
抗寒苜蓿品系生产性能评价及ISSR遗传多样性分析
2019-07-19 07:28魏双霞师尚礼康文娟谭谌淼
草原与草坪 2019年3期
魏双霞,师尚礼,康文娟,谭谌淼
(1.甘肃农业大学 草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
苜蓿具有产量高,粗蛋白含量高,适口性好的特点,在世界范围内广泛种植[1-2]。青藏高原素有 “世界屋脊"和 “世界第三极"之称[3],是我国重要的草地畜牧业基地之一,但过度放牧使天然草地产量下降,毒杂草滋生,牧草品质降低[4],造成严重的经济损失。解决这一问题,需要了解紫花苜蓿对寒冷地区的适应性特征和规律。因此,深入研究苜蓿的生产性能和遗传基础,选育出适应于该地区种植的苜蓿新品系,扩大其种植范围,提高生产力,具有举足轻重的意义。
目前,我国关于苜蓿品种适应性评价的研究主要在干旱半干旱地区,朱博等[5]、王虹等[6]通过综合评价筛选出在西北荒漠地区种植的高产优质苜蓿品种。而对苜蓿品种在高寒地区适应性的研究很少,其中丁培俊等[7]在甘南州进行了苜蓿引种品比试验,挑选出适宜于青藏高原种植的传统苜蓿材料金皇后和阿尔冈金。杨瞾等[8]在东北寒冷地区对引种的 16个苜蓿材料应用灰色关联度进行生产性能评价,筛选出适应性较好的品种有俄罗斯西伯利亚杂花等。采用3份抗寒苜蓿品系和3份对照苜蓿材料,进行生长适应性对比试验,综合评价生产性能,以期为高寒牧区培育多年生抗寒苜蓿新品系,发展苜蓿产业提供技术依据。
分子标记是以DNA多态性为基础的遗传标记,在苜蓿生物技术育种和种质资源评价中的地位不可忽视[9]。李哲等[10]用ISSR在遗传距离0.266处将供试苜蓿材料聚为5大类,为其后代株系杂交组合育种的研究提供理论支持。杨瞾等[11]用 ISSR在遗传距离0.833处将供试抗旱苜蓿材料分为高产、优质和营养价值相对较低2大类。应用ISSR分子标记方法研究3份抗寒苜蓿品系和3份对照苜蓿材料俄罗斯西伯利亚杂花、金皇后、阿尔冈金之间的亲缘关系远近和遗传多样性,确定不同品系之间的遗传差异及遗传背景,为验育种工作提供分子水平的理论依据和技术支持。
1 材料和方法
1.1 试验地概况
试验于2013年4月~2015年5月在高寒湿润气候区的甘南州夏河县牧草试验站进行。夏河县位于甘南藏族自治州西北部,E 101°54′~103°25′,N 34°32′~35°34′,地处青藏高原东北边缘,地势由西北向东南倾斜,海拔3 000~3 800 m,属于寒冷湿润性气候,土壤类型为高寒草甸土,有明显的腐殖质积聚,腐殖质层厚10 cm,呈灰棕至黑褐色粒状-扁核状结构,土壤有机质含量45 140 mg/kg,pH 7.87,速效氮50.83 mg/kg,速效磷10.57 mg/kg,速效钾230.0 mg/kg,土壤肥力均匀。
1.2 试验材料
供试材料GNKH-1(Cold resistance No.1 of alfalfa)、GNKH-2(Cold resistance No.2 of alfalfa)和GNKH-3(Cold resistance No.3 of alfalfa 3)为甘肃农业大学草业学院选育的3个抗寒苜蓿新品系,具有较强的抗寒能力和优质高产性能。对照材料为抗寒能力强的俄罗斯西伯利亚杂花苜蓿(M.sativasubsp.varia)(FD.2)、金皇后苜蓿(M.sativacv.Golden Empress)(FD.3)和阿尔冈金紫花苜蓿(M.sativacv.Algonguin)(FD.3),均由甘肃农业大学草业生态系统教育部重点实验室提供(表1)。
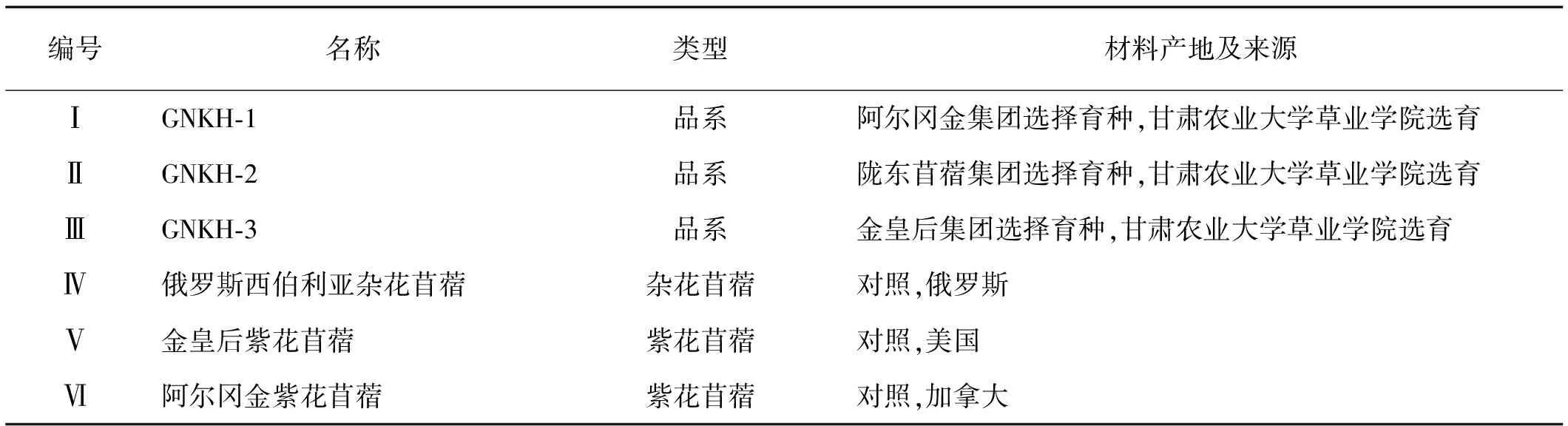
表1 供试苜蓿材料
1.3 试验设计
田间试验采用随机区组设计,小区面积3 m×5 m,3次重复,采用深开沟,浅覆土,冬前耙平的播种技术:人工开沟条播,沟深12~15 cm,覆土1~2 cm,冬前耙平埋颈,行距30 cm,播量1.7 g/m2。适时对试验地进行锄草及病虫害防治等田间管理。
于2013年12月在实验室内沙培种植,放置于光照充足、温度适宜的组织培养室内,期间定期定量加入Hogland营养液。待出苗后30 d采集各供试苜蓿材料幼嫩叶片提取DNA待用。
1.4 测定项目及方法
1.4.1 生产性能 (1) 物候期特征比较 对每个品种生长第1年和第2年的出苗期(返青期)、分枝期、现蕾期、初花期进行观测记录。苜蓿品种每小区留2行进行物候期观测,鉴别的标准是50%的植株达到某一个生育阶段为某生育期;10%~20%的植株达到为初期,70%~80%达到为盛期[18]。
(2) 越冬率 采用随机取样法进行调查,避开边缘地段,在小区株行内随机选取长为1 m的样段,调查每一样段内的株数。翌年植物返青后调查原样段内返青的株数,然后计数返青后存活的株数和越冬前总株数,用下式计算越冬率。

(3) 干鲜比和干草产量 干鲜比测产时,每个小区称鲜草500 g,自然风干后称重,计算干鲜比,干鲜比=苜蓿风干重/苜蓿鲜重;干草产量 在每茬初花期,每小区随机取样1 m2,重复3次,刈割后称重量,留茬高度3~5 cm,取平均值为鲜草产量,根据干鲜比计算干草产量。青藏高原高寒湿润气候区2013年刈割1茬,2014年刈割2茬,年度产量为各茬草产量之和。
(4) 株高和生长速度 株高 在第1茬初花期测产前,每个小区随机选取10株,测量植株绝对高度(拉直测定);生长速度 第1茬草从分枝期开始,每个小区随机选取10个单株,做好标记,直到初花期测定生长高度,两次生长高度之差除以生长天数即为生长速度。
(5)分枝数和叶茎比 分枝数 魏臻武等[35]的方法,每年第1茬初花期测定地表根茎部形成的分枝数。
叶茎比 测产时,取500 g全株鲜草将茎叶分离,称量叶片和茎秆鲜重,105℃杀青15 min,并置于65℃下烘至恒重,冷却后取出用1%天平称量叶片和茎秆干重。
1.4.2 遗传特征 (1) DNA提取 每份材料随机取10个单株的幼嫩叶片0.5 g,洗净叶片后等量混合,采用柱式植物基因组抽提试剂盒提取苜蓿总DNA。
(2) DNA检测 在浓度2.0%的琼脂糖凝胶上对提取的DNA进行电泳,于紫外凝胶成像仪中拍照;用紫外分光光度计于D260 nm与D280 nm检测所提取的DNA浓度。
(3) 引物合成与筛选 试验初期选择了10条引物,由上海生工有限公司合成。经引物筛选试验,筛选了PCR扩增效果好、条带清晰的8条引物用于6份苜蓿的遗传多样性研究。
(4)ISSR扩增与检测 采用25 μLPCR反应体系,模板DNA 2 μL、ISSR合成引物1 μL,DreamTaqGreen PCR Master Mix 12.5 μL,双蒸水9.5 μL。扩增条件为预变性94℃ 4 min,然后循环,94 ℃ 1 min,45~60℃ 40 s,72℃ 1~2 min,共40轮循环。循环结束后,72℃ 10 min,最后在4℃条件下保存。取PCR产物7 μL于1.6%的琼脂糖胶上电泳,在0.5×TBE缓冲液中电泳约1.5 h,在紫外凝胶成像仪中拍照。
1.5 数据处理和评价方法
1.5.1 生产性能评价 用SPSS 19.0统计软件和EXCEL进行数据处理及分析;应用主成分分析法对多个指标进行综合评判。先把参与主成分分析的n个指标变量进行标准化,计算出特征值和贡献率以及主成分,最后比较总得分值F的大小,F值越大表明该苜蓿材料生产性能适应性越强。
1.5.2 遗传特征分析 对强带以及清晰可辨的弱带进行统计赋值,采取0/1赋值的记带方法,即有带计为“1”,无带计为“0”,从而实现PCR扩增产物的条带总数和多态性条带数的统计。
多态性位点比率:p=i/j×100%
式中:i为多态性位点的数目,j为统计出的总位点数目。
使用软件Ntsys-pc 2.1软件进行分析,计算 Nei’s遗传距离,使用UPMGA法进行聚类分析,建立聚类图。使用软件 POPGEN 32计算ISSR扩增产物多态信息含量。
2 结果与分析
2.1 生物学特性及生产性能比较
2.1.1 物候期比较 高寒湿润气候区各苜蓿材料物候期差异很大,各苜蓿材料于2013年5月17日播种,5月25日出苗,截止6月1日齐苗,7月初开始分枝,抗寒苜蓿品系分枝期比对照苜蓿品种早10 d。播种第1年,GNKH-2和GNKH-3出苗期最早,比对照材料提前5~10 d;GNKH-3、GNKH-2有现蕾并有开花,但开花数不足10%,其他品种均只生长到分枝期。播种第2年,3个抗寒苜蓿品系返青、分枝、现蕾、开花、结荚、枯黄各物候期时间较为一致,对照品种俄罗斯西伯利亚杂花、金皇后和阿尔冈金各物候期时间较为一致,但GNKH-3和GNKH-2较其他材料提前5 d进入现蕾期。播种第2年,各品种均能结荚,种子均未成熟(表1)。

表2 高寒湿润气候区供试苜蓿材料的物候期
2.1.2 生产性能指标特征分析 播种当年,供试苜蓿材料均能安全越冬,但越冬率较低,苜蓿材料平均越冬率为60.32%,其中GNKH-3和GNKH-2的越冬率最高,达到了76.66%,72.34%,显著高于其他苜蓿品种(P<0.05),GNKH-1次之,阿尔冈金最低(40.39%)。播种的第2年,供试苜蓿材料的越冬率均超过90%,显著高于播种当年的越冬率(P<0.05),GNKH-3的越冬率也最高,为94.43%,显著高于对照苜蓿俄罗斯西伯利亚杂花苜蓿(P<0.05),GNKH-2次之,为93.21%。GNKH-3、GNKH-2、GNKH-1两年的平均越冬率分别为85.55%、82.78%、78.99%,均高于对照(图1)。
综合分析供试苜蓿材料在高寒湿润气候区的越冬率表现,GNKH-3为适宜高寒湿润气候区的越冬率最高的苜蓿材料,GNKH-2和GNKH-1为高寒湿润气候区越冬率较高的苜蓿材料。
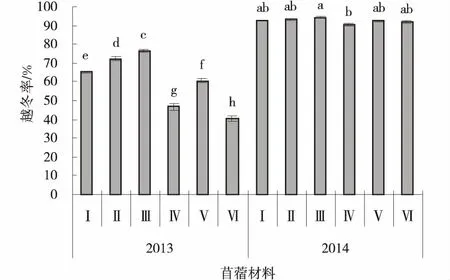
图1 越冬率Fig.1 Wintering rate
供试苜蓿材料干鲜比随着生长年限的增加均呈减小趋势。播种当年平均干鲜比为0.28,其中阿尔冈金干鲜比最高(0.29),俄罗斯西伯利亚杂花最低(0.26)。播种第2年,供试苜蓿材料的干鲜比均为0.25,其中GNKH-2干鲜比最高,为0.27,显著高于俄罗斯西伯利亚杂花和阿尔冈金(P<0.05),俄罗斯西伯利亚杂花也最低(0.23)。GNKH-2、GNKH-1两年的平均干鲜比分别为0.27、0.27,均高于对照(图2)。
综合分析供试苜蓿材料在高寒湿润气候区的干鲜比表现,GNKH-2为适宜高寒湿润气候区的干鲜比最高的苜蓿材料,GNKH-1为高寒湿润气候区干鲜比较高的苜蓿材料。
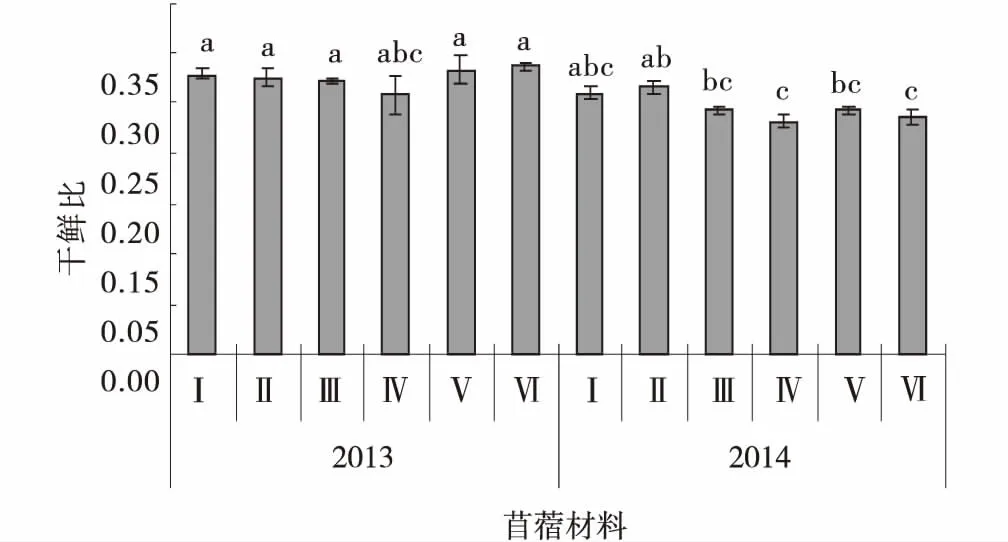
图2 干鲜比Fig.2 Ratio of dry to fresh mater
播种当年,供试苜蓿材料由于生育期没有达到初花期,干草产量较低,随着生长年限的增加,不同苜蓿材料的干草产量均呈增加趋势。播种当年平均干草产量为2.20 t/hm2,其中GNKH-2干草产量最高,为2.95 t/hm2,显著高于对照俄罗斯杂花和金皇后(P<0.05),GNKH-1和GNKH-3次之,分别为2.77、2.67 t/hm2,俄罗斯西伯利亚杂花最低,为1.38 t/hm2。播种第2年,抗寒苜蓿材料的干草产量显著高于播种当年(P<0.05),对照苜蓿材料的干草产量相对于播种当年有所增加,但是不显著。供试苜蓿材料的干草产量平均为5.15 t/hm2,其中GNKH-3干草产量最高,为8.81 t/hm2,显著高于GNKH-1和对照苜蓿材料(P<0.05),GNKH-2和GNKH-1次之,也显著高于对照苜蓿材料(P<0.05),金皇后最低(1.94 t/hm2)。GNKH-3,GNKH-2和GNKH-1两年的平均干草产量分别为5.74、5.49和4.94 t/hm2,均高于对照(图3)。
综合分析供试苜蓿材料在高寒湿润气候区的干草产量表现,GNKH-3为适宜高寒湿润气候区的干草产量最高的苜蓿材料,GNKH-2和GNKH-1为高寒湿润气候区干草产量较高的苜蓿材料。
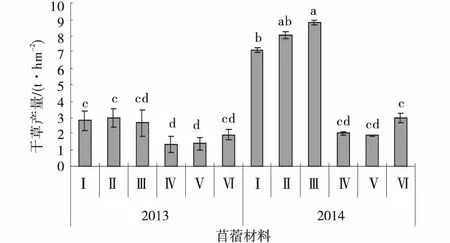
图3 干草产量Fig.3 Hay yield
(3) 株高和生长速度 播种当年,供试苜蓿材料的平均株高为48.10 cm,其中GNKH-3、GNKH-2株高最高,分别为53.45 cm、52.42 cm,显著高于对照材料俄罗斯西伯利亚杂花和金皇后(P<0.05),金皇后最低(41.33 cm)。播种第2年,供试苜蓿材料的株高显著高于播种当年(P<0.05),平均株高为64.66 cm,其中GNKH-3株高最高,为70.62 cm,显著高于对照苜蓿材料(P<0.05),GNKH-2、GNKH-1次之,为68.32、65.57 cm,均高于对照材料(57.49~64.21 cm)。GNKH-3,GNKH-2和GNKH-1两年的平均株高分别为62.03,60.37和58.56 cm,均高于对照(图4)。
供试苜蓿材料在高寒湿润气候区生长缓慢,生长速度低,播种当年只能收获1茬,第2年只能收获2茬。播种当年,供试苜蓿材料的生长速度平均值为0.65 cm/d,其中GNKH-3、GNKH-2生长速度显著高于对照俄罗斯杂花和金皇后(P<0.05),分别为0.74、0.73 cm/d,GNKH-1次之,为0.70 cm/d,金皇后最低(0.55 cm/d)。播种第2年,供试苜蓿材料的平均生长速度为0.77 cm/d,供试苜蓿材料的生长速度有所增加,其中GNKH-3、GNKH-2生长速度也显著高于对照俄罗斯杂花和金皇后,分别为0.86、0.83 cm/d,GNKH-1次之,为0.82 cm/d,金皇后最低(0.66 cm/d)。GNKH-3、GNKH-2、GNKH-1两年的平均生长速度分别为0.80,0.78和0.76 cm/d,均高于对照(图5)。
综合分析供试苜蓿材料在高寒湿润气候区的株高和生长速度表现,GNKH-3为高寒湿润气候区的株高和生长速度最高的苜蓿材料,GNKH-2和GNKH-1为高寒湿润气候区株高和生长速度均较高的苜蓿材料。

图4 株高Fig.4 Plant height

图5 生长速度Fig.5 Growth rate
(4) 分枝数和叶茎比 播种当年,供试苜蓿材料的分枝数平均为11.0个/株,其中GNKH-2分枝数显著高于俄罗斯杂花和金皇后(P<0.05),为12.7个/株、GNKH-1、GNKH-3次之,分别为12.2、12.1个/株,金皇后最少(8.7个/株)。播种第2年,供试苜蓿材料的平均分枝数为14.3个/株,抗寒苜蓿材料的分枝数显著高于播种当年(P<0.05),对照苜蓿材料的分枝数相对播种当年也有所增加,其中GNKH-3、GNKH-2、GNKH-1分枝数显著多于对照苜蓿材料(P<0.05),分别为18.5,17.8和17.1个/株。GNKH-3、GNKH-2、GNKH-1两年的平均分枝数分别为15.3,15.2和14.7个/株,均高于对照(图6)。
播种当年,供试苜蓿材料的叶茎比平均为1.11,其中GNKH-3,GNKH-2和GNKH-1叶茎比均比对照多,分别为1.15,1.14和1.13,金皇后最小(1.01),显著低于其他苜蓿材料(P<0.05)。播种第2年,供试苜蓿材料的叶茎比显著低于播种当年(P<0.05),平均叶茎比为0.84,其中GNKH-3,GNKH-1和GNKH-2叶茎比均大于对照苜蓿材料,分别为0.89,0.88和0.86。GNKH-3,GNKH-1和GNKH-2两年的平均叶茎比分别为1.02,1.01和1.00,均高于对照(图7)。
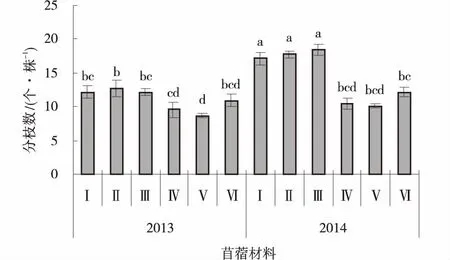
图6 分枝数Fig.6 Branching number
综合分析供试苜蓿材料在两个气候区的分枝数和叶茎比表现,GNKH-3为高寒湿润气候区的分枝数和叶茎比最高的苜蓿材料,GNKH-1、GNKH-2为高寒湿润气候区的分枝数和叶茎比较好的苜蓿材料。

图7 叶茎比Fig.7 Leaf to stem ratio
2.2 遗传多样性分析
2.2.1 DNA 检测 分子标记是以DNA多态性为基础的遗传标记,在苜蓿生物技术育种和种质资源评价中的地位不可忽视。因此,在苜蓿常规育种的基础上,结合分子标记辅助育种,进行种质资源评价,可为我国苜蓿种质资源的科学利用和育种工作提供更为可靠的技术支持。简单重复序列间扩增分子标记技术能够更好的检测出生物遗传物质的多态性,因此,在遗传上有远景[13],ISSR标记为显性标记,可以揭示整个基因组的一些特征,符合孟德尔遗传规律,具有稳定性好,多态性高,试验操作简单、快速、用时少等优点,该技术被广泛用作研究植物种质资源遗传多样性及亲缘关系的有效手段[14-15]。
挑选每个材料条带清晰、无降解、无拖尾现象的DNA样品进行检测。6 份参试材料DNA条带清晰,点样孔干净,表明DNA片段大小比较均一,纯度比较高,总DNA稍带乳白色,在D260 nm及D280 nm处的D值为 1.80~1.88,浓度2.0%的琼脂糖凝胶电泳检测结果比较理想,能够达到ISSR分子标记分析的要求(图8)。

图8 6份苜蓿材料的DNA检测Fig.8 DNA testing chart of 6 alfalfa materials注:从左到右依次为M,GNKH-1,GNKH-2,GNKH-3,俄罗斯西伯利亚杂花苜蓿,金皇后苜蓿,阿尔冈金苜蓿
2.2.2 PCR 扩增产物多态性 用通过引物筛选试验得到的8条多态性好的引物,对6份苜蓿材料进行扩增,并对扩增图谱进行统计分析。结果表明8个引物共扩增出87条清晰谱带,平均每个引物能扩增出10.9个条带,平均多态性比率(PPB)为62.07%。不同引物扩增出来的片段数不同,除了引物844和847的条带较少,分别为7个和9个;引物822,845和888的条带数均大于10个,他们的扩增产物多态性最丰富,图谱也很清晰,并且扩增产物的多态性比率均在80%,多态性信息含量较高。其他3个引物825,855和885的条带数也均大于10个,他们的扩增产物多态性也较丰富,图谱也清晰。6份材料的有效等位基因数为1.494 3、基因多样性指数为0.2676和Shannon’s信息指数为0.3842(表3)。
2.2.3 遗传相似性系数分析 6份材料间的遗传相似性系数在0.482 8~0.954 0,GNKH-1和GNKH-2的遗传相似性最大,为0.954 0,GNKH-1与亲本阿尔冈金的遗传相似性为0.517 2;GNKH-3与阿尔冈金间的遗传相似性最小,为0.482 8,说明它们在亲缘关系上离得最远,GNKH-3与亲本金皇后的遗传相似性为0.551 7;与GNKH-3遗传相似性最高的是俄罗斯西伯利亚杂花,为0.6667。3个抗寒苜蓿品系与对照材料的遗传相似性都比较小,为0.482 8~0.666 7,表明抗寒苜蓿品系与对照材料的亲缘关系相对较远,具有相对独立的遗传特性。整体分析,6份苜蓿材料间遗传异质性丰富(表4)。

表3 ISSR分析所用引物序列和扩增结果
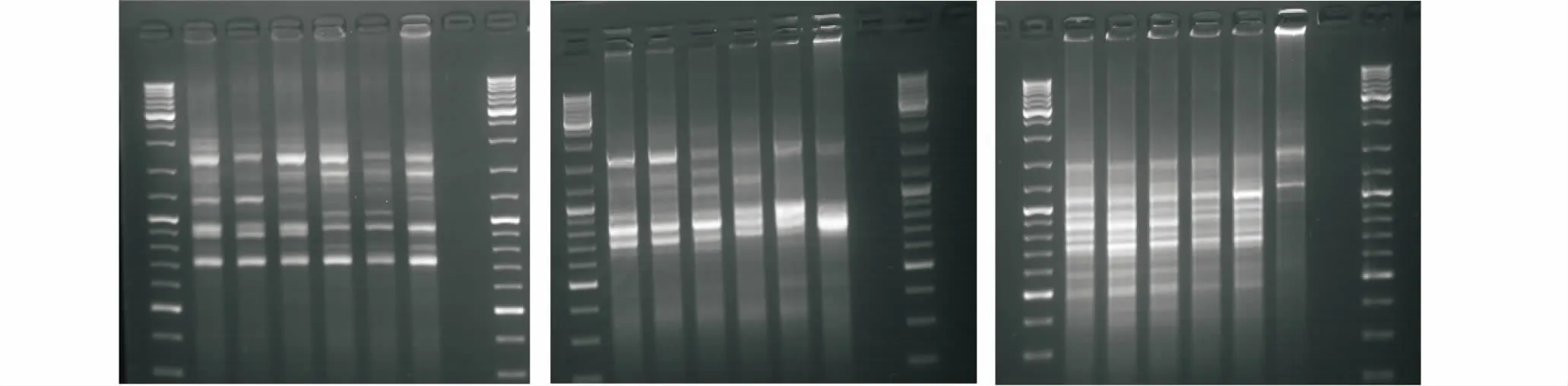
图9 引物822,845和888 ISSR扩增Fig.9 Amplification map of 822,845,888 ISSR注:从左到右依次为M,GNKH-1,GNKH-2和GNKH-3,俄罗斯西伯利亚杂花苜蓿,金皇后苜蓿,阿尔冈金苜蓿
2.2.4 ISSR遗传多样性分析 利用ISSR遗传相似性系数矩阵,根据UPGMA法进行聚类分析,构建了6份参试苜蓿材料的亲缘关系系统树状图。在遗传距离0.78处,6份苜蓿材料被分为2个大类,第1个大类包括了GNKH-1,GNKH-2和GNKH-3抗寒苜蓿品系,第2大类包括俄罗斯西伯利亚杂花,金皇后和阿尔冈金。在遗传距离0.86处,6份苜蓿材料被分为3个大类,第1个大类包括了GNKH-1,GNKH-2和GNKH-3抗寒苜蓿品系,第2大类包括俄罗斯西伯利亚杂花和金皇后,第3大类只有阿尔冈金。在遗传距离0.88处,6份苜蓿材料被分为4个大类,第1个大类包括了GNKH-1、GNKH-2、GNKH-3 抗寒苜蓿品系,第2大类只有俄罗斯西伯利亚杂花,第3大类只有金皇后,第4大类只有阿尔冈金。在遗传距离0.90处,GNKH-3单独聚类(图10)。
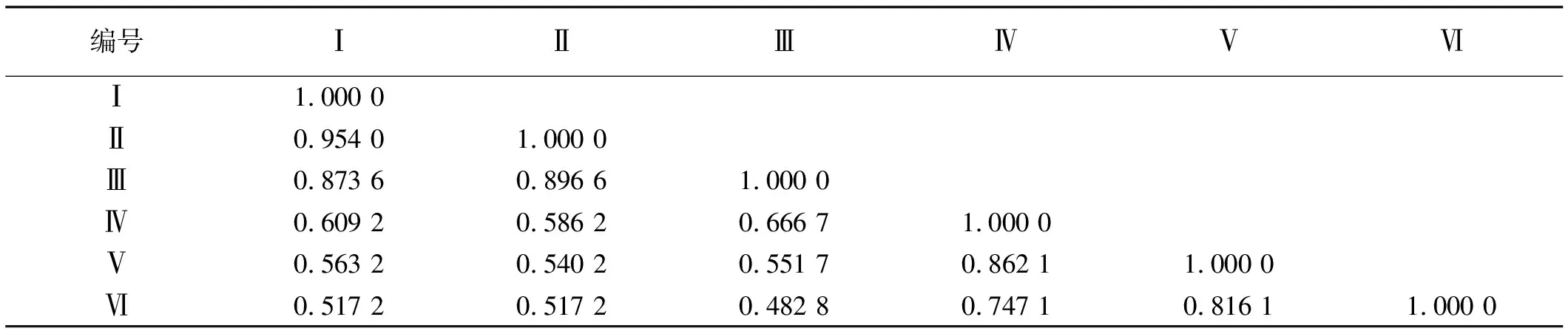
表4 6份苜蓿材料遗传相似系数矩阵

图10 6份苜蓿材料基于ISSR的遗传相似性UPGMA聚类Fig.10 UPGMA cluster analysis based on ISSR genetic identities among 6 alfalfa materials注:1-GNKH-1,2-GNKH-2,3-GNKH-3,4-俄罗斯西伯利亚杂花苜蓿,5-金皇后苜蓿,6-阿尔冈金苜蓿
2.3 生产性能综合评价
由于供试苜蓿材料的来源不同、基因型不同,其在形态特征和产量及品质指标上变化非常大,很难用一个或几个指标对其作出准确的评价。因此,选择影响生产性能的主要因子越冬率、干草产量、株高、生长速度、分枝数、叶茎比,采用主成分分析法对6份苜蓿种质材料连续两年的生产性能进行综合评价。
根据综合评价函数,计算各种质材料的综合得分(表5),给予供试苜蓿材料定量化的描述,综合得分越大,表明综合表现越好。高寒湿润气候区各苜蓿材料主成分分析综合得分值排名为:GNKH-3>GNKH-2>GNKH-1>阿尔冈金>俄罗斯西伯利亚杂花>金皇后。GNKH-3的综合得分最高,在高寒湿润气候区的适应能力最强;其次是GNKH-2和GNKH-1,它们在高寒湿润气候区的适应能力较强;俄罗斯西伯利亚杂花和金皇后的综合得分最低,它们在高寒湿润气候区的适应能力最弱(表5)。
3 讨论
3.1 物候期及农艺性状差异
植株整个生育期的生长是植物自身对外界生态因子适应性的反应。供试苜蓿材料在高寒湿润气候区生育期差异显著,GNKH-3、GNKH-2返青期最早,阿尔冈金返青期最迟,比抗寒苜蓿品系晚5 d。3个抗寒苜蓿品系的物候期与畅云喜等[16]在青藏高原观测的抗寒苜蓿新品系的物候期差别不大。

表5 在高寒湿润气候区供试苜蓿材料综合得分及排名
潘正武等[17]于2009~2011年在天祝高寒湿润气候区和西藏“一江两区”进行品种比较试验,挑选出了适应性较高的抗寒苜蓿品种苜蓿王等[18]。陈玲玲等[19-27]对国外苜蓿的研究表明,其物候期、株高、产量、分枝数、叶茎比等农艺性状的差异较大,这与青藏高原的高寒环境有着直接的关系,由于海拔高、气候多变、温差较大、降水时间不均匀,适宜苜蓿生长的时间相对较短(5~8月),致使苜蓿生长缓慢,无法完成整个生育期。越冬率是最重要的一个抗寒生理指标,其大小可以反映苜蓿品种的抗寒能力。在低温胁迫下,植株的越冬率越高,抗寒性越强,越能适应高寒气候。试验所用的抗寒苜蓿品系在高寒湿润气候区两年平均越冬率比对照材料高,其中,GNKH-3、GNKH-2越冬率均超过80%。高寒湿润气候区两年的平均干草产量均是抗寒苜蓿品系显著高于对照苜蓿材料(P<0.05),其中GNKH-3最高,GNKH-2、GNKH-1次之。有研究表明植株高度是衡量牧草生长状况的重要指标,与产量呈正相关,高植株通常有更高的相对产量潜力[21-22],这与试验结果相一致;高寒湿润气候区3个抗寒苜蓿品系株高比对照的3个苜蓿材料高,他们之间差异不显著。生长速度在一定程度上反映苜蓿生长能力的强弱[23],高寒湿润气候区两年的平均生长速度为抗寒苜蓿品系高于对照苜蓿材料。牧草分枝(分蘖)数的多少在一定程度上影响其生产性能的高低[24],苜蓿由于分枝数的增多,干物质产量也随之增加,高寒湿润气候区两年的平均分枝数抗寒苜蓿品系多于对照苜蓿材料,因此,干草产量也较高。供试材料高寒湿润气候区两年的平均叶茎比以GNKH-3最高,达到了1.02。
3.2 ISSR分子标记的检测效率及材料的遗传多样性分析
分子标记己经成为苜蓿育种中的重要辅助选择手段。用8对引物在供试苜蓿材料中共获得87条扩增带,其中54条具有多态性,引物的多态位点百分率62.07%。李红等[15]用10个引物对30份苜蓿材料共扩增出112条带,其中59条带是多态性条带,多态性比率的平均值为74.5%。李哲等[16-17]用4条引物对39份苜蓿材料共扩增22条带,其中21条为多态性条带,多态性比率为95.45%。张颖娟等[25]用13条引物对12份苜蓿材料共扩增出346条带,多态性条带332条,多态性条带百分比为95.95%。扩增产物的多态性,能反映材料的遗传多样性[26],以此为依据,供试苜蓿种质资源具有较丰富的遗传多样性。与多态位点百分率相比,基于Nei’s基因多样性指数和基于条带表型频率的Shannon’s信息指数更能客观地衡量材料的遗传多样性水平[27],研究的Nei’s基因多样性指数和Shannon’s信息指数分别为1.494 3和0.384 2,进一步表明供试苜蓿种质资源具有一定水平的遗传多样性。
3.3 供试材料间的亲缘关系分析
遗传相似系数是反映个体之间遗传相似程度和遗传背景一致性的度量值。3个抗寒苜蓿品系间的遗传相似性为0.873 7~0.954 0,对照材料间的遗传相似性为0.747 1~0.862 1,材料间的遗传相似程度较高,亲缘关系较近。此次研究亲本材料阿尔冈金、陇东苜蓿、金皇后苜蓿在海拔3 050 m的青藏高原连续种植很多年,抗寒性能得到长期驯化,在选育过程中移植到暖温地带,又移植到青藏高原高寒地区,进行多带驯化,抗寒能力得到提高,选育的抗寒苜蓿新品系与亲本材料形成明显的差异,GNKH-1与亲本阿尔冈金的遗传相似性为0.517 2;GNKH-3与亲本金皇后的遗传相似性为0.5517,遗传距离增大,遗传多样性丰富。GNKH-3与阿尔冈金间的遗传相似性最小,为0.482 8,说明它们在亲缘关系上离得最远,3个抗寒苜蓿品系与对照材料的遗传相似性都比较小,介于0.482 8~0.666 7,表明抗寒苜蓿品系与对照材料的亲缘关系相对较远,具有相对独立的遗传特性。
根据6份供试苜蓿材料的亲缘关系系统树,在遗传距离0.78处将6个苜蓿供试材料分为2类。第1类包含GNKH-1,GNKH-2和GNKH-3 3个抗寒苜蓿品系材料,具有高产、越冬率高、生产性能强的特点;第二类包含俄罗斯西伯利亚杂花、金皇后、阿尔冈金苜蓿3个对照品种材料,其产量、越冬率、生产性能相对第1类材料较低。在遗传距离0.90处,GNKH-3单独聚类,为青藏高原地区抗寒苜蓿品种的选育提供了理论基础。GNKH-1和GNKH-2各方面表现仅次于GNKH-3。因此,在常规育种实践中,采用集团选择法在原有种质资源中发掘新的、有变异的种质资源及遗传特性相对独立的材料具有重要意义。
青藏高原缺乏适应性能好、高产优质的苜蓿品种,GNKH -3,GNKH-1和GNKH-2的选育为在东北缘区域实现苜蓿的产业化提供了物质基础,有望解决青藏高原东北缘区豆科饲草缺乏的问题,实现高寒牧区家畜的营养平衡和健康养殖。
4 结论
(1)高寒湿润气候区各苜蓿材料主成分分析综合得分值排名为:GNKH-3>GNKH-2>GNKH-1>阿尔冈金>俄罗斯西伯利亚杂花>金皇后。GNKH-3的综合得分最高,在高寒湿润气候区的适应能力最强;其次是GNKH-2和GNKH-1。
(2)在遗传距离 0.78处将6个供试苜蓿材料分为2类。第1类包含GNKH-1,GNKH-2和GNKH-3抗寒苜蓿品系材料,在高寒湿润气候区具有高产、越冬率高、生产性能强的特点;第2类包含俄罗斯西伯利亚杂花、金皇后、阿尔冈金苜蓿3个对照品种材料,其产量、越冬率、生产性能相对第1类材料较低。在遗传距离0.90处,GNKH-3单独聚类。GNKH-3为高寒湿润气候区最理想的优良苜蓿品系,GNKH-2次之。
猜你喜欢
蔬菜(2022年10期)2022-11-10
今日农业(2021年20期)2021-11-26
现代畜牧科技(2021年4期)2021-07-21
河北果树(2020年4期)2020-11-26
河北果树(2020年1期)2020-02-09
园林科技(2020年2期)2020-01-18
山东林业科技(2019年2期)2019-06-03
云南农业科技(2019年2期)2019-04-04
作文小学高年级(2019年1期)2019-02-18
中国三峡(2017年4期)2017-06-06
