而为该树种收集优良种质资源。
1 材料与方法
1.1 试验地概况
试验地位于福建省大田县桃源镇赤头坂国有林场场部工区,属于戴云山西延的低山带,为中亚热带季风气候。年平均气温15.3~19.6 ℃,无霜期255~275 d,年均降水量1491.2~1809.6 mm。区域内气候温暖湿润、雨量充沛,适宜钟萼木生长(气象资料来源于大田县气象局)。试验地海拔325~605 m,土壤为黄红壤。
试验圃地为农田,沙壤土;于2015 年3 月进行整地,畦面宽90 cm、沟深20 cm,过磷酸钙作为基肥,按100 kg·667m-2整地前散施,整地后播种前用600 倍多菌灵消毒。于2015 年3 月下旬播种,播种采用点播,在畦面挖10 cm 深条沟,条沟间隔20 cm,种子间隔20 cm 点播,用细黄心土覆盖,及时喷水至畦面潮湿。种子来源于福建省永安天宝岩自然保护区采种母树,经休眠催芽处理,出芽后搭盖遮阳网。
1.2 方法
钟萼木一年生苗木生长于环境条件基本一致的苗圃地,环境因子对个体生长影响基本相同,个体生长差异来源于遗传分化。生长性状遗传多样性调查采用大样本调查方法,该方法适宜基因重组后性状表现的分类统计;同一圃地中优势木分布是由随机播种后产生的,选择苗木高生长优势木作为调查群体,预选中心设置调查样点属于完全随机概率。根据试验调查要求进行整体样本踏查,预选优势木,然后设置调查样点。以优势木为调查中心,共设置6 个调查样点,每样点内调查个体不少于500 株,有多株优势木处于同一调查群体的,按一个调查群体处理,所有优势木都作为预选木,优势木不在同一群体的,另设置一个调查群体,每个群体分别调查苗木地径与苗高[3,9]。各调查样点的优势个体生长情况见表1(群体Ⅱ无特别优势个体),记录调查样点内优势木生长状况,对优势木按群体顺序编号。地径测量采用游标卡尺,苗高测量采用钢卷尺。于2015 年12 月8 日当年苗木生长期结束后进行生长性状调查,分别测量苗木地径与苗高。
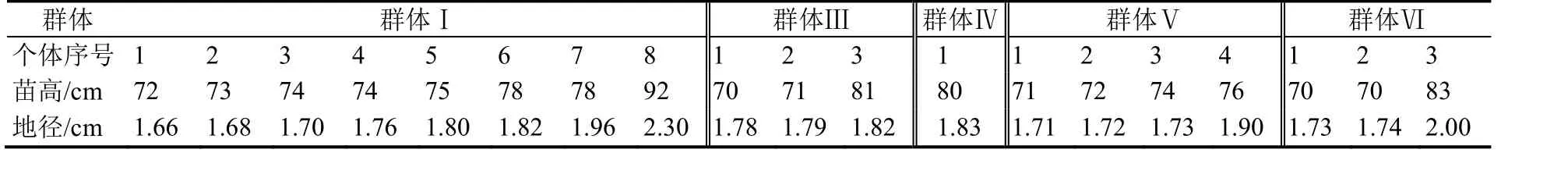
表1 不同群体优势个体生长性状分布Table 1 Growth distribution of the dominant individual in different investigating colony unit
生长性状遗传多样性采用大样本调查方法,应用正态分布标准差分类法,划分地径与树高生长性状的不同生长类型,应用不同生长类型评价生长性状遗传分化[1—3]。优良个体筛选根据性状遗传分化结果,应用t 检验选择法,确定优良个体选择标准,然后进行优株筛选。已有研究认为,林木生长在未受人为干扰条件下,群体的高与地径生长呈正态分布,适宜应用正态分布标准差分类法对群体生长性状进行分类,以及应用t 检验选择法筛选优良个体[6,10—12]。
1.3 选择标准确定
根据t 检验选择法,应用表2 中合并群体的相关统计值与t 检验值计算选择标准值,确定苗高与地径的选择标准,依据选择标准选择优良单株。苗高、地径生长性状选择标准临界值依据t 检验选择法计算,临界值计算公式:
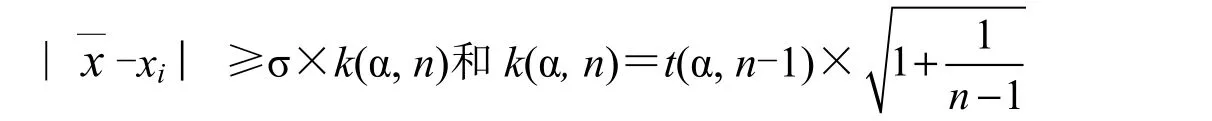

以该值为临界值,所筛选的优良基因型就是群体中的优势个体。故临界值可作为筛选优良基因型的入选标准。

当n 在500~1000 区间,t(0.001,n-1)=3.291,k(0.001,n)=3.293
n 为调查群体的个体数,依据上式与t 分布值可得k(0.001, n),由此得调查选择优势个体的临界值,由上述σ 值和式(1)、(2)计算得群体苗高、地径选择标准指标分别是71.2 cm、1.91 cm。
1.4 数据处理
上述不同样点的样本可共同应用于生长性状遗传多样性分化与个体筛选,合并作为总调查样本进行统计分析[10],总样本的方差、标准差等相关统计值见表2。依据正态分布标准差分类法,按一个标准差值作为不同生长类型的区间临界值划分生长类型[4,10]。在实际生产中,平均值的生长应为一个群体区间分布,本试验以作为平均值生长类型,其他以此类推分别为,生长区间作为同一群体不同生长类型,相关数据以此进行归类[4,13—14]。
优良个体筛选的预选优势个体单独调查相关因子见表1,优势木各样点调查按地径、苗高生长分别统计不同群体的平均值、方差、标准差和波动系数(表2),遗传分化与优株筛选的苗高生长平均值、标准差与区间上下限取整数,地径生长平均值、标准差与区间上下限取2 位小数。
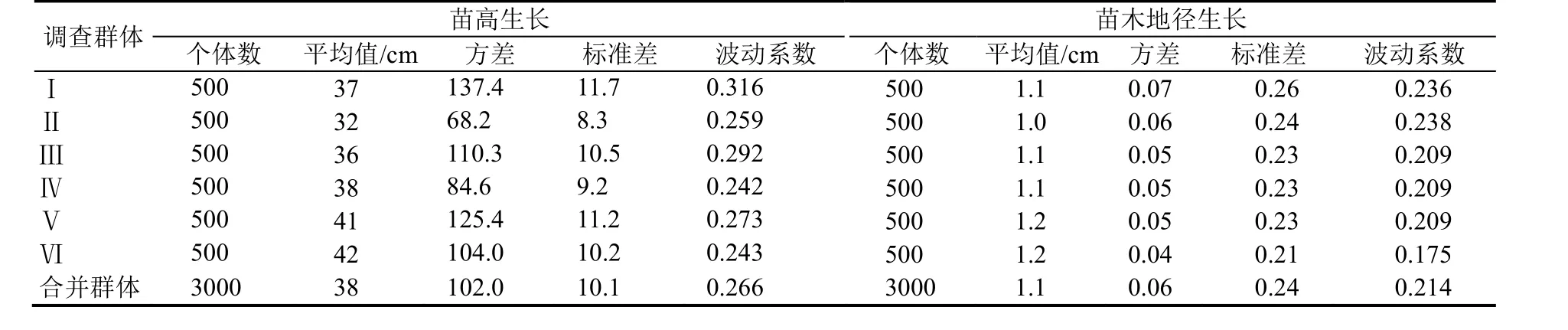
表2 不同群体相关统计值Table 2 The statistics value of different colony
2 结果与分析
2.1 钟萼木苗高与地径生长分化表现
2.1.1高生长习性遗传分化钟萼木一年生苗木高生长划分为8 个不同遗传品质生长类型(3~13 cm、13~23 cm、23~33 cm、33~43 cm、43~53 cm、53~63 cm、63~73 cm、73 cm 以上),各生长类型权重依次分别为0.13%、6.70%、24.43%、36.63%、23.13%、6.87%、1.40%、0.37%(图1)。以上结果显示,平均值所在区间占比最大,其次是平均值相邻的两个生长类型权重。各类型权重由平均值所在区间逐渐向两边呈正态分布。生长性状表现由基因型遗传表现为不同生长类型,理论上属于不连续的性状表现,数量性状遗传在环境因子与基因互作累加效应作用下,同一遗传类型的性状表现不完全一致,在实际生长中呈连续性分布,因此较难分清不同的遗传类型,利用上述方法把连续性性状表现划分成不同遗传类型,可以较好弄清不同遗传类型的性状分化。从不同遗传类型的性状分化可以较好探讨不同遗传类型生长适应性与遗传的相关性,了解生长表现差的、平均值及相近类型与优良生长类型的遗传相关性。由不同类型的权重比例,可以推测完全显性表现与隐性表现的概率以及多基因中部分显性与隐性个体的表现[1]。由基因遗传特性可知,通常更多显性基因型表现为杂种优势生长类型,完全隐性基因型表现为较差生长性状类型。由图1 可知,群体中杂种优势生长类型占比为0.37%,该类基因型绝大多数或全部控制基因是显性的,该类型生长是具有遗传表现的生长类型;生长表现差的生长类型占比为0.13%,由基因型表现可知,该类型属于完全隐性基因或多数控制基因型是隐性基因,多数等位基因为纯合型。从不同性状表现分化与不同生长类型划分,能较好地了解该物种主要经济生长性状的遗传表现和生长潜能[3,10]。
2.1.2地径生长习性遗传分化高生长与地径生长密切相关,苗木地径生长与高生长具有一致性,是评价苗木质量的重要生长指标。在个体空间竞争不剧烈条件下,地径生长与高生长的遗传分化遵循一致的规律。钟萼木一年生苗木地径生长划分为8 个不同生长类型(0.28~0.52 cm、0.52~0.76 cm、0.76~1.0 cm、1.0~1.24 cm、1.24~1.48 cm、1.48~1.72 cm、1.72~1.96 cm、1.96 cm 以上),各生长类型权重分别为0.63%、6.40%、23.30%、38.37%、24.17%、6.40%、0.67%、0.07%,与苗木高生长类型分化表现基本一致,地径不同遗传类型的性状分化表现为差、平均值及相近类型与优良生长类型(图2)。平均值及相近类型所占比例分别为 23.3%、38.37%、24.17%,特别优良生长类型占比为0.07%,生长表现差的生长类型占比为0.63%,地径生长分化与高生长类型分化表现基本是一致的,通过地径和高生长性状共同评价主要生长性状的遗传表现,能更好地了解该树种的生长潜能和遗传相关性[3]。
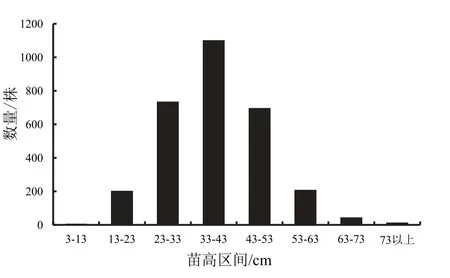
图1 苗高生长不同遗传品质分布Fig.1 Distribution of seedling height growth of different genoptype
2.2 优良个体的选择
2.2.1优株选择根据优良个体选择标准,由表1 中优势个体筛选得群体Ⅰ的1~8 号、群体Ⅲ的3 号、群体Ⅳ的1 号、群体Ⅴ的2~4 号和群体Ⅵ的3 号共14 株;以地径生长为选择指标,达到标准要求的优势个体是群体Ⅰ的7、8 号、群体Ⅴ的4 号和群体Ⅵ的3 号共4 株。经综合评价,群体Ⅰ的4、5、6 号,群体Ⅲ的3 号,群体Ⅳ的1 号在高生长上有潜在优势,且地径生长处于选择标准的临界值附近,认为可选择为优良个体。因此,最终选择结果为群体Ⅰ的4~8 号、群体Ⅲ的3 号、群体Ⅳ的1 号、群体Ⅴ的4 号和群体Ⅵ的3 号共9 株,所筛选的优势个体苗高、地径生长分别见表1 记录。将群体Ⅰ的4~8 号、群体Ⅲ的3 号、群体Ⅳ的1号、群体Ⅴ的4 号和群体Ⅵ的3 号依次命名为田钟1~9 号,并进行建档保护,作为优良基因资源。
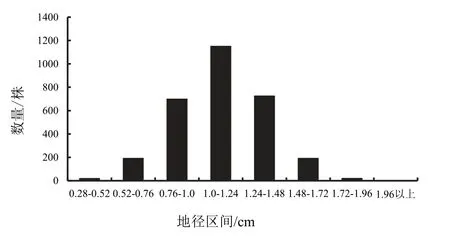
图2 地径生长不同遗传品质分布Fig.2 Distribution of caliper growth of different genoptype
2.2.2选择结果根据t 检验选择法,所筛选个体是分布于区间以外的特优生长类型,具有良好的杂交优势。由所筛选个体的主要生长性状表现,田钟1~9 号的高生长比平均值分别提高95%、97%、105%、105%、142%、113%、111%、100%、118%;地径生长比平均值分别提高57%、61%、63%、75%、105%、63%、64%、57%、79%。优良个体高与地径生长性状与群体平均值比较,表现出较好的遗传增益,说明优良个体主要生长性状具有很好的速生潜能。
3 讨论
钟萼木是国家一级保护濒危树种,对人体内外伤治疗有良好功效。钟萼木主要为天然分布[15],近年来才开始人工培育,人工林尚处于幼树阶段,优良种质资源筛选与利用较缺乏。现有人工培育幼林来源于天然种源,个体生长表现为不同的生长适应性,遗传品质有待改良[3,13—14]。
钟萼木一年生苗木主要生长性状表现显著分化,苗木群体一年生高生长权重呈连续型正态分布,群体平均生长量仅38 cm,生长最好个体可达92 cm,最差个体为13 cm。应用正态分布标准差分类法,群体划分为8 种不同生长类型,高生长表现为明显的遗传分化与多样性生长[3],不同生长类型差异明显,群体84%以上的高生长处于平均值与相邻两生长类型。地径生长与高生长表现基本一致性,地径平均值仅1.12 cm,最好个体生长量达2.3 cm,最差的个体生长量为0.3 cm,应用正态分布标准差分类法,群体划分为8 个不同生长类型,群体85%以上的地径生长处于平均值与相邻两生长类型,遗传分化与多样性生长显著。生长性状遗传多样性研究表明,钟萼木虽然处于濒危状态,但所保存种质资源具有不同遗传品质生长类型,个体多样性生长性状中具有多种类型基因型,也存在优良的遗传品质资源,筛选优良个体能改善种质资源[3]。
遗传多样性分化与前人的林木育种研究表明,杂种子代表现早期生长分化,分化的不同遗传类型是由基因类型决定,具有遗传效果,适宜进行早期选择[1,7—8]。本研究所筛选个体是分布于区间以外的特优生长类型,属于自然杂交的杂种优势,是一种具有遗传效果的基因类型,具有选择价值。以苗木的高和地径生长作为双重指标,依据t 检验选择法确定选择标准,经综合评价,确定筛选优势个体,选择方法具有科学性,所选优良个体在群体生长中有很好生长表现,遗传增益显著,有良好的潜在利用价值。
猜你喜欢
区域治理(2022年40期)2022-11-27
清华金融评论(2022年4期)2022-04-13
武汉工程职业技术学院学报(2022年1期)2022-04-13
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
数学物理学报(2020年2期)2020-06-02
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
文理导航(2015年14期)2015-05-22