
马氏珠母贝β2肾上腺素能受体(β2AR)分子特征和表达分析
2019-07-17 01:10:26张雨婷曾嫚玉马佳仪李嘉欣杜晓东
广东海洋大学学报 2019年4期
张雨婷,曾嫚玉,马佳仪,李嘉欣,焦 钰,,杜晓东
马氏珠母贝肾上腺素能受体()分子特征和表达分析
张雨婷1,曾嫚玉1,马佳仪1,李嘉欣1,焦 钰1,2,杜晓东2
(1. 广东海洋大学水产学院 / 2. 广东省珍珠养殖与加工工程技术研究中心,广东 湛江 524088)
【目的】研究马氏珠母贝()肾上腺素能受体()的生物学功能。【方法】利用cDNA末端快速扩增技术(RACE)克隆获得基因的cDNA的全长序列并对其序列特征进行分析,实时荧光定量技术(qRT-PCR)检测马氏珠母贝不同组织中基因mRNA的表达水平。【结果与结论】的cDNA全长2 426 bp,包括开放阅读框(ORF)2 091 bp,编码696个氨基酸,5′UTR长144 bp,3′UTR长191 bp,预测分子质量79.0 ku,理论等电点9.18,多重序列比对结果发现在不同物种间的保守性较高。软件分析结果显示的氨基酸序列具有7个典型的跨膜结构域。qRT-PCR结果表明,基因在各组织中均有表达,在鳃中表达量最高,在肝胰腺和性腺中表达量也较高。
马氏珠母贝;肾上腺素能受体;分子特征;表达分析
肾上腺素能受体(beta 2 adrenergic receptor,)属于G蛋白偶联受体(G Protein-Coupled Receptors,GPCR)家族,在体内可被肾上腺素激活[1]。参与调控免疫细胞[2-3],以及胶质细胞、成纤维细胞、上皮细胞等的调控[4-6]。肾上腺素能受体有3个亚型,分别是、、,在同种动物体内这3种受体的氨基酸同源性接近50%,同一种受体的同源性在不同种类动物之间可达75%及以上[7]。含有7个跨膜α螺旋结构域,有3个细胞外环,其中一个是氨基末端,3个细胞内环,有羧基末端[8]。
调节免疫反应是通过G蛋白依赖的G蛋白偶联受体激酶2信号通路(G-protein-coupled receptor kinases,GRK2)和抑制蛋白2(β-arrestin2)信号通路。被相应的配体激活后被GRK2磷酸化,后β-arrestin 2移动至细胞膜并结合磷酸化的受体,使该受体与G蛋白结合,可促进内化和脱敏[4]。在炎症反应细胞因子网络中发挥免疫抗炎功能,通过抑制肿瘤坏死因子α(tumor necrosis factor alpha,TNF-α)生成、阻止白细胞黏附和减少毛细血管通透性等[9]。研究表明,在神经系统对免疫的调控中,交感神经可以通过释放神经递质,激活细胞表面的肾上腺素能受体,通过介导神经递质发挥细胞免疫功能[2]。去甲肾上腺素刺激后单核或者巨噬细胞产生的抗炎因子增加、促炎因子减少[10]。关于的功能报道,在软体动物中仅见于缢蛏[11],在马氏珠母贝中尚未见相关报道。
马氏珠母贝(),属于软体动物门双壳纲,又称合浦珠母贝(),是海水珍珠养殖的重要经济物种,其生长环境的特殊性,使其易于被病原体和微生物侵染[12]。目前对贝类病害的相关研究尚不成熟,致使我国贝类养殖业发展缓慢,因此贝类免疫相关基因研究具有重要的现实意义。本研究利用cDNA末端快速扩增技术(RACE)克隆获得基因的cDNA的全长序列并对其序列特征进行分析;实时荧光定量技术(qRT-PCR)检测马氏珠母贝不同组织中基因mRNA的表达水平,分析马氏珠母贝关于免疫和应激反应的相关基因,了解其免疫机制,以期为提高马氏珠母贝抗病能力和降低插核后的免疫排斥提供理论支持,为进一步探究在贝类免疫机制中的生物学功能提供参考。
1 材料与方法
1.1 材料
1.1.1 实验材料 实验用贝为大小一致、生长状况良好的2龄马氏珠母贝,用于qRT-PCR的材料取自贝体的外套膜套膜区(MP)和中央膜(MC)、闭壳肌(A)、鳃(GI)、血细胞(B)、性腺(GO)、足(F)和肝胰腺(HE),RACE扩增所用材料为马氏珠母贝的性腺和鳃。样品取出后立即放于液氮中速冻,超低温冰箱-80 ℃保存。感受态细胞DH5α保存于-80 ℃冰箱。
1.1.2 实验试剂 Trizol Lipofectamine®2000 CD购买于Invitrogen公司,Reverse Transcriptase M-MLV、SMARTer RACE 5′/3′Kit、rTaq酶、PMD-18T Vector购买于TaKaRa公司,GeneJET PCR Purification Kit购买于Thermo Scientific公司。
1.1.3 引物设计 根据马氏珠母贝珍珠囊转录组文库中筛选出注释基因高度相似的unigene序列为模板,利用Primer Premier 5.0软件设计引物(表1)。
1.2 方法
1.2.1 总RNA 提取及cDNA第一链的合成 分离马氏珠母贝的各组织,在2.5 mL离心管中加入1 mL Trizol后,再加入约100 mg组织量,经组织研磨仪研磨后参照Trizol试剂说明书抽提马氏珠母贝各组织的总RNA。分别取各组织2 μL用于琼脂糖凝胶电泳和核酸定量仪[(260 nm)/(280 nm)比值]检测其完整性、浓度及纯度。根据Reverse Transcriptase M-MLV SMARTer RACE 5′/3′ Kit说明书,反转录合成cDNA第一链[13]。
1.2.2基因的克隆 使用巢式PCR依次进行中间片段、5′端、3′端的扩增。将产物纯化回收后连接至pMD18-T载体,再转入DH5α感受态细胞中,经细菌基础培养基LB培养后,挑取单一阳性克隆样品送至上海生工进行测序[14]。

表1 本实验所用引物
1.2.3 序列分析 利用DNAMAN软件将中间片段与5′端、3′端片段进行拼接,获得了全长cDNA。利用在线工具ORF Finder (http://www.ncbi.nlm.nih.gov/gorf/orfig.cgi)对基因进行开放阅读框(ORF)和氨基酸序列的预测;利用ExPASY(http://web.expasy.org/ protparam/)分析其理化性质;利用SignalP4.1(http://www.cbs.dtu.dk/services/Signal/)预测其是否有信号肽;经Motif(http://myhits.isb-sib.ch/cgi -bin/motif_scan)对其进行功能位点的预测;利用SMART(http://smart.embl-heidelberg.de/smart/)进行结构域预测;用SWISS-MODEL(http:// swissmodel.expasy.org/)预测的3级结构;采用MEGA7的 Neighbor-Joining 法构建进化树,检验 Bootstrap 值并设置抽样次数为 500 次;利用DNAMAN软件对马氏珠母贝与已知的不同动物氨基酸多序列进行比对[15]。
1.2.4 实时荧光定量检测组织差异表达 以为内参基因,各组织的反转录cDNA为模板,反应体系(10 μL):10 μmol/L的上、下游引物各 0.4 μL,cDNA 0.5 μL,SYBR®Select Master Mix 5 μL,灭菌的ddH2O 3.7 μL。反应条件:95 ℃下预变性 2 min,95 ℃下变性15 s,60 ℃下退火 1 min,40个循环,添加1个溶解曲线。每个样品进行重复 3 次进行。采用2-ΔΔCT分析法,样本重复性和组织间差异性利用SPSS19.0软件进行单因素方差分析,显著性水平设为0.05。
2 结果与分析
2.1 Pmβ2AR基因cDNA全长的克隆及序列分析
克隆得马氏珠母贝()894 bp 的5′ 端上游序列和415 bp 的3′ 端下游序列,拼接后得到2 426 bp的全长cDNA。其中5′UTR为144 bp,3′UTR为191 bp,包含3′末端25 bp的poly A尾巴。开放阅读框长2 091 bp,编码696个氨基酸(图1)。
2.2 Pmβ2AR氨基酸序列理化性质
预测分子质量约为69.0 ku,理论等电点为9.18。氨基酸序列中带负电荷氨基酸残基(Asp和Glu)共63个,正电荷数的氨基酸残基(Arg和Lys)共80个。预测此氨基酸不稳定指数(II)为46.35,将此蛋白质分类为不稳定蛋白;亲水指数为-0.398,为亲水蛋白(图2),N-端不存在信号肽;具有跨膜区域,表明该蛋白不是胞质蛋白。经预测,氨基酸序列在73~342位含有7个跨膜结构域,在371~440位含有两个低复杂度结构域,对虾夷扇贝进行结构域分析得知其具有相同的7个跨膜结构域(图3)。氨基酸序列中其他功能位点包括cAMP和cGMP依赖性蛋白激酶磷酸化位点5个,n-糖基化位点10个,酪氨酸激酶磷酸化位点1个等。经SOPMA 软件预测其二级结构,α螺旋为33.19%,延伸链(Ee)为17.96 %,β转角(Tt)为4.17%,无规则卷曲(Cc)为44.68 %。用SWISS-MODEL软件预测蛋白分子的三维结构,与虾夷扇贝有较高相似性(图4)。
2.3 Pmβ2AR的多序列比对和同源性分析
采用DNAMAN软件将与珍珠鸡(, XP_021266164.1)、墨西哥丽脂鲤(,XP_007232946.2)、热带爪蟾(,NP_001116897.2)和虾夷扇贝(,OWF49060.1)的氨基酸序列进行多序列比对分析,结果显示,与虾夷扇贝的相似性最高为70%(图5)。将的氨基酸序列用NCBI与其他物种进行对比,得到小鼠(,NP_031446.2)、马氏珠母贝、虾夷扇贝、地中海果蝇(,XP_023159092.1)、智人(,NP_000015.1)、斑马鱼(,NP_001082940.2)、亚洲水牛(,XP_006052758.1)、绿头鸭(,XP_005010059.3)、美洲牡蛎(,XP_022294171.1)、美洲鲎(,XP_013791385.1)、红鳍东方鲀(,XP_011609112.1)、多疣壁虎(,XP_015262106.1)、青鳉(,XP_015232667.1)、热带爪蟾和长牡蛎(,XP_011439690.1)的氨基酸序列,以NJ法构建系统进化树,结果显示,脊椎动物聚为一个分支;马氏珠母贝与虾夷扇贝聚为一簇,与虾夷扇贝的亲缘关系最近,结果与传统的分类相吻合(图6)。

5′ 和 3′ 非编码区用小写字母表示; 始密码子ATG和终止密码子TAG用方框标注; 双划线部分为7个跨膜结构域;阴影部分为2个低复杂区;虚线部分为cAMP和cGMP依赖性蛋白激酶磷酸化位点;单划线部分为n-糖基化位点;波浪线部分为酪氨酸激酶磷酸化位点
5′ UTR and 3′ UTR are indicated with small letters; Nucleotide with a frame represents the initiation codon (ATG) and stop codon (TGA); The double underline sequence represents 7 Transmembrane region; The shaded part represents 2 low complexity; The dashed underlined sequence represents cAMP and cGMP-dependent protein kinase phosphorylation sites; The single underlined sequence represents n-glycosylation sites; The wavy underlined sequence represents indicates tyrosine kinase phosphorylation sites
图1基因的核酸序列分析
Fig. 1 The nucleotide sequence analysis of
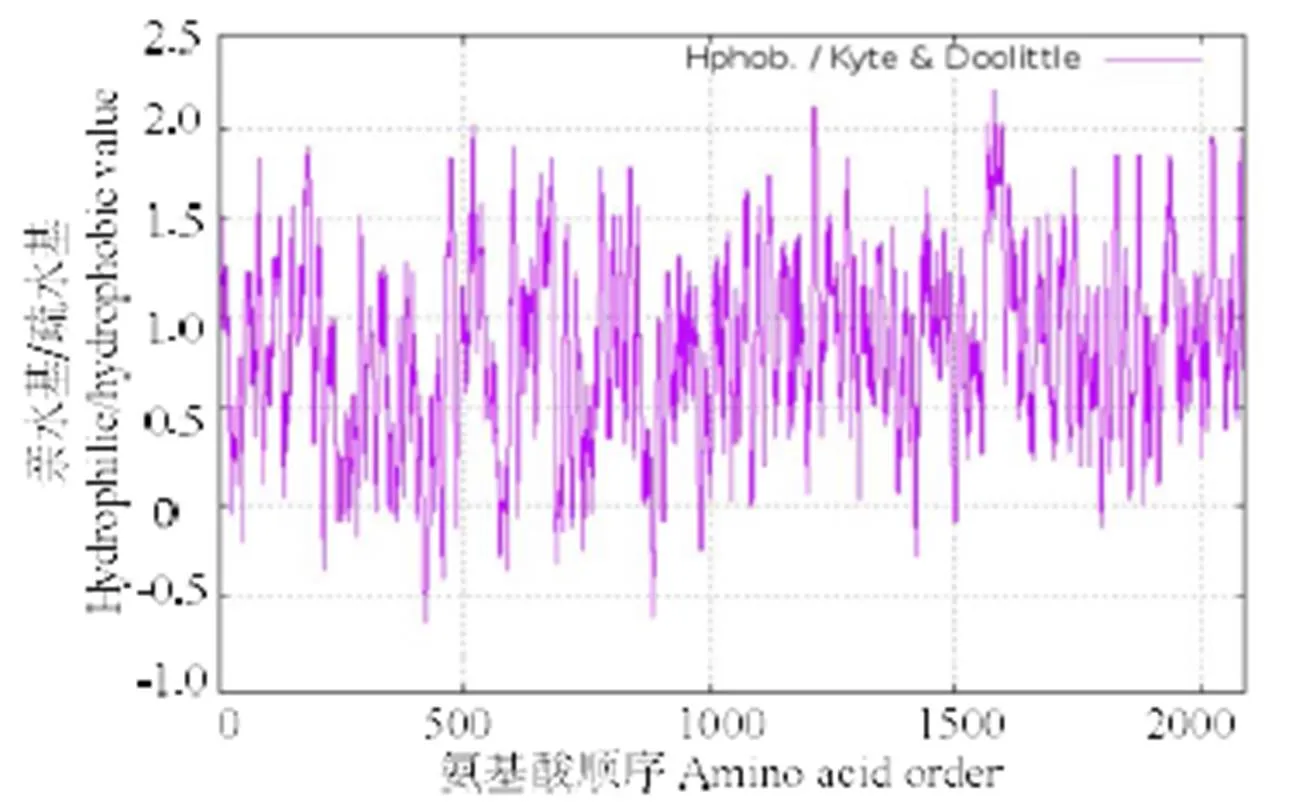
“+”: 疏水性 Indicates hydrophobicity; “-”: 亲水性 Indicates hydrophilicity
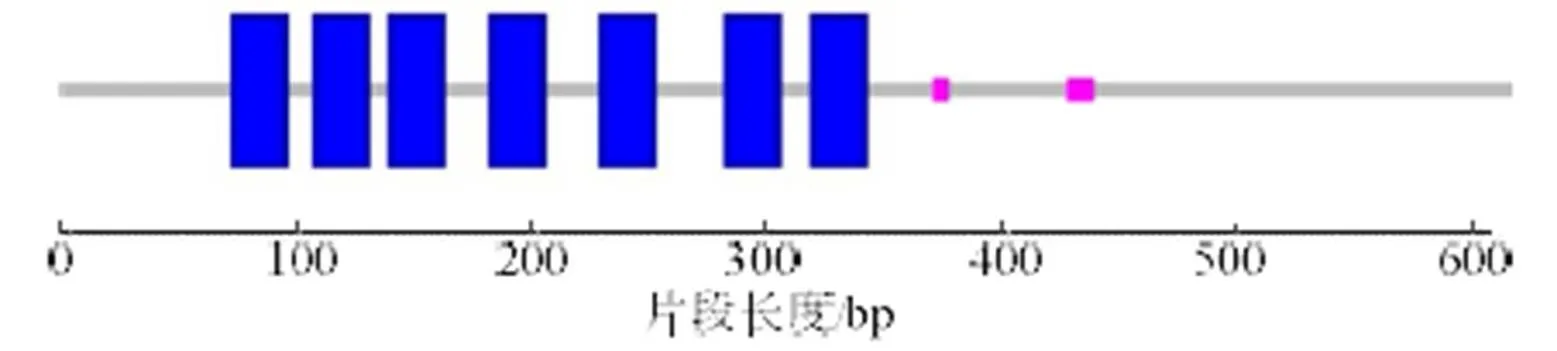
图3 Pmβ2AR蛋白质结构域
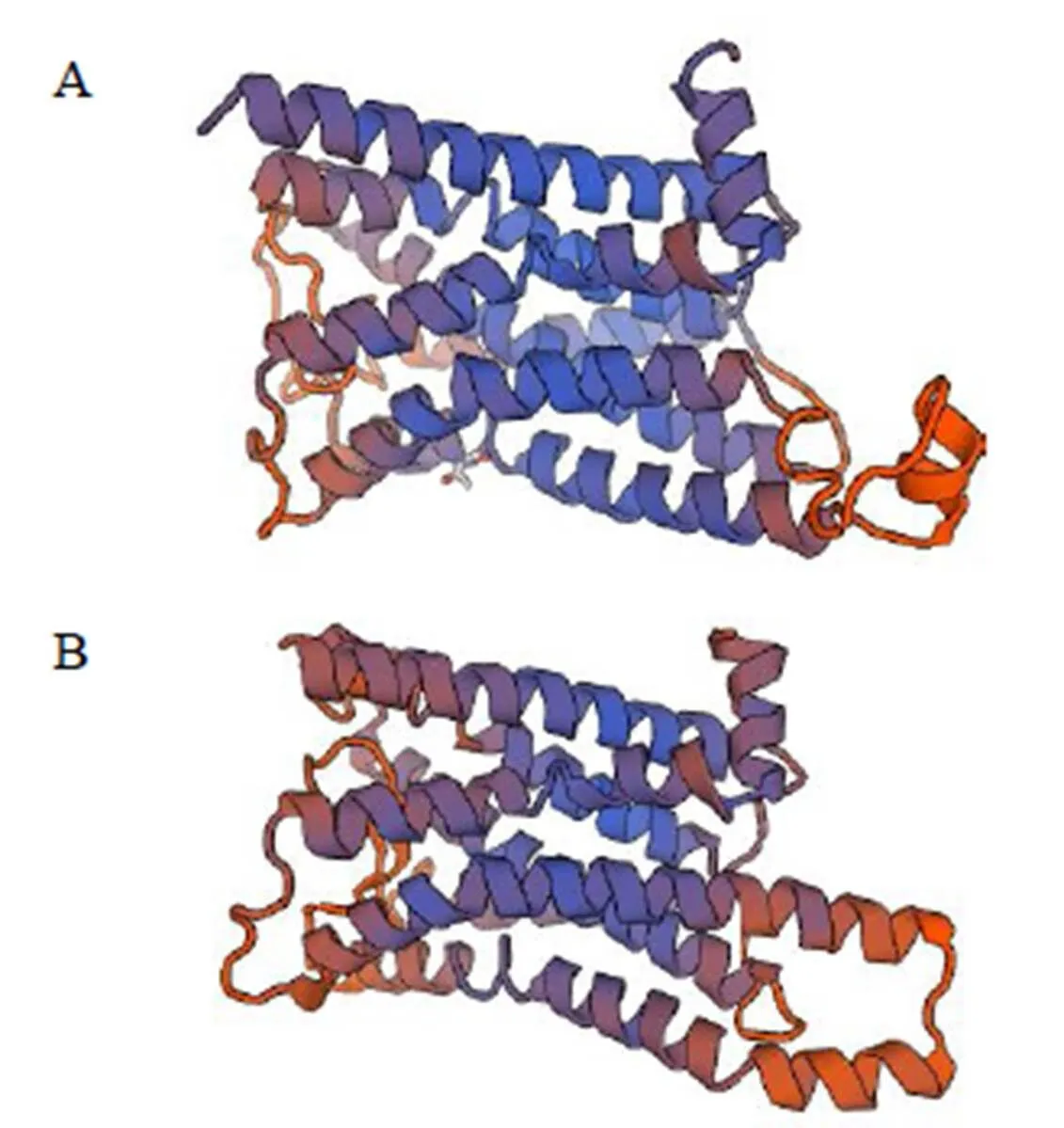
图4 虾夷扇贝(A)、马氏珠母贝(B)β2AR蛋白分子结构

“*”与灰色背景均表示保守的氨基酸
“*”and the gray background indicated the conserved amino acid
图5结构域区的氨基酸序列多序列比对
Fig. 5 Multi-sequence alignment of amino acid sequence of Pdomains
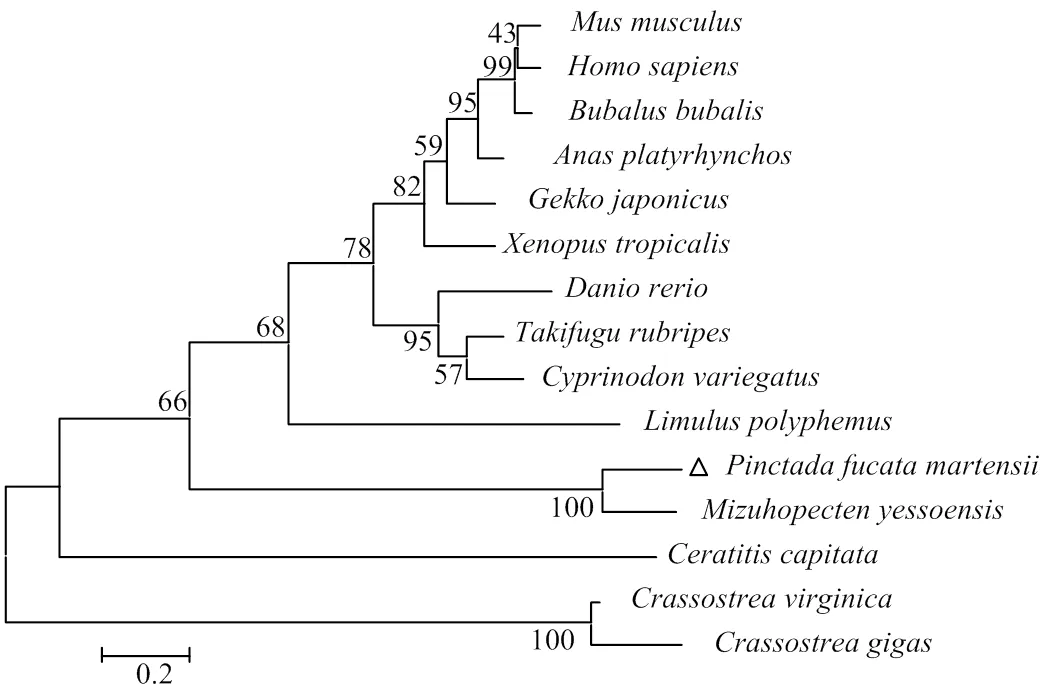
图6 基于NJ法构建的β2AR系统进化树
2.4 Pmβ2AR的组织表达分析
在马氏珠母贝的血细胞、鳃、闭壳肌、性腺、肝、中央膜区、足和套膜区中均有表达,在鳃中表达量最高(< 0.05);其次是肝胰腺和性腺,在闭壳肌中表达量最低。
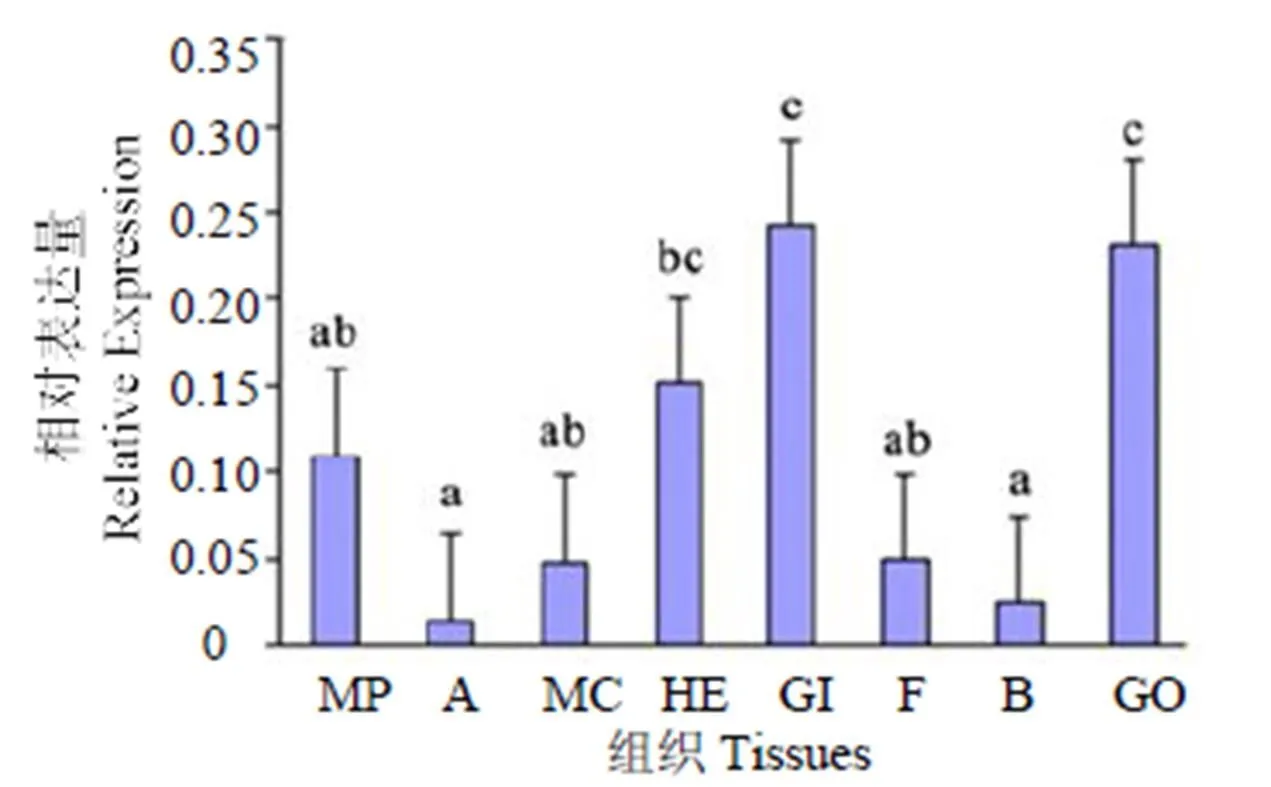
MP: 套膜区; A: 闭壳肌; MC: 中央膜区; HE: 肝胰腺; GI: 鳃; F: 足; B:血细胞; GO: 性腺; 凡有一个标记相同字母即为差异不具统计学意义(> 0.05)
MP: Pallial zone; A: Adductor muscle; MC: Central zone; He: Hepatopancreas; GI: Gill; F: Foot; B: Blood cells; GO: Gonads; Same letters indicated no significant difference(>0.05)
图7在马氏珠母贝不同组织中的表达分布
Fig. 7 Expression distribution ofin different tissues from
3 讨论
目前的研究主要集中在脊椎动物中,无脊椎动物中特别是海洋软体动物的研究极少。本实验通过软件分析显示氨基酸序列与其他物种的具有相似的7个跨膜结构。经软件预测没有信号肽,氨基酸序列分析认定为亲水蛋白。是受体蛋白,分布在3个细胞相中,而N端位于细胞外表面并延伸到疏水区域,其与细胞内和细胞外的亲水环相接触,与前人关于的研究相符合[16-17]。同源性分析显示基因在不同物种间保守性较高,其中与虾夷扇贝的相似性为70%,进化树分析显示两者聚为一支,表明两者遗传上关系较近,同属于软体动物这一类别。
为进一步探讨的功能,我们检测了在马氏珠母贝中不同组织中的表达量情况,发现在鳃中表达量最高,其次是肝胰腺和性腺。Christine等[18]的研究表明在腮腺组织中高表达,高表达有利于伤口修复。软体动物的鳃和人类的皮肤一样,是抵御有害环境的主要防御屏障。马氏珠母贝的鳃直接与环境接触,对水中的细菌和有害物质起到过滤作用[19]。鳃具有丰富的毛细血管,能促进血管舒张,鳃中还具有各种抗菌活性成分,并能进行多种酶类分子的合成、某些应激反应等过程[20]。在马氏珠母贝鳃中高表达,推测其可能在抵抗各种外界环境刺激时发挥了作用,但具体的功能仍需进一步验证。在性腺中的高表达表明与生殖细胞的成熟过程、减数分裂密切相关[21],这与在缢蛏各组织中的相对表达一致[11]。贝类没有特异性免疫,肝胰腺可分泌各种免疫酶来对抗外来物的干扰[22]。笔者推测,在贝体受到外界刺激后,激活GRK2或β-arrestin2通路,促进肝胰腺分泌免疫酶来对抗外界刺激,这进一步说明了肝胰腺在贝类先天性免疫中的作用。
在免疫调节方面具有重要作用。在人体中,能调解支气管扩张、血管舒张和促进去甲肾上腺素释放[23]。在炎症反应中,使新生大鼠心肌成纤维细胞中的 IL-6 和 TNF-α升高[24]。TNF-α即肿瘤坏死因子-α,主要作用是调节免疫细胞的功能,作为一种内源性致热源,它能够促使发热,引起细胞凋亡;IL-6 是白细胞介素的一种,其作为促炎细胞因子和抗炎性肌球蛋白起作用[25]。在缢蛏水管损伤修复过程中,表达量的上升与介导 TNF-α和 IL-6 的合成并在炎症反应中发挥作用有关。
4 结论
本研究成功克隆了基因的全长,预测的cDNA全长是2 426 bp,包括开放阅读框(ORF)2091 bp,编码696个氨基酸,5′UTR长144 bp,3′UTR长191 bp,预测分子质量79.0 ku,理论等电点9.18。蛋白为亲水蛋白,N-端不存在信号肽,具有7个典型的跨膜结构域。多重序列比对结果表明在不同物种间的保守性较高。NJ法构建系统进化树结果表明,马氏珠母贝与虾夷扇贝聚为一簇,与虾夷扇贝的亲缘关系最近。qRT-PCR结果表明,基因在各组织中均有表达,在鳃中表达量最高,在肝胰腺和性腺中表达量也较高。
[1] ASSEM E S K, RICHTER A W. Comparison of in vivo and in vitro inhibition of the anaphylactic mechanism by-adrenergic stimulants and disodium cromoglycate[J]. Immunology, 1971, 21(5): 729-739.
[2] PADRO C J, SANDERS V M. Neuroendocrine regulation of inflammation[J]. Seminars in Immunology, 2014, 26(5): 357-368.
[3] THERON A J, STEEL H C, TINTINGER G R, et al. Can the anti-inflammatory activities ofagonists be harnessed in the clinical setting?[J]. Drug Design, Development and Therapy, 2013, 7(7): 1387-1398.
[4] JOHNSON M. Molecular mechanisms of-adrenergic receptor function, response, and regulation[J]. Journal of Allergy and Clinical Immunology, 2006, 117(1): 18-24.
[5] MANTYH P, ROGERS S, ALLEN C, et al. Beta 2-adrenergic receptors are expressed by glia in vivo in the normal and injured central nervous system in the rat, rabbit, and human[J]. Journal of Neuroscience, 1995, 15(1): 152-164.
[6] PULLAR C E, RIZZO A, ISSEROFF R.-Adrenergic receptor antagonists accelerate skin wound healing[J]. Journal of Biological Chemistry, 2006, 281(30): 21225-21235.
[7] BRISTOW M R, HERSHBERGER R E, PORT JD, et al. Beta-adrenergic pathways in nonfailing and failing human ventricular myocardium[J]. Circulation, 1990, 82(2): I12-25.
[8] MIALET-PEREZ J, GREEN S A, MILLER W E, et al. A primate-dominant third glycosylation site of the-adrenergic receptor routes receptors to degradation during agonist regulation[J]. Biol Chem, 2004, 279 (37): 38603-38607.
[9] 陈艳敏, 肖瑞平, 王珏.-肾上腺素能受体激动剂治疗心衰研究进展[J]. 药学进展, 2017(3): 163-169.
[10] GOYARTS E, MATSUI M, MAMMONE T, et al. Norepinephrine modulates human dendritic cell activation by altering cytokine release[J]. Experimental Dermatology, 2010, 17(3): 188-196.
[11] 何圣耀, 杜蕴超, 牛东红, 等. 缢蛏-肾上腺素能受体基因及其在损伤修复中的作用[J/OL].基因组学与应用生物学:1-10[2019-03-30].http://kns.cnki.net/ kcms/ detail/45.1369.Q.20190116.1506.002.html.
[12] WANG Q, LIU Y, ZHENG Z, et al. Adaptive response of pearl oyster,, to low water temperature stress[J]. Fish & Shellfish Immunology, 2018, 78 (7): 310-315.
[13] 王姿曼, 郝瑞娟, 邓岳文, 等. 马氏珠母贝基因的克隆及表达[J]. 广东海洋大学学报, 2019, 39(1): 14-21.
[14] 雷超, 贝伟烈, 郑哲, 等. 马氏珠母贝B型清道夫受体基因克隆与表达性分析[J]. 广东海洋大学学报, 2017, 37(1): 7-14
[15] 郑哲, 吴宣瑾, 焦钰, 等. 马氏珠母贝基因的克隆及组织表达[J]. 广东海洋大学学报, 2019, 39(3) 24-29.
[16] SUCHAROY C C. Beta-adrenergic pathways in human heart failure[J]. Expert Review of Cardiovascular Therapy, 2007, 5(1):119.
[17] RUBENSTIN R C. The hydrophobic tryptic core of the beta-adrenergic receptor retainsregulatory activity in response to agonists and thiols[J]. Journal of Biological Chemistry, 1987, 262(34): 16655-16662.
[18] YEH C K, CHANDRASEKAR B, LIN A L, et al. Cellular signals underlying-adrenergic receptor mediated salivary gland enlargement[J]. Differentiation, 2012, 83(1): 68-76.
[19] 焦钰. 马氏珠母贝基因cDNA的分子特征及组织表达分析[J]. 中国农学通报, 2014, 30(20): 35-41.
[20] 杜俊俏, 刁晓平, 郑鹏飞, 等. 芘暴露对马氏珠母贝鳃和肝胰腺抗氧化酶活性的影响[J]. 生态环境学报, 2013, 22(10): 1711-1716.
[21] 王志新, 梁海鹰, 杜晓东, 等. 马氏珠母贝热休克蛋白基因的克隆与表达分析[J]. 广东海洋大学学报, 2013, 33(6): 14-23.
[22] TISCAR P G, MOSCA F. Defense Mechanisms in Farmed Marine Molluscs[J]. Veterinary Research Communications, 2004, 28(1): 57-62.
[23] URSINO M G, VASINA V, RASCHI E, et al. The beta3-adrenoceptor as a therapeutic target: current perspectives[J]. Pharmacological Research, 2009, 59(4): 221-234.
[24]KOLMUS K, TAVERNIER J, GERLO S.-Adrenergic receptors in immunity and inflammation: Stressing NF-KB[J]. Brain, Behavior, and Immunity, 2015, 45(3): 297-310.
[25] FERGUSON-SMITH A C, CHEN Y F, NEWMAN M S, et al. Regional localization of the interferon-beta 2/B-cell stimulatory factor 2/hepatocyte stimulating factor gene to human chromosome 7p15-p21 [J]. Genomics, 1988, 2(3): 203-208.
Molecular Characterization and Expression Analysis ofAdrenergic Receptor () from
ZHANG Yu-ting1, ZENG Man-yu1, Ma Jia-yi1, Li Jia-xin1, JIAO Yu1,2, DU Xiao-dong2
(1./ 2.,524088,)
【Objective】To study the biological function of(). 【Methods】The full length ofgene was cloned using rapid amplification of cDNA end technology(RACE)and its sequence characteristics were analyzed. Meanwhile, the expression ofmRNA in different tissues ofwas detected by Real-time PCR(RT-PCR).【Result and conclusion】It shows that the total length ofgene was 2 426 bp, including 2 091 bp of the open reading frame(ORF)which encoded 696 amino acids, a 5′ untranslated region of 144 bp and a 3′ untranslated region of 191 bp. The predicted molecular weight was 79.0 ku, and the isoelectric point was 9.18. Multiple sequence alignment results showed thatwas conservative among different species. Domain analysis showed that the amino acid sequence ofhad 7 typical transmembrane domains. In addition, qRT-PCR indicated thatwas expressed in all the detected tissues, and the highest expression level was in the gills, followed by hepatopancreas and gonads.
;; molecular characterization; expression analysis
Q954.4
A
1673-9159(2019)04-0108-07
10.3969/j.issn.1673-9159.2019.04.016.2019.04.016
2019-04-01
国家自然科学基金(31672626)
张雨婷(1995-)女,硕士研究生,研究方向为水产养殖学。Email:zytgou@163.com
焦钰,女,副教授,研究方向为生物化学与分子生物学。Email:jiaoyu1981@hotmail.com
张雨婷,曾嫚玉,马佳仪,等. 马氏珠母贝β2肾上腺素能受体()分子特征和表达分析[J].广东海洋大学学报,2019,39(4):108-114.
(责任编辑:刘朏)
猜你喜欢
家教世界·创新阅读(2019年11期)2019-12-10 09:22:04
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
人生与伴侣·共同关注(2018年8期)2018-02-16 06:58:48
凤凰生活(2018年1期)2018-01-09 17:49:48
天津科技大学学报(2016年1期)2016-02-28 16:59:45
广东海洋大学学报(2015年4期)2016-01-13 08:39:30
广东海洋大学学报(2015年4期)2016-01-13 08:39:28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
现代检验医学杂志(2015年2期)2015-02-06 02:01:01
作文周刊·小学一年级版(2014年31期)2014-04-29 00:44:03
