
内蒙古典型草原群落内部植物和土壤空间异质性
2019-07-20 03:40石椿珺李艳龙程建伟张桐瑞李永宏
草业科学 2019年6期
石椿珺,李艳龙,程建伟,张桐瑞,郭 颖,李永宏
(内蒙古大学生态与环境学院 / 蒙古高原生态学与资源利用教育部重点实验室 / 省部共建草地生态学国家重点实验室培育基地,内蒙古 呼和浩特 010021)
植物群落空间异质性及其动态是认识生态系统生物多样性和稳定性维持机制的基础。植物群落空间异质性是指其物种组成和结构在空间上的不均匀分布形成的生态格局,通常具有斑块镶嵌以及随环境梯度变化的特点[1],并与微地形和气候等自然条件以及人类活动的影响紧密相关[2-3]。天然草原植物群落的斑块镶嵌结构是草原生态系统固有的特征,反映了草原生态系统中生物与环境因子之间以及不同生物类群之间的相互作用,影响生态系统的生物多样性和功能[4-5]。植物群落的空间异质性及其驱动力分析是认识植物生态学的核心问题,一直为生态学界所关注[6]。
大量的研究表明,植物群落的空间异质性与土壤的养分和水分相关。土壤养分供给能力,尤其是碳(C)、氮(N)、磷(P)养分的供给能力,影响植物群落的物种组成和生产力[7-11]。随着土壤养分的变化,适应不同土壤养分环境的植物种会相互替代[12-13],进而改变土壤养分的空间异质性,影响植物对资源的利用率,形成植物群落的镶嵌分布[14-15]。气候变化和人类活动都会直接或间接地影响生境中的养分和水分资源分布,增大或减小其异质性,从而影响物种在小尺度上的空间分布[16-17]。在较小尺度上探究草原植物群落空间异质性对了解植物群落格局及形成机制具有重要意义[18]。然而国内外关于群落斑块空间异质性的研究多集中于大尺度的景观格局以及植物的斑块化分布对动物采食行为的影响等方面,而对于小尺度的草地微斑块方面的探究较少[5]。本研究在内蒙古高原中东部的锡林郭勒地区,选择典型草原的代表性植物群落,通过系统设立研究样点,同时调查样点植物群落特征和土壤性质,研究植物群落内部的空间异质性,探讨其与土壤空间异质性间的关系,分析空间异质性形成机制,以期为草原生物多样性保护和可持续管理提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古自治区锡林浩特市毛登牧场境内,是内蒙古大学草地生态学研究基地的典型草原示范牧场,地理坐标为 116°31′ - 116°32′ E 和44°15′ - 44°15′ N。该区域具有温带半干旱草原气候,冬冷夏热,昼夜温差大,年均温在0.5~1.0 °C;年降水量280.5 mm,多集中在夏季,年际变率大;年蒸发量1 600~1 800 mm,远大于降水量;干燥度在1.50~3.99。土壤为典型栗钙土,植被为典型草原,以旱生和中旱生草本植物为主,优势植物有大针茅(Stipa grandis)、羊草(Leymus chinensis)、糙隐子草(Cleistogenes squarrosa)、黄囊苔草(Carex korshinskyi)和猪毛菜(Salsola collina)等。
1.2 研究草原样地
研究草原在2012年前是天然的典型草原放牧场,2012年开始围封禁牧,2016年建立了草原放牧试验样地。该样地地势较为平坦,有轻微南向坡度(< 2%),总面积13.5 hm2,划分54个0.25 hm2(50 m × 50 m) 的小区。为了解草原本底特征,尤其是不同小区间草地有无空间异质性,2017年初,在每个小区的4个方位规则放置4个1.2 m × 1.2 m的围笼,使其免受动物放牧的影响,共设置216个样点 (12 行 × 18 列,排列为平均距离为 25 m × 25 m的方格)。8月中旬在设置围笼的216个点调查植物和土壤特征,获取植物和土壤样品。
1.3 植物和土壤样品采集
在每个样点,用1 m × 1 m样方记录植物种名,分种测定植物高度和盖度,剪取植物,带回实验室,记录其鲜重及干重(65 °C烘箱烘至恒重);存留植物样品供化学元素含量分析。在剪去每个样方里的植物,沿对角线用土钻(直径5 cm)取3钻0-10 cm表层土壤样品,三钻合一,装入塑封袋带回实验室,自然阴干,用于土壤化学特性分析;同时采用环刀法测定0-10 cm土层土壤容重和含水量,环刀容积为100 cm3。
1.4 植物和土壤样品分析
植物C、N、P含量通过vario ELIII,Elementar,Germany元素分析仪测定;土壤有机碳(SOC)采用有机总碳分析仪Liqui TOC测定,全氮(TN)采用vario ELIII,Elementar,Germany元素分析仪测定,全磷(TP)用浓酸消解-磷钼蓝分光光度法测定[19];土壤pH采用pH测试仪测定。
1.5 数据处理
分析216个取样点的植物群落和土壤形状间的关系。首先根据群落中不同物种的生物量,通过K-均值聚类方法,并优化聚类数[20],将不同样地的植物群落划分为不同的植物群落斑块类型,对每类群落斑块依照其优势度高的植物种命名;然后使用方差分析检验不同斑块类型之间植物或土壤性质的差异性,并分析植物与土壤性质之间的关系;最后用冗余分析(RDA)综合分析不同群落斑块植物与环境因子关系。所有统计分析采用SPSS Statistics 22.0与Canoco 4.5进行。
2 结果与分析
2.1 植物群落斑块类型及其物种组成
根照植物群落的物种生物量组成,采用K-均值聚类方法将216个群落斑块聚为4个类型,分别以其优势植物命名为针茅斑块(65个)、羊草斑块(50个)、猪毛菜斑块(23个)以及混合斑块(78个)。混合斑块中针茅、羊草和糙隐子草具有相近的优势度(表1)。除优势种外,不同斑块的共有种还有展枝唐松草(Thalictrum squarrosum)、矮葱(Allium anisopodium)、刺穗藜(Chenopodium aristatum)、灰绿藜(C.album)和鹤虱(Lappula intermedia)。上述4个优势种和5个共有种的生物量可占群落总生物量的99%。其他记录到的植物种还有麻花头(Serratulla centauloides)、狗尾草(Setaria viridis)、草木樨状黄芪(Astragalus melilotoides);羊草斑块记录到的独有物种有画眉草(Eragrostis pilosa)和尖头叶藜(C.acuminatum);猪毛菜斑块独有物种有小叶锦鸡儿(Caragana microphylla)、细叶鸢尾(Iris tenuissimum)和黄花葱(Allium condensatum);混合斑块独有物种有冰草(Agropyron cristatum)、野韭(Allium ramosum)、多根葱(A.polyrhizum)、冷蒿(Artemisia frigida)和薄荷(Mentha canadensis)。
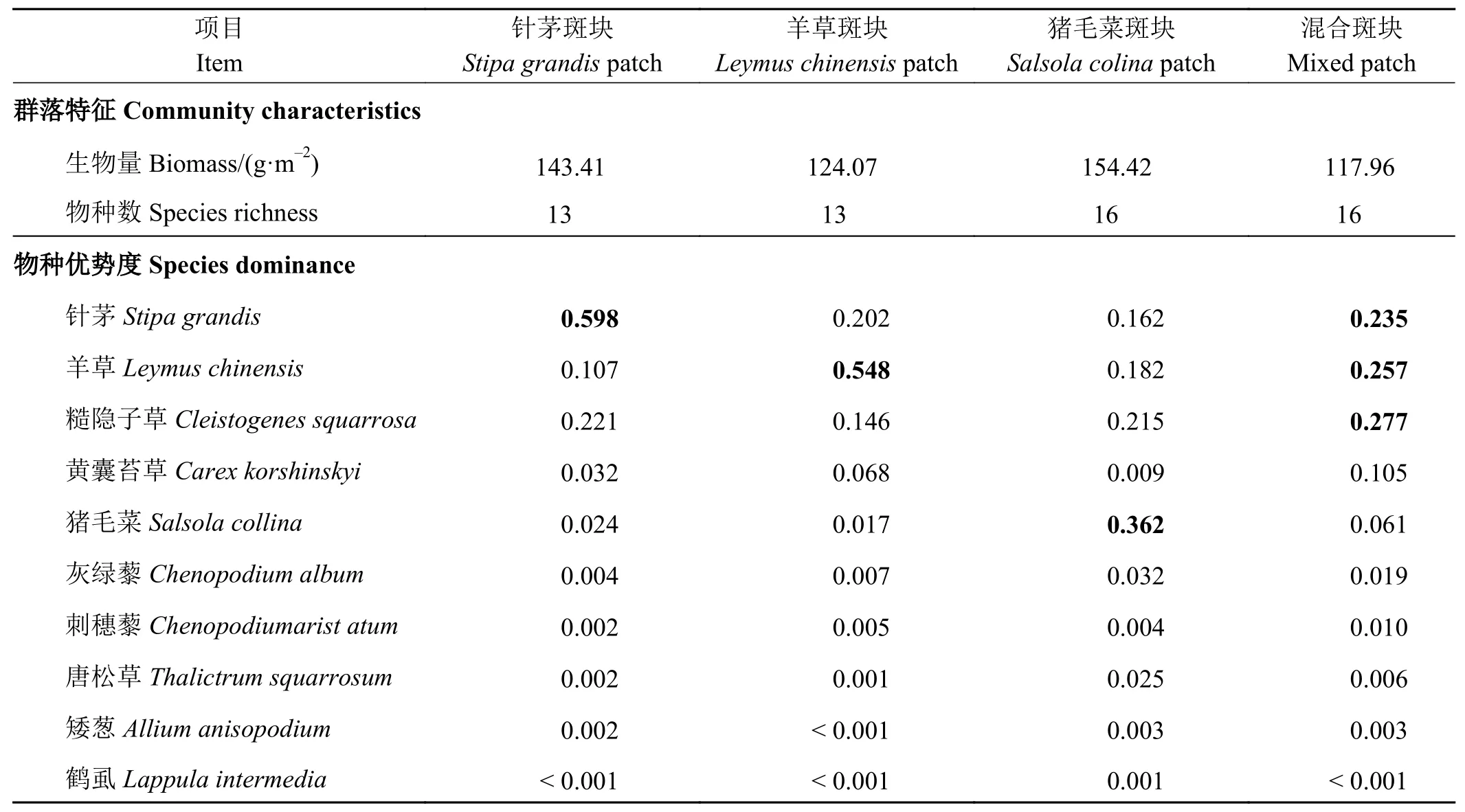
表1 4种植物群落斑块的群落特征和优势种组成Table 1 Dominant species and biomass composition of plant communities in four patch types
2.2 不同斑块植物群落及土壤特征分析
2.2.1 不同群落斑块的植物生产力和多样性
植物群落的地上生物量在猪毛菜斑块最高,显著高于其他斑块(P< 0.05),而针茅斑块的地上生物量也显著高于羊草斑块和混合斑块(图1A)。植物群落的物种丰富度也在猪毛菜斑块显著高于其他斑块(P< 0.05),在混合斑块高于羊草斑块(P< 0.05)(图1B);群落中植物种的均匀度在猪毛菜斑块和混合斑块显著高于针茅和羊草斑块(P< 0.05)(图1C);群落的多样性指数从高到低的次序为猪毛菜斑块 >混合斑块 > 针茅斑块 > 羊草斑块 (图1D)。
2.2.2 不同斑块植物化学计量特征
植物群落中针茅、羊草、糙隐子草、黄囊苔草和猪毛菜是研究草地中最优势的5种植物,其生物量占群落总生物量的93.5%~98.2%(表1),这些物种间的C、N、P含量均存在差异,其中猪毛菜的C含量显著低于其他物种(P< 0.05),而大针茅的P含量显著低于其他物种(P< 0.05)(表2)。

图1 不同斑块类型植物群落生产力和多样性比较Figure 1 Plant biomass and diversity of grassland communities in different patches
针茅物种的C含量在针茅斑块中(46.89%)显著大于羊草斑块(45.76%)(P< 0.05),但羊草、糙隐子草的C含量在不同斑块中没有差异;针茅、羊草、糙隐子草的P含量在不同斑块之间也没有差异(P>0.05)。针茅、羊草、糙隐子草的N含量均在针茅斑块中最低(分别为2.39%、2.71%和2.22%),并显著低于混合斑块(分别为3.09%、3.45%和2.64%)(P<0.05),其他两斑块的含量介于二者之间。
根据主要优势种在群落生物量中的比例(表1)及其C、N、P含量(表2)估算植物群落地上生物量的C、N、P含量(表3)。群落植物地上部C含量在猪毛菜斑块显著低于其他斑块(P< 0.05),而其他斑块之间没有显著差异(P> 0.05);N含量在混合斑块中最高,显著高于大针茅斑块(P< 0.05);P含量在针茅斑块显著低于其他斑块(P< 0.05),其他斑块之间没有显著差异(P> 0.05)。大针茅斑块的C∶N和C∶P显著高于其他3种斑块(P< 0.05)。整体上看,猪毛菜斑块群落植物C含量低,N、P含量高;针茅斑块群落植物C含量高,N、P含量低。
2.3 不同草地斑块的土壤特征
土壤表层(0-10 cm)含水量在4种斑块类型间没有显著差异(P> 0.05)(表4)。土壤容重在猪毛菜斑块显著高于针茅和羊草斑块P< 0.05)。土壤pH在猪毛菜斑块显著低于其他3种斑块(P< 0.05)(猪毛菜斑块 < 混合斑块 = 羊草斑块 < 针茅斑块)。土壤SOC含量在猪毛菜斑块的显著低于其他斑块(P< 0.05),TN、TP含量也在猪毛菜斑块较低;而针茅斑块的土壤SOC、TN含量均显著高于其他斑块(P< 0.05),TP含量显著高于羊草与猪毛菜斑块(P< 0.05);羊草斑块与混合斑块的SOC、TN、TP含量相近,均低于针茅斑块,高于猪毛菜斑块。土壤C∶N和C∶P在4种不同斑块间没有显著差异(P> 0.05)。土壤0-10 cm容重和pH空间异质性低(变异系数CV小于0.1),而土壤含水量、SOC、TN有一定的空间变异 (0.13 < CV < 0.16),TP 空间异质性最大(CV = 0.28)。
2.4 植物群落化学计量特征与土壤指标间的关系
植物地上部分P含量与土壤SOC、TN、TP显著负相关(P< 0.05);地上部分植物C∶P与土壤SOC、TN、TP 极显著 (P< 0.01)或显著 (P< 0.05)正相关。植物C、N含量与土壤各指标之间没有显著相关关系(P> 0.05),植物C含量与土壤SOC有正相关的趋势(P= 0.065)。植物地上部化学计量特征与土壤pH 之间没有显著关系 (P> 0.05)(表5)。
植物与土壤环境因子冗余分析(RDA)结果(图2)显示,第1和2轴特征值分别为8.4%和2.4%;所有排序轴的蒙特卡洛置换检验呈极显著差异(P=0.002)。土壤pH、TP、TN与第1排序轴相关性较大,其方向与针茅的优势度方向一致,表明针茅物种与这些土壤因子的梯度变化正相关;而羊草、黄囊苔草与猪毛菜与这些因子的梯度变化负相关;土壤SM、BD、SOC与第2排序轴相关性较大,草原建群植物针茅和羊草的方向,显示其与土壤SM和SOC正关联,而糙隐子草和猪毛菜等放牧退化群落的优势度方向与土壤BD正关联。
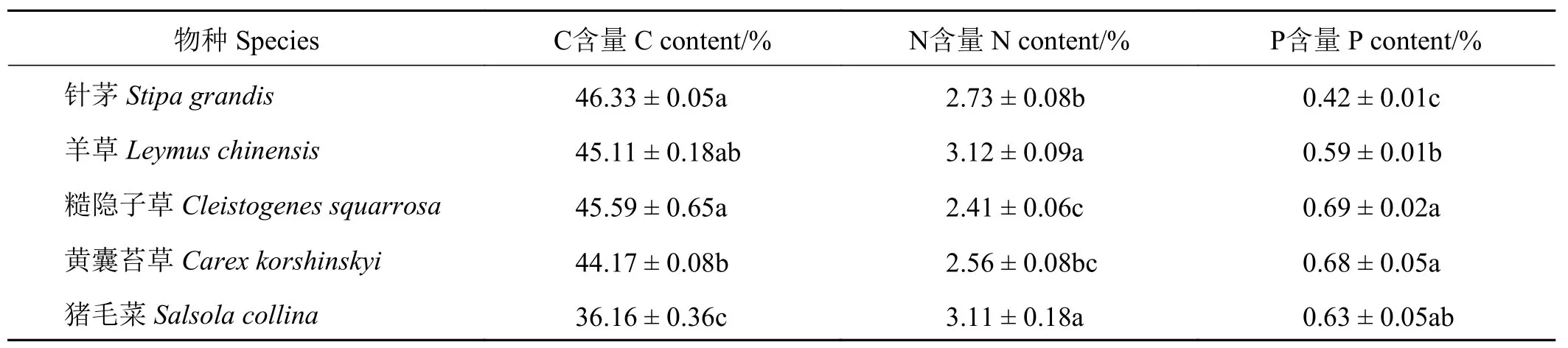
表2 优势种植物地上部分化学计量特征Table 2 Stoichiometric statistics of dominant species

表3 不同斑块草原植物群落地上部分化学计量特征Table 3 Stoichiometric statistics of plant communities on different patches

表4 不同植物群落斑块土壤化学计量统计Table 4 Stoichiometric statistics of the soil under different plant community patches

表5 群落地上部分与土壤各指标之间的相关关系Table 5 Correlation between the aboveground parts of plants and soil indexes
3 讨论
3.1 植物群落斑块的性质
空间异质性是生态系统的固有特性,反映了自然生态因子或干扰的空间分布,及其造成的植物或土壤结构与功能上的空间差异[21]。研究不同尺度的生态异质性有助于判断与分析自然或人类因素对生态系统格局与过程的影响,进而为生态系统管理和退化生态系统修复提供基础[22]。依据植物在群落中的优势度,划分研究植物群落为4种斑块类型,其中针茅或羊草占优势的斑块是典型的天然草原群落;糙隐子草与针茅和羊草共占优势的混合斑块往往是天然草原在放牧利用后演替中的群落[23];一年生草本猪毛菜占优势的斑块,实际上是春季草地上植被稀疏甚至裸露的斑块,夏雨后一年生草本迅速生长。本研究表明,天然草原建群植物占优势的针茅斑块中针茅、羊草和糙隐子草植物种水平及群落水平的N含量均显著低于混合斑块,说明植物N含量在斑块类型间的差异不仅来自不同的物种组成,也来自同一物种N含量随斑块类型的变化。植物群落C、P含量在斑块类型间的变化则主要受优势种C、P含量的影响,猪毛菜斑块的群落C含量显著低于其他斑块群落,主要是由于猪毛菜的C含量显著低于其他物种;针茅斑块植物群落的P含量显著低于其他斑块,主要是由于针茅的P含量显著低于其他物种。

图2 植物与环境因子的冗余分析(RDA)Figure 2 Bioplot diagram of redundancy analysis (RDA)of species constrained by environmental factors
3.2 植物群落空间异质与生物多样性
草原植物群落空间异质性有助于生物多样性的维持[5]。4种斑块类型植物的α多样性(物种丰富度和多样性指数)具有明显的差异,混合斑块与猪毛菜斑块的多样性高于以针茅和羊草为优势的2种斑块,并且有9种植物只在4种斑块之一中出现。不同斑块植物群落间的差异本身是草地β多样性的组成,斑块结构体现的草地空间异质性在很大程度上影响了植物群落的物种丰富度与多样性。植物群落空间异质性在维护群落生物多样性的作用,已为很多研究所肯定[24-25]。
3.3 植物和土壤空间异质性间的关系
植被与土壤的空间变化具有对应性[26],土壤养分在植物群落空间格局中的作用为许多研究所证实[27-28]。本研究是在外貌非常均匀的试验草地上,测定的不同样点上草原植物和土壤性状的变化,样点间距约25 m。试验草地不同斑块类型的样点间没有可观测到的微地形差异,同时测定的土壤表层(0-10 cm)含水量在不同斑块类型之间也没有差异,说明微地形和及其相关的土壤含水量并不是导致斑块间植物群落差异的主要原因。土壤pH和养分(TN、TP)含量在针茅斑块显著高于其他斑块(P< 0.05),而在猪毛菜斑块最低;相反,土壤容重在针茅和羊草斑块显著高于猪毛菜斑块;草原的空间异质性可能与引起土壤养分和容重变化的驱动因子相关。
本研究表明,植物C含量与土壤SOC含量在斑块间的变化是协同的,如针茅斑块植物C含量与土壤SOC含量均高于其他斑块,猪毛菜斑块植物C含量与土壤SOC含量均低于其他斑块。这个结果与Dan等[29]的研究结果一致,显示多年生丛生禾草对土壤碳具有聚集作用。本研究也表明,植物N、P含量与土壤TN、TP含量在斑块间的变化是相反的,如针茅斑块植物N、P含量低于其他斑块,而猪毛菜斑块植物N、P含量高于其他斑块;但土壤中TN、TP含量在不同斑块间的变化趋势则相反,针茅斑块高于猪毛菜斑块。冗余分析(RDA)的结果表明,针茅的优势度与土壤TN、TP、SOC含量具有相同的变化趋势,与土壤容重具有相反的变化趋势;而猪毛菜的优势度与土壤TN、TP、SOC含量的变化趋势相反,与容重的变化趋势一致;说明以猪毛菜为优势的斑块土壤养分含量低,容重大,因而春季植物密度低,夏雨后土壤有机质快速分解释放养分,同时中生性的一年生植物猪毛菜等迅速生长,形成占优势群落层片。这个结果说明,斑块的存在是土壤养分和植物生活型动态互作的结果。
3.4 草地植物土壤空间异质性的原因
在景观尺度上,水、热等环境条件分异是植被空间异质性的主要原因[30]。在较小微地形尺度上,与微地形关联的土壤水分和养分变化与植被的空间异质性紧密相关[31]。在本研究的微小尺度上,草地的空间异质性可能有三方面的原因,其一是微小地形变化导致的土壤水分和养分的空间变化[32];其二是植物营养繁殖(如针茅枝条分蘖和羊草根茎繁殖),从占有区域逐步向外拓展,形成植物种群的斑块状分布[33-34];其三是放牧动物对草地选食、践踏和排泄物归还的空间变化。大量的研究表明,草原放牧尤其过度放牧会降低土壤的养分,增大土壤紧实度和容重[11,33];退化草原恢复过程中早期放牧对草地植物与土壤的空间异质性的影响会在较长时间内存在[35]。本研究结果显示,不同植物群落斑块所对应的土壤养分和容重间具有显著差异,而微地形及其相关的土壤水分含量没有差异,由此推论,草地植物群落在斑块尺度上的空间异质性,很可能与该尺度上不同放牧影响的异质性相关,而不是微地形的差异所致。草地斑块结构的成因和变化,需要进一步深入研究草地空间结构动态。
4 结论
内蒙古草原区以针茅和羊草为优势的典型草原群落具有斑块结构。这些群落斑块包括以多年生针茅或羊草为优势的天然草原群落和多种多年生植物共优势的群落,以及一年生植物优势群落。草原植物群落的空间异质性有助于维持草原群落的生物多样性。多年生丛生禾草针茅斑块具有聚集养分的作用,土壤有机碳和氮磷元素相对丰富,容重低;一年生植物猪毛菜斑块土壤养分低,容重大,限制了草原优势植物的生长及其对群落伴生植物的抑制。草原植物群落斑块异质性与土壤pH、容重及养分含量的变化具有对应性,而与土壤水分没有相关性。长期动物放牧影响的空间异质性可能是研究草原植物群落斑块尺度空间异质性的主要原因。
猜你喜欢
临床肝胆病杂志(2022年8期)2022-11-23
当代水产(2022年8期)2022-09-20
中国临床医学影像杂志(2022年6期)2022-07-26
中国农业科学(2022年11期)2022-06-27
草原与草业(2022年1期)2022-04-07
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
现代畜牧科技(2021年4期)2021-07-21
现代企业(2021年2期)2021-07-20
种子(2020年4期)2020-12-22
