
What is a tree in the Mediterranean Basin hotspot?Acritical analysis
2019-07-08 02:35FrricdailAnneChristineMonnetDanielPavonToniNikolicPanayotisDimopoulosGianluigiBacchettaJuanArroyoZoltBarinaMarwanCheikhAlbassatnehGianniantonioDominaBrunoFadyVladoMatevski0StephenMifsudandAgatheLeriche
Forest Ecosystems 2019年2期
Frédéric Médail,Anne-Christine Monnet,Daniel Pavon,Toni Nikolic,Panayotis Dimopoulos,Gianluigi Bacchetta,Juan Arroyo,Zoltán Barina,Marwan Cheikh Albassatneh,Gianniantonio Domina,Bruno Fady,Vlado Matevski0,Stephen Mifsud and Agathe Leriche
Abstract Background:Tree species represent 20%of the vascular plant species worldwide and they play a crucial role in the global functioning of the biosphere.The Mediterranean Basin isone of the 36 world biodiversity hotspots,and it is estimated that forests covered 82%of the landscape before the first human impacts,thousands of years ago.However,the spatial distribution of the Mediterranean biodiversity is still imperfectly known,and a focuson tree species constitutes a key issue for understanding forest functioning and develop conservation strategies.Methods:We provide the first comprehensive checklist of all native tree taxa(species and subspecies)present in the Mediterranean-European region(from Portugal to Cyprus).We identified some cases of woody species difficult to categorize as trees that we further called“cryptic trees”.We collected the occurrences of tree taxa by“administrative regions”,i.e.country or large island,and by biogeographical provinces.We studied the species-area relationship,and evaluated the conservation issues for threatened taxa following IUCNcriteria.Results:We identified 245 tree taxa that included 210 speciesand 35 subspecies,belonging to 33 families and 64 genera.It included 46 endemic tree taxa(30 species and 16 subspecies),mainly distributed within a single biogeographical unit.The countries with the highest tree richness are Greece(146 taxa),Italy(133),Albania(122),Spain(155),Macedonia(116),and Croatia(110).The species-area relationship clearly discriminated the richest central-eastern(Balkans)and northern(Alpine and Cevenno-Pyrenean)biogeographical provinces,against the five western provinces in the Iberian Peninsula.We identified 44 unrecognized“cryptic trees”,representing 21%of the total trees.Among the 245 taxa identified,19 are considered to be threatened(15 CR+EN+VU)or near threatened(4 NT)by IUCN.Conclusions:The Mediterranean-European region includesan unsuspectedly high number of tree taxa,almost 200 tree taxa more than in the central European region.Thistree diversity isnot distributed evenly and culminates in the central-eastern part of the Mediterranean region,whereas some large Tyrrhenian islands shelter several narrow endemic tree taxa.Few taxa are recognized as threatened in the IUCNRed list,and the vulnerability of these species isprobably underestimated.
Keywords:Plant biogeography,Mediterranean region,Threatened trees,Tree definition,Tree distributions
Background
A recent census of the biomass distribution on Earth indicates that plants,mostly terrestrial species,represent about 80%(i.e.≈450 Gt C)of the sum of the biomass across all taxa worldwide,and that plant biomass includes about 70%stems and tree trunks(Bar-On et al.2018).This result underlines the crucial role of trees in the global functioning of the biosphere.With ca.60,000 tree species representing 20%of the vascular plant species worldwide according to GlobalTreeSearch(Beech et al.2017),trees constitute a major component of the structure and function of forest ecosystems located in temperate and tropical biomes.Higher levels of ecosystem services,such as biomass production,water supply,nutrient cycling and soil carbon storage,characterize forests with a higher tree diversity(Gamfeldt et al.2013).Since forests play a crucial role in the conservation of terrestrial biodiversity by providing habitats and ecological niches for a majority of plants and animals(www.globaltrees.org),it is necessary that assessments of the different tree taxa involved into the composition of ecosystems should be comprehensive and robust by including a biogeographical perspective.
Nevertheless,the global knowledge of the number and distribution of tree species remains surprisingly low both at the international level(Beech et al.2017)and for large biogeographic areas(regions and provinces).While threats on the tropical forests urge the scientists to fill the gap in knowledge on their diversity(Serra-Diaz et al.2017),the five Mediterranean ecoregions(Mediterranean Basin,California,central Chile,Cape Region of South Africa and south-western Australia)still require further studies.With only 2%of the world's terrestrial surface,Mediterranean ecoregions contain nearly 20%of the Earth's total plant diversity,making very significant biodiversity hotspots,second only after tropical ones(Cowling et al.2005).The Mediterranean Basin is one of these 36 current biodiversity hotspots worldwide(Médail and Myers 2004),including both regional(Médail and Quézel 1997)and local hotspots,i.e.micro-and nano-hotspots sensu Cañadas et al.(2014).It is estimated that Mediterranean forests covered 82%of the landscape before the beginnings of human impacts on ecosystems thousands of years ago(WWF 2001).But the whole Mediterranean Basin still comprises a high tree richness estimated to 290 indigenous taxa(species and subspecies)including 201 endemics(Quézel and Médail 2003;Médail 2008).The spatial distribution of this woody plant biodiversity is still imperfectly known even in Mediterranean-European region.A biogeographical focus on tree species distribution is needed for conservation of the Mediterranean forests which have undergone rapid changes in structure and composition in recent decades(Mazzoleni et al.2004),even if forest surface remains stable on the global scale(FAO and Plan Bleu 2018).
Among the woody diversity,the definition of a tree per se might be problematic and requires some agreement(e.g.Gschwantner et al.2009).This is especially the case for historically disturbed tree species that often appears as shrubs and display as true trees only after a long period without disturbance or in cultivation in gardens.This aspect is particularly important in the Mediterranean region where the impacts of natural and human disturbances are both very old and severe.
In thisstudy,we provide the first checklist of all tree taxa(speciesand subspecies)present in the northern part of the Mediterranean ecoregion(i.e.the Mediterranean-European region),since Mediterranean forests occur mainly in this area(in Europe,forestscover ca.33%of total land area:see Alberdi Asensio et al.2015)and theavailabledataare more robust and readily available there than in the southern and south-eastern Mediterranean.
Therefore,the main objectives of this study are:(i)to examine whether an extensive survey of the whole putative tree taxa challenge the definition of what is generally considered as a Mediterranean tree;(ii)to provide a comprehensive checklist of tree taxa for the Mediterranean-European region,i.e.from Portugal to Cyprus;(iii)to examine the global spatial distribution of tree taxonomic diversity(natives and endemics)at the biogeographical and administrative scales.Finally,we compared the distribution of tree diversity in the Mediterranean-European region from previous assessment made in the whole Mediterranean Basin and the European continent and discussed the implications for conservation.
Methods
Study area
We defined the Mediterranean-European region as the North Mediterranean terrestrial ecoregion following the definition of terrestrial ecoregions of the world from Olson et al.(2001)(Fig.1).They provide a biogeographic scheme defined by existing global maps of floristic provinces and by regional maps of units based on the distribution of selected groups of plants and on published regional classification systems of terrestrial habitats.The biogeographical limits for the Mediterranean region are similar to those proposed by Médail and Quézel(1997),except for some areas of the mountain ridge of the Italian peninsula and of the Balkans.Indeed,climate indices are not strictly relevant to define the biogeographical limits per se,notably for the upper parts of the Mediterranean mountains.
Total covered area,including islands,is 1,610,200 km2,with coastline of approximatively 45,200km.The orographic variation in the area is large,and the altitudes range from 0m a.s.l.up to 3482m a.s.l.(Mulhacén,Sierra Nevada-Spain).
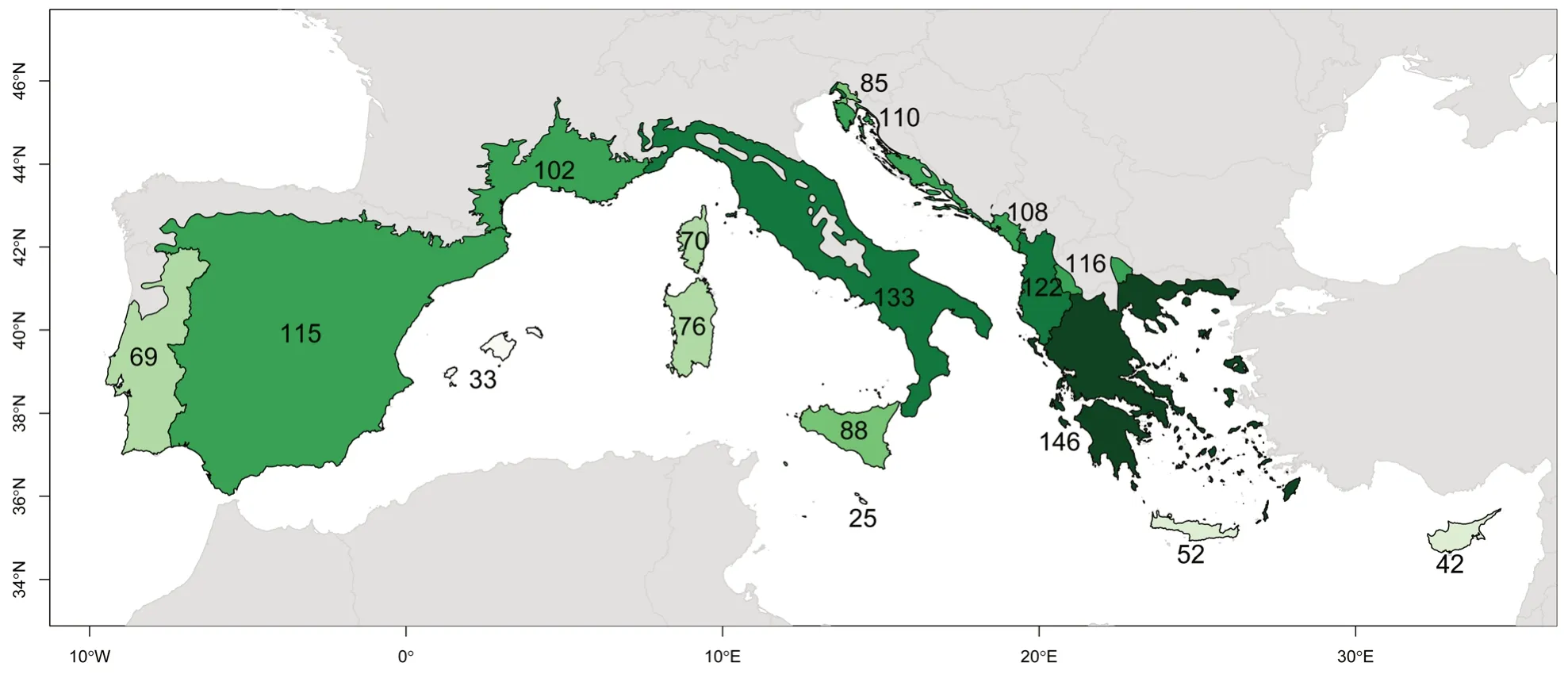
Fig.1 Number of native tree taxa(species or subspecies-including endemic and putative native)in each continental administrative area and main islands in the Mediterranean-European region.Darker greens indicate higher richness
The study area coversthe following countries(as administrative units)from west to east:Portugal,Spain,France,Italy,Malta,Slovenia,Croatia,Montenegro,Albania,Macedonia,Greece and Cyprus.Larger Mediterranean islands(in addition to Malta and Cyprus)corresponding to the Balearic archipelago,Corsica,Sardinia,Sicily and Crete,are treated as independent units to achieve a better spatial resolution of tree taxa and island specificities.The study area does not include the non-European Mediterranean area,i.e.North Africa and Near East.
The area is characterized by a Mediterranean climate types marked by mild and humid winters,hot and dry summers and a relative long period of aridity each year which impose an effective and severe drought on the plants.Rainfall patterns and the oceanity are extremely varied,and this explain why the Mediterranean Basin is characterized by the lowest rainfall reliability(quantified as interannual variations in monthly and seasonal rainfalls,and asthe frequency of individual rainfalls)of the five Mediterranean-climate ecosystems of the world(Cowling et al.2005).In general,annual rainfalls are greater in the northern than in the southern areas of the Mediterranean Basin and increase with elevation in the mountain ranges.During moist season the rainfall can be unpredictable and often intense from autumn through spring.
Mediterranean vegetation types vary widely with environmental constraints(climate,geology,geomorphology,soil type),dominant plant species,and disturbances induced by man and its herds.The woody vegetation is constituted by diverse types of matorrals or shrublands(maquis,garrigue,phrygana)and forests.In relation to the bioclimate type and the altitude,several major types of this vegetation can be recognized in the Mediterranean-European region(Quézel and Médail 2003;Rivas-Martínez et al.2007;Médail 2008):(i)a thermo-Mediterranean belt from sea level to~200-500(800)m a.s.l.,dominated by sclerophyllous communities most often as a narrow strip along the coast(with Olea europaea,Ceratonia siliqua,Chamaerops humilis,Pistacia lentiscus,Pinus halepensis,Pinus brutia,etc.);(ii)a meso-Mediterranean belt,between~100-500(1000)m a.s.l.,mainly with sclerophyllous forests(Quercus ilex,Quercus suber and Q.coccifera)or Pinus halepensis/P.brutia forests in areas with low rainfall or formerly disturbed by man;(iii)a supra-Mediterranean belt,between~500-1500(1800)m a.s.l.,with diverse deciduous oaks forests(including Acer,Carpinus,Ostrya,Quercus and Sorbus)in the more humid ombrotypes;(iv)a mountain-Mediterranean level(~1500-2000 m a.s.l.)including Fagus and deciduous and semi-deciduous Quercus forests and notably coniferous forests with Pinus nigra,Pinus sylvestris and firs(Abies alba and most of the Mediterranean Abies spp.)and even cedar(Cedrus libanis.l.)on Cyprus.
An extremely important feature of this woody Mediterranean vegetation is that it has been dramatically influenced by human activities for thousands of years(e.g.Thirgood 1981;Blondel et al.2010).Forest logging,fires,grazing,agriculture development,soil disturbances,excessive uptake of water to the detriment of riparian trees,changes in the distribution of native species due to large reforestations,aforestations,and introductions of alien taxa led to massive landscape transformations and changes in the structure and dynamicsof these natural forest ecosystems(Quézel and Médail 2003;Blondel 2006).Mediterranean islands are in this respect of particular concern,due to the higher threat they suffer,as a combination of higher typical sensitivity to disturbances,and long history of human occupancy(e.g.Médail 2017).For this reason,we considered larger islands as separate units in our analysis.
Tree definition
The IUCN's Global Tree Specialist Group(GTSG)defines a tree as“a woody plant with usually a singlestem growing to a height of at least two meters,or if multi-stemmed,then at least one vertical stem fivecentimetersin diameter at breast height”(Beech et al.2017).Our definition is slightly different and we define as tree the plants with the following biological features:(i)secondary growth(Fahn 1990);(ii)perennials(they live many years,mostly for decades or centuries);(iii)typically(at least in some environmental conditions or locations)having a single stem or trunk,growing orthotropic with monopodial or more often sympodial branching systems,and bearing lateral branches at some distance from the ground;and(iv)the height of the mature individual is at least three meters.Such plants correspond to the growth-form“phanerophytes”according to Raunkiær(1934)and Ellenberg et al.(1991)(originally P,excluding NP=nanophanerophytes,height 0,5-5 m),or phanerophytes“scapose”(originally Pscap),phanerophytes“caespitose”(originally Pcaesp)and phanerophytes“striscianti”(originally Prept)according to Pignatti(1982).
By strictly following those criteria,we identified some cases of woody species difficult to categorize that we further called“cryptic trees”.Indeed,this definition includes some taxa that generally occur as shrubs with poorly defined multicolous stems but can form true trees under certain environmental conditions or in situations where the disturbances have been absent or very reduced for at least several decades.Surprisingly,it turned out that these cases were not rare in the study area.This is the case for instance of Spartium junceum or Genista etnensis(Fabaceae)which usually grows in the form of a bush(nanophanerophyte)of less than 5 m high.We included them in the checklist because in some localities of Corsica,Sardinia and Sicily(Aeolian islands)these taxa meet the above criteria of tree.Many of the tree species reported in this study may very frequently appear as shrubs or even prostrate scrubs due to intense disturbance,particularly herbivory.
Occurrences and status by administrative or by biogeographical areas
Occurrences of each tree taxa were compiled(i)by country(n=12)and large island(n=5)(hereafter,“administrative regions”),and(ii)by biogeographical provinces.To date,a finer spatial approach is not available for all taxa per countries throughout the study area.
The definition of the biogeographical provincesfollowed the Biogeographic map of Europe provided by Rivas-Martínez et al.(2004).Hence,thirteen biogeographic provinces occur in the study area(Fig.3a).Three of them are included into the Eurosiberian biogeographical region but are representative of transition zones between the Mediterranean and the Eurosiberian regions(Cevenno-Pyrenean,Alpine,and Apennino-Balkan);the other ten are included in the Mediterranean region.We studied the species-area relationship within the administrative or biogeographical regions by plotting the total native tree taxa richnessof each geographic unit as a function of their surface,after log transformation.
For each species or subspecies,the presence in each administrative or biogeographical unit,was defined as native(N)or introduced(I).We consider endemic tree taxa as range-restricted taxa in and outside of the study area,in general located within a unique biogeographical province sensu Rivas-Martínez et al.(2004).
We marked taxa with unclear autochthonous or allochthonous origin as putative native(i.e.“N?”,e.g.Tetraclinis articulata in Malta),or putative introduced(i.e.“I?”).Taxa for which the presence is not confirmed but possible were quoted as putative presence(i.e.“?”,e.g.Crataegus pentagyna in Montenegro).Many cultivated and sometime naturalized tree species have not been considered.We focused only to a small number of taxa that have been cultivated for a long period(archeophytes,i.e.present before the classical date of the“discovery”of America in 1492)and,apart from culture,are widely naturalized in some parts of the studied area but also native in some areas of the Mediterranean-European region i.e.Castanea sativa Mill.,Ceratonia siliqua L.,Cupressus sempervirens L.,Ficus carica L.,Juglans regia L.,etc.(see Additional file 1).We considered threatened tree taxa using the IUCN(2018)classification system,as taxa included into the three classes:critically endangered(CR),endangered(EN),or vulnerable(VU).The other categories included near threatened taxa(NT),least concern taxa(LC),and data deficient taxa(DD)are also considered in the present work.
Data sources
The data were collected and aggregated for each country and large island.The primary sources of data were the international database Euro+Med PlantBase(http://www.emplantbase.org/home.html),Flora Europaea(Tutin et al.1964-1993),and the volume series Chorology of trees and shrubs in south-west Asia and adjacent regions(Browicz 1982-1996).These comprehensive data were carefully completed and checked with available national floras,national databases,regional databases,other publications,and our own expertise,when the aforementioned sources were not available:Albania(Barina et al.2017;Z.Barina pers.comm.),Corsica(Jeanmonod and Gamisans2013;CBN Corse pers.comm.),Crete(Chilton and Turland 1997;http://www.cretanflora.com/;Strid 2016),Croatia(Nikolić 2004-onward),Cyprus(Hand et al.2011-onward),continental France(Silene-Flore database:http://flore.silene.eu,IGN Inventaire Forestier:http://inventaire-forestier.ign.fr/,Tison et al.2014),Greece(Dimopoulos et al.2013,2016;Strid 2016),Iberian Peninsula i.e.Balearic Islands,Spain and Portugal(Anthos:http://www.anthos.es;Castroviejo 1986-2015;Sociedade Portuguesa de Botânica 2014),Italy(Bartolucci et al.2018;Galasso et al.2018;Vegitaly:http://www.vegitaly.it/),Macedonia(V.Matevski ined.),Malta(Mazzocchi 1969;Baldacchino and Stevens 2000;Mifsud 2002-2014;Casha 2015;Baldacchino 2018;Calleja 2018);Montenegro(F.Médail&A.-C.Monnet obs.pers.),Sardinia(Arrigoni 2006-2015;Bacchetta et al.2012;G.Bacchetta obs.pers.),Sicily(Giardina et al.2007),Slovenia(Jogan et al.2001).
In addition,some other more thematic and comprehensive data sources were used such as the European Atlas of Forest(San-Miguel-Ayanz et al.2016)and completed by some recent papers concerning the description of some new tree taxa,such as Tamarix minoa in Crete(Villar et al.2015)or two cryptic species in the Alnus glutinosa group(Vít et al.2017).Data synthesis was led by D.Pavon&F.Médail between 2014 and 2018.
Taxonomy and nomenclature
The present checklist of Mediterranean-European trees includes only the taxa at the species and subspecies levels which meet both criteria:“study area”and“tree definition”.This list does not contain other sub-specific levels,such as varieties,forms,or hybrids,because of their taxonomical and nomenclatural instabilities.We followed the classification of the Angiosperm Phylogeny Group for the definition of families(APGIII 2009;APG IV 2016).The nomenclature of Mediterranean and European trees is often fluctuating and controversial between the different major taxonomic works.We considered in priority the Euro+Med database(http://www.emplantbase.org/home.html)and the Plant List database(https://www.theplantlist.org),but also the major national floras(see references in the“data sources”paragraph).In case of taxonomic-level disagreement,we retained the most consensual taxonomic level based also on recent phylogenetic studies or systematic revision of a given group,if available,adding consulting local experts or recent floras.
Results
Mediterranean-European trees in a nutshell
Our analysis of all the taxa that can potentially be tree species in the Mediterranean-European region provides a checklist of 245 tree taxa(i.e.species and subspecies),including 210 speciesand 35 subspecies(see Additional file 1).These taxa belong to 33 different families;the most represented families are Rosaceae(42 taxa),Fagaceae(33 taxa),Pinaceae(21 taxa),Salicaceae(20 taxa),Sapindaceae(19 taxa),Betulaceae(16 taxa),Tamaricaceae(16 taxa),and Cupressaceae(14 taxa).
These tree taxa are included in 64 different genera which represents 86%of the total tree genera(n=74)found in the whole Mediterranean biogeographic region(Quézel and Médail 2003).Four of these genera(Chamaerops,Phillyrea,Spartium,Tetraclinis)have their distribution centered in the Mediterranean region and can be considered as Mediterranean endemics sensu lato(at the biogeographical region level),whereas three taxa have a Tethysian origin(Ceratonia,Myrtus,Nerium)and occur in adjacent biogeographic regions as well.The other genera have larger distributions,in the Palearctic realm or beyond.
Among this checklist,46 are endemic trees(30 species and 16 subspecies),i.e.mainly located within a single biogeographic province sensu Rivas-Martínez et al.(2004).This means that the rate of endemism is equal to 18.9%if we consider the species and subspecies levels,or 14.3%at the species level.The genera with the more endemic taxa are Quercus(6 taxa),Abies,Acer,and Pinus(4 taxa each),Alnus,Salix and Sorbus(3 taxa each).The presence of three narrow endemic species included in two relict genera(Liquidambar and Zelkova)and characteristic of the Cenozoic paleoflora distributed in the Palearctic realm should also be noted(Palamarev 1989).
Even though we only took into account taxa considered as indigenous to a given country or a biogeographical area,the native status of some tree species can sometimes be difficult to estimate,notably when they are frequently cultivated or used for afforestation,or when their center of origin or native range are still unclear (e.g.Castanea,Ceratonia,Ficus,Juglans,
Olea:see discussion).Overall,it concerns 24 taxa(ca.10%of the total checklist)that are widely cultivated or selected for afforestation purposed by forest services.The distribution of those native but locally domesticated Mediterranean tree taxa will be discussed in detail below since it represents a crucial issue for the conservation of local genetic resources.
This checklist also includes 44 unrecognized real trees or“cryptic trees”,i.e.usually with a shrubby habit but which form a tree habit under certain environmental conditions(Table 1).These taxa belong to four main families(Rosaceae:11 tree species,Cupressaceae:5 tree species,Anacardiaceae and Fabaceae:4 tree species each),and to two main genera(9 tree species in Crataegus and 5 in Juniperus).These cryptic trees represent the impressive and unexpected rate of≈18%of the Mediterranean-European trees.

Table 1 The 44 unrecognized real trees or“cryptic trees”,i.e.usually with a shrubby form,occurring in the Mediterranean-European region,with indications about their distribution and ecology
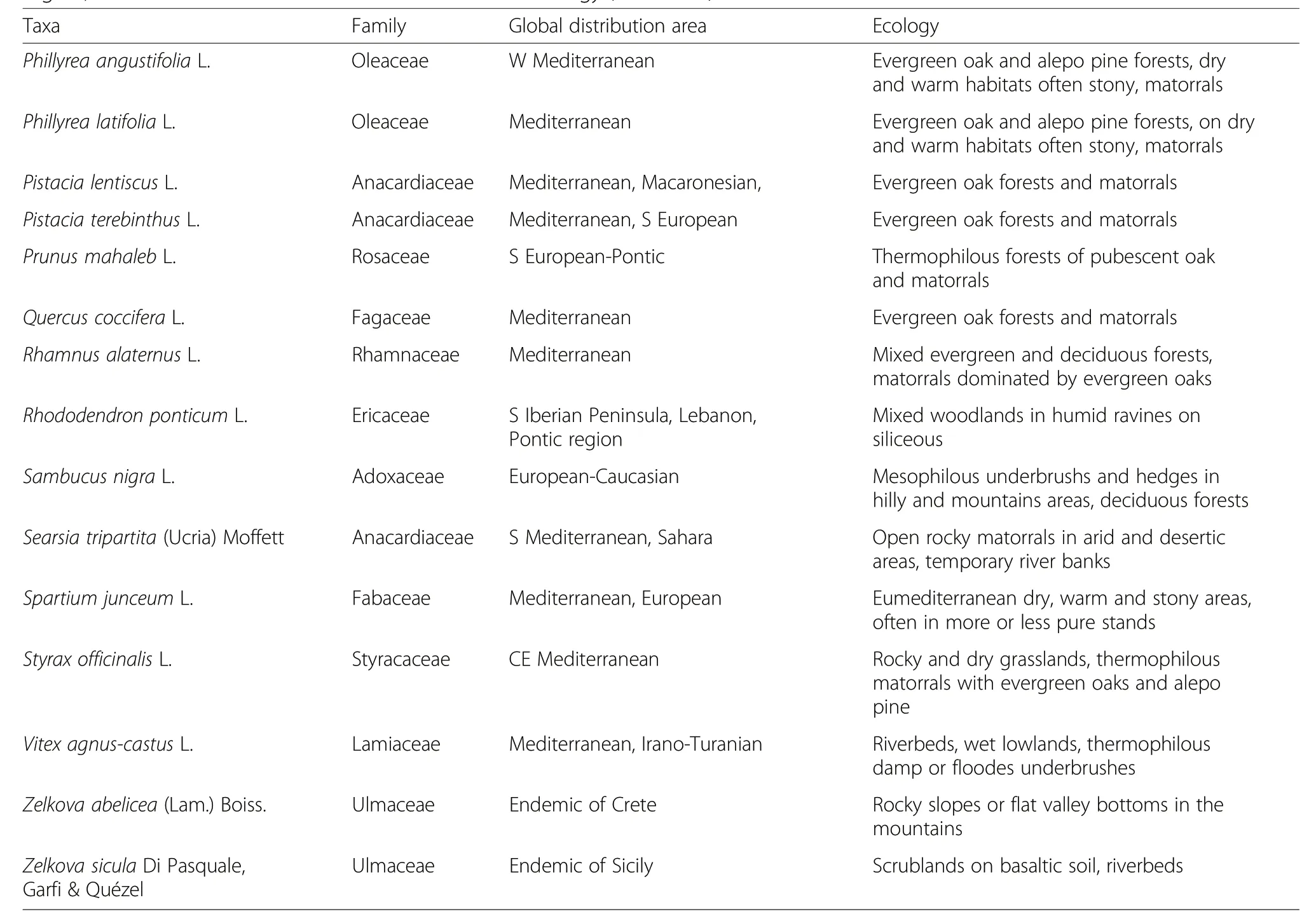
Table 1 The 44 unrecognized real trees or“cryptic trees”,i.e.usually with a shrubby form,occurring in the Mediterranean-European region,with indications about their distribution and ecology(Continued)
Mediterranean-European tree diversity by administrative regions
Mapping the presence of tree species in each Mediterranean-European country is important to estimate conservation priorities at the administrative level.Our analysis shows that the countries(continental parts)with the highest total richness of native trees(>110 taxa)are Greece(146 taxa),Italy(133 taxa),Albania(122 taxa),Spain(115 taxa),Macedonia(116 taxa),and Croatia(110 taxa)(Fig.1a,Table 2).There is a strong disparity in total richness between the large Mediterranean islands:Sicily has the highest richness(88 taxa),and the Balearic Islands the lowest one(35 taxa),whereas Corsica and Sardinia in thewestern Mediterranean Basin exhibit a similar richness(respectively 70 and 76 taxa)and Cyprus and Crete in the eastern basin share quite similar level of tree richness(respectively 42 and 52 taxa).
While the total tree richness is concentrated mainly in the Balkans,endemic trees mostly occurred in the large countries of the three peninsula in western,central and eastern Mediterranean area(Table 2).Greece(11 endemic taxa),Spain(9 endemic taxa),and Italy(7 endemic taxa)exhibited the highest number of endemic taxa.Emphasis should be given to the place of some large Tyrrhenian islands that shelter a significant number of endemic trees(Sicily:10 taxa,Sardinia:7 taxa).
The species-area relationships show contrasting results between administrative regions and large Mediterranean islands(Fig.2).The relationship is significant for the islands(log richness islands:-1.89841+0.25788 log surface islands,p=0.01)but not for the continental areas of the countries(log richness continents:3.8459+0.03493 log surface continents,p=0.41).The continental areas of the central and eastern Mediterranean countries,from Italy to Greece(i.e.mainly the Balkans),have a significantly higher tree taxonomic richness than those located further west or on the large islands.Thus,the islands always have a lower richness per unit area than continents.This is particularly the case for Malta and the Balearic Islands.Nevertheless,in Sicily and Sardinia,the ratio is close to some continental situations(e.g.France,Slovenia)and is even higher that Portugal which shows the lowest area-species ratio for a continental country.

Table 2 Number of total native tree taxa(including endemic and putative native taxa),endemic tree taxa,and putative native tree taxa)by administrative areas and main islands of the Mediterranean-European region.“Taxa”include both species and sub-species level
Mediterranean-European tree diversity by biogeographical areas
To go beyond their distribution in the administrative regions,we extracted the number of native tree taxa(species and subspecies)for each of the 13 biogeographic provinces sensu Rivas-Martínez et al.(2004)(Fig.3a).The raw numbers of tree richness are culminating in the central-eastern provinces of the Mediterranean-European region:Apennino-Balkan(142 taxa),Graeco-Aegean(137 taxa),Adriatic(135),and Italo-Tyrrhenian(128 taxa).The lowest richness(40 taxa)is found in the Cilicio-Phoenician province which corresponds only(in our study area)to the island of Cyprus(Fig.3b).The highest taxonomic richness in endemic trees occurs in the Italo-Tyrrhenian(n=16 taxa)and in the Graeco-Aegean(n=12 taxa)provinces,then in the Betican(n=9 taxa)and Adriatic(n=8 taxa)provinces(Fig.3c).The lowest endemic richness(n=1 taxa)is found in southern France,and in the southern parts of the Alpine and Cevenno-Pyrenean provinces.
The species-area relationship for biogeographic provinces clearly discriminated the central-eastern(mainly Balkans)and northern(Alpine and Cevenno-Pyrenean)provinces,against the five western provinces in the Iberian Peninsula(Fig.4).
Threatened trees
Among the 245 tree taxa of the checklist,15 constitute threatened taxa sensu IUCN(2 CR,7 EN,6 VU)and 4 are classified as near threatened taxa(NT)(Table 3).Only 154 tree taxa(145 species and 9 subspecies)are classified as“least concern”(LC),and 23 species with data gap for assessment(DD).What is most worrying is that 84 taxa(19 species and 65 subspecies)included in the checklist are not considered at all in the global IUCN assessment.Eleven of these threatened tree taxa,including some very narrow endemic trees such as Abies nebrodensis,Cytisus aeolicus,Rhamnus persicifolia,Zelkova abelicea and Zelkova sicula are only distributed on some large(notably Sicily,Crete,Cyprus and Sardinia)or medium (few Croatian islands sheltering Pinus nigra subsp.dalmatica populations)sized islands of the Mediterranean Sea.
Discussion
Distribution of tree diversity in the Mediterranean-European region
Our comprehensive analysis of tree taxonomic diversity in the Mediterranean-European region identified 210 tree species or 245 species and subspecies,including 46 endemic trees of the study area.This is of course a tiny portion of the estimated tree diversity worldwide(ca.60,065 tree species currently known to science,see Beech et al.2017),but an important number regarding to the European forests.Tree taxonomic diversity was estimated at 135 species and subspecies within the medio-European region (Quézel and Médail 2003),whereas only 42 tree species from 11 families occur in Central Europe(Leuschner and Meier 2018).It represents also of high tree diversity compared to the one in the entire Mediterranean biogeographical region which was estimated at 290 species and subspecies(Quézel and Médail 2003),i.e.85%of the trees of the whole Mediterranean region occur in the European part.This key result shows the importance of the southern part of Europe in preserving this rich biological heritage for a temperate region.
The taxonomic richnessof the Mediterranean-European region is also found in tree genera,since 86%of the total genera(n=74)of the whole Mediterranean biogeographic region are present in this European part.Only 11 genera(Acacia,Amygdalus,Argania,Balanites,Calotropis,Diospyros,Dracaena,Moringa,Prospis,Pterocarya,Ziziphus)are restricted to the southern or eastern parts of the Mediterranean region.
These results are not surprising since Mediterranean forests are very diverse and heterogeneous regarding their structure,their specific composition and their dynamics(Quézel and Médail 2003),and this explains the existence of 35 terrestrial sub-ecoregions including forests(WWF 2001).The main forest types are:(i)sclerophyllous forests(notably with the oaks Quercus ilex,Q.suber,Q.coccifera);(ii)dry conifer(Pinus halepensis,P.brutia) forests;(iii) deciduous mixed forests with broad-leaved oaks(Quercus pubescens,Q.faginea,Q.ithaburensis)and other genera(Acer,Carpinus,Fraxinus,Sorbus);(iv)mountain conifer forests with Abies,Cedrus,Juniperus,Pinus nigra.Tree diversity and the ecological complexity in forest types can be explained both by the complex paleogeography and historical biogeography of the Mediterranean region,and also by the high heterogeneity in the climate(Médail 2008;Blondel et al.2010).
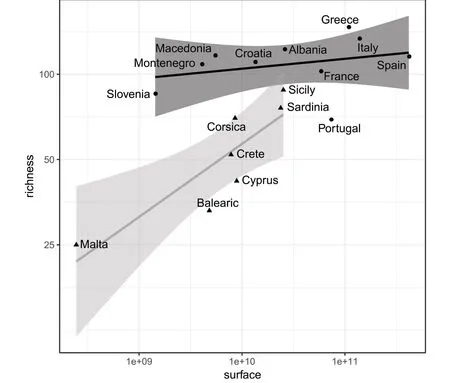
Fig.2 Linear regressions between the total native tree taxa richness and the surface of continental administrative areas(black regression line and black circles)or main islands(grey regression line and black triangles)in the Mediterranean-European region with their respective 95%confidence interval.Abscissa and ordinate axes have a logarithmic scale.Dotsabove each line indicate higher richnessthan predicted by the species-area relationship,and dots below each line indicate lower richness than expected
This woody biodiversity is unevenly distributed within the Mediterranean-European region.Our analysis,performed either at the administrative or biogeographic level,indicate that the continental areas of the central and eastern Mediterranean countries,from Italy to Greece(i.e.mainly the Balkans),have a significantly higher tree richness compared to those located further west(Iberian Peninsula)or on large Mediterranean islands.The higher taxonomic diversity in the eastern Mediterranean Basin is found in several other studies and this pattern is probably related to the key biogeographical crossroad of this area and to the existence of more suitable conditions during the LGM(notably warmer summer temperatures)inducing larger biodiversity refugia(Médail and Diadema 2009;Fady and Conord 2010).Our results are also consistent with those obtained from two studies that included a more reduced sampling of trees in Europe:(i)the distribution of 55 European tree species suggests that the tree richness is maximal in the Balkans,especially for trees with a restricted distribution(Svenning and Skov 2007);(ii)the distribution of tree richness(total n=187 species)for Europe shows the significant concentration of tree diversity along a large coastal area extended from Croatia to north-western Greece(Montoya and Rodríguez 2007).Both studies concluded that the main driver of the variation of tree richness is the current climate,and that the climate during the Last Glacial Maximum(LGM,ca.20 ky ago)or the period since an area became free of ice have a significant influence,notably to explain the concentration of range-restricted tree taxa.Some studies also revealed that older climatic events throughout the Miocene were also critical for extinction in the western part of the Mediterranean Basin(the Iberian Peninsula)and may partly explain the relative lower richness of tree taxa there(Postigo Mijarra et al.2009).The species-area relationships indicate that Mediterranean islands always have a lower richness per unit area,and only the two largest islands,Sicily and Sardinia,are grouped with the continental territories of higher tree richness.For Sicily,this pattern can also be explained by the proximity of this large continental island to the Italian Peninsula and its connection to the mainland during the LGM which facilitated the putative latitudinal migration of trees.
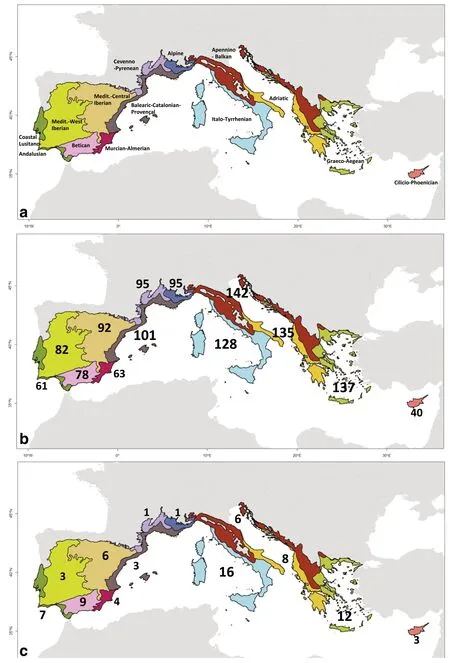
Fig.3 a Distribution of the 13 biogeographical provinces sensu Rivas-Martínez et al.(2004)in the study area;b Number of native tree taxa(species and subspecies)and(c)Number of endemic tree taxa(speciesand subspecies)in each biogeographical province
The distribution of the native or endemic tree taxa richness by biogeographical provinces sensu Rivas-Martínez et al.(2004)shows congruent results.Taxonomic richness is highest in the central-eastern Mediterranean provinces,notably:(i)in the Graeco-Aegean and the Italo-Tyrrhenian provinces for both native and endemic taxa,(ii)in the Apennino-Balkan and Adriatic provinces for native tree richness only.By contrast,in the western Mediterranean,only the Betican Provinceisnoteworthy for itsendemic tree richness,since this area represents a hotspot for the differentiation of the endemic flora through geographical isolation and/or repeated specialization to contrasted environments(Molina-Venegaset al.2015).
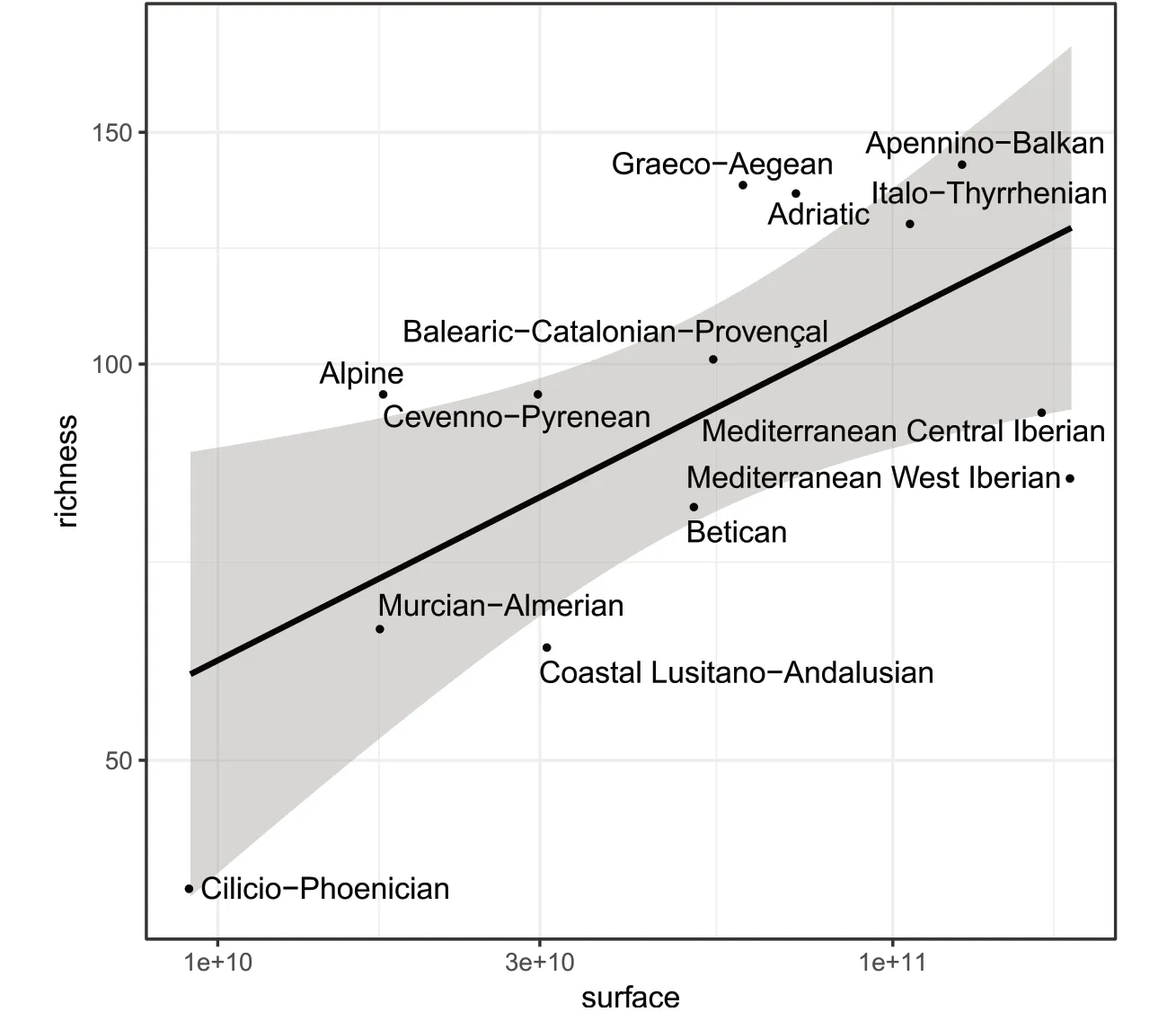
Fig.4 Linear regression between the total native tree richnessand the surface of each of the 13 biogeographic provincessensu Rivas-Martínez et al.(2004).Abscissaand ordinate axeshave a logarithmic scale.Shade arearepresentsthe 95%confidence interval around the predictions.Dotsabove the line indicate higher richnessthan predicted by the species-area relationship,and dotsbelow the line indicate lower richnessthan expected
Of course,this assessment ultimately depends on the taxonomic conceptions used,which can vary very significantly depending on the published sources.We retained a rather restrictive approach and not consider trees with uncertain taxonomic status as Bartolucci et al.(2018)did for instance in the recent Updated checklist of the vascular flora native to Italy;for the only island of Sicily,11 tree taxa could be included(Betula aetnensis,Fraxinus excelsior subsp.sicilienesis,Malus crescimannoi,Pyrus castribonensis,Pyrus ciancioi,Pyrus sicanorum,Pyrus vallis-demonis,Rhamnus lojaconoi,Salix nebrodensis,Sorbus madoniensis,Sorbus busambarensis),but more precise taxonomic studies are hardly needed.
What is a Mediterranean tree?
In this study,we defined as tree a species with a height that can potentially reach 3m,with a single stem and an arborescent habit(i.e.with a well individualized crown).This corresponds mainly to the definition of phanerophytes in the Raunkiær system for categorizing life forms(Raunkiær 1934).Therefore,several woody taxa of the Mediterranean matorrals were excluded(e.g.Amelanchier ovalis,Anthyllisbarba-jovis,Viburnum tinus),even if they can exceed 3m high.On the contrary,theonly two monocot species,the Mediterranean palms Chamaeropshumilis and Phoenix theophrasti,with a solitary or multiple trunks,were included in our checklist even if they do not form a real trunk(but a stipe)from an anatomical point of view.Our definition is quite similar,but more restrictive,to that of the IUCN's Global Tree Specialist Group:“a woody plant with usually a single stem growing to a height of at least two meters,or if multi-stemmed,then at least one vertical stem five centimetersin diameter at breast heigh t”(Beech et al.2017).
However,while our tree definition is more conservative than that of the GTS database,our analysis highlights the absence of 22 tree taxa in this latter(Table 4).Assessing if a taxon could sometimes have the habits of a tree or if it is always a shrub was one of the biggest difficulties during the elaboration of the checklist.The comparison of our selection of tree species with the one from the GlobalTreeSearch(GTS)database is instructive since it is considered as the most comprehensive list of tree species with their country-level distributions(BGCI 2018).The missing taxa correspond for almost half(n=10)to species that possess a shrub habit in general but can form trees in certain geographical situations(e.g.Spartium junceum in the Aeolian Islands,Italy)or ecological conditions (e.g.Buxus balearica,Crataegus laciniata,Nerium oleander,Phillyrea angustifolia,Rhododendron ponticum).Some other missing taxa are related to recently described species(e.g.Alnus lusitanica and A.rohleana,see Vit et al.2017),or to different taxonomic conceptions(e.g.Abies borisii-regis or Quercus gussonei considered or not as a hybrids).
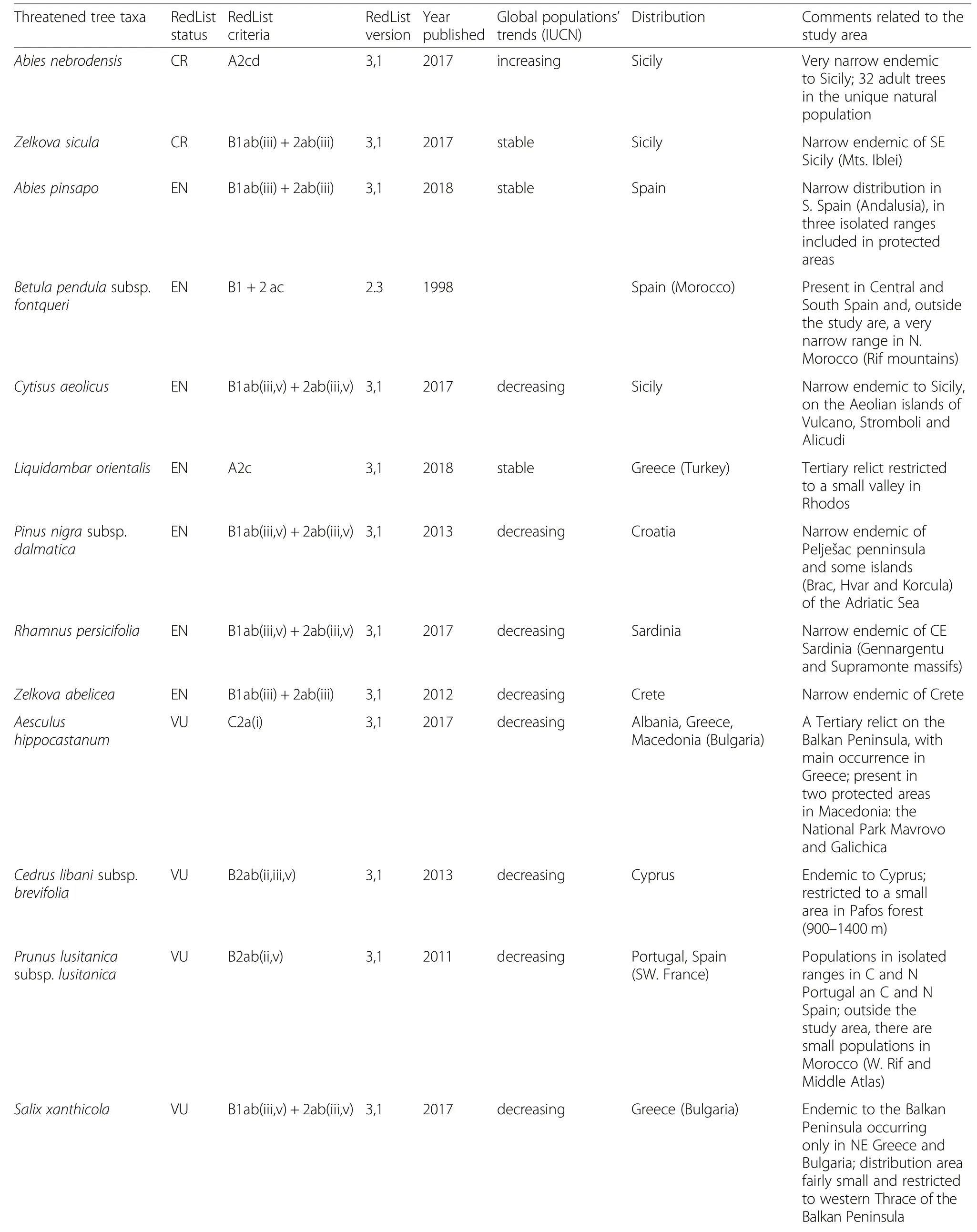
Table 3 Threatened tree taxa sensu IUCNRed List categories(CR,EN,VU,NT)(IUCN 2018)occurring in the Mediterranean-European region(in brackets the distribution outside the study area)

Table 3 Threatened tree taxa sensu IUCNRed List categories(CR,EN,VU,NT)(IUCN 2018)occurring in the Mediterranean-European region(in brackets the distribution outside the study area)(Continued)
Thus,our precise assessment of the expression of the potential arboreal character of the woody species located in the Mediterranean-European region(based on the literature data and on many in situ novel observations)allows us to identify 44 unrecognized real trees,or“cryptic trees”,representing 21%of the total trees occurring in this area(Table 1).This result constitutes an important issue to better understand the links between shrub vs.tree habit in relation to current and future climatic conditions(Olson et al.2018),but also to past and current human impacts which are very important in the Mediterranean region for several millennia(Thirgood 1981).With increasing drought and warming,trees worldwide experience mortality or dieback and it is important to understand how plant height relates to climate.A recent meta-analysis suggests that plant height is linked to climate via plant hydraulics which explains why vegetation height differs across biomes and is altered with climate change(Olson et al.2018).In the Mediterranean region,due to millenia-long human impact,we consider that the ecophysiological expression of the tree habit has been often inhibited in most of ecological situations,in parallel with the increase of climate drought.The very arid conditions that many Mediterranean ecosystems experience implies severe constraints on tree architecture and growing.This is the case of the trees belonging to the mesophilous relict genus Zelkova(Ulmaceae):the dwarfed Zelkova abelicea trees(a narrow Cretan endemic)can attain ages>500 yr.and these individuals survive under high browsing pressure for centuries,surpassing normally growing trees of the same population in number and age(Fazan et al.2012).

Table 4 The 22 tree taxa occurring in the Mediterranean-European region but not considered in the GlobalTreeSearch database(http://www.bgci.org/global_tree_search.php?action=about;see also Beech et al.2017)
While some cryptic trees are narrow endemics(e.g.Cytisus aeolicus and Genista tyrrhena from the Aeolian archipelago in Sicily),most of them are widespread taxa(e.g.Chamaerops humilis,Erica arborea,Myrtus communis,Juniperus oxycedrus,Pistacia lentiscus,Phillyrea latifolia,Prunus mahaleb)that form the keystone species of most of the matorrals(Quézel and Médail 2003).For example,under some constant humid and thermophilous conditions of a coastal marsh,a real forest dominated by impressive Myrtus communis(Myrtaceae)and Phillyrea latifolia(Oleaceae)trees has been recently described in southern Corsica Island(CBN Corse,ined.).Some other cryptic trees are much rarer,and their discoveries were surprising.This is the case of a 12-m high individual of Cotinus coggygria(Anacardiaceae),with a trunk circumference of 60 cm,found in a Montenegro forest(F.Médail and A.-C.Monnet,ined.,VI.2018).
An interesting evolutionary question would be to analyze more finely whether the insular conditions also favor the expression of the arborescent habit for these trees of the Mediterranean islands,an insular syndrome already demonstrated for various endemic groups on some remote oceanic islands,due to speciation by cladogenesis(adaptive radiation,e.g.Lens et al.2013).The remarkable lability in growth forms between herbaceous and woody habits in these insular situations suggest that the transition from a bush habit to a tree habit is an easier step.But it remains a neglected and overlooked process,even if it must have played a quite significant role in the dynamics and adaptive response of woody plants within a high-stress biome such as the Mediterranean region.Another alternative explanation could be related to better refuge conditions in some situations:unusual big trees or cryptic trees would represent in some Mediterranean areas,notably islands,the remains of old forests in lesser harsh and more stable climates.
The complex native status of domesticated or useful Mediterranean trees
Since the Neolithic,human pressures on Mediterranean ecosystems,especially on the forests,have been intense and the profound transformations of natural into cultural landscapes are the result of millennia of human activities(Thirgood 1981;Quézel and Médail 2003;Blondel 2006).With the exploitation of resources provided by tree species,humans interfered with their natural distribution.These useful plants have been gathered or cultivated since ancient times and some of them(e.g.olive,walnut and chestnut trees)are even used as biomarkers for estimating the beginnings of anthropization of a given Mediterranean area(e.g.Mercuri et al.2013;Ucchesu et al.2015).This explains why it is often difficult,and to some extent controversial,to estimate the native status of domesticated Mediterranean trees to a given country or a biogeographical area,especially because the putative sympatric presence of wild specimens and of cultivated varieties in a same locality.
Among the 24 trees that are widely cultivated or selected in afforestation by forest services in the Mediterranean-European region,the origin of several of them has been particularly discussed by biogeographers.But some recent phylogeographical and/or paleoecological studies provide sometimescounterintuitiveresults.For somesclerophyllous species(the laurel tree Laurus nobilis:Rodríguez-Sánchez et al.2009;Marino et al.2014;the myrtle,Myrtus communis:Migliore et al.2012)that are not domesticated but only used by humans,the classical hypothesis of an eastern Mediterranean origin followed by subsequent westward expansion and diversification across the western Mediterranean and Macaronesia is verified.These cases suggest here a limited influence of historical human-mediated translocations despite the ancient use of these species.But for other widely domesticated Mediterranean trees,such an east-west vicariance across the Mediterranean region is not applicable.The biogeography and domestication of the olive tree(Olea europaea),the most iconic Mediterranean tree,are well documented(e.g.Besnard and Rubio de Casas 2016).Four large areas of persistence(macro-refugia)of oleaster populations occurred during the Last Glacial Maximum(LGM),with probable isolated micro-refugia in the central Mediterranean Basin;then,came the post-glacial diffusion of oleaster throughout the Mediterranean Basin facilitated by human intervention,but a single primary domestication of the olive seems to have occurred in the eastern Mediterranean.Similar global patterns of persistence into multiple peri-Mediterranean refugia followed by post-glacial migrationsthat preceded domestication seem to apply to other domesticated trees such as the common fig,Ficus carica(Khadari et al.2005)and the carob tree,Ceratonia siliqua(Baumel et al.2018).This challenges the classical hypothesis of an exclusive eastern Mediterranean origin and then a westward colonization for these important fruit tree species.
The difficulty of estimating the native distribution arises also in other cultivated medio-European trees that are not only present in the Mediterranean region.It exists indeed conflicting results between an ecological niche model projecting a significant presence of walnut(Juglans regia)in the Balkan Peninsula during the LGM in contrast to genetic analysis that suggest refugia in Southwest Asia(Aradhya et al.2017).For another key cultivated tree,the chestnut(Castanea sativa),a paleodistribution modelling approach validated with pollen and charcoal records(Roces-Díaz et al.2018)seems to be in agreement with a phylogeographical analysis(Mattioni et al.2013):LGM refugia were suggested in the north of the Iberian and Italian peninsulas,in Greece,and along the southern coast of the Black Sea.But if the hypothesis of chestnut migration from Turkey and Greece to Italy and Spain is clearly rejected,the authors cannot conclude whether“the populations from Italy and Spain could have originated from common refugia in western Europe or could have resulted from human-mediated transplanting of materials or both”
(Mattioni et al.2013)!These uncertainties are also linked to genetic sampling that is too incomplete with respect to the current distribution area of chestnut,with unfortunately the same shadow areas in the Mediterranean region(Northwest Balkans,Corsica,Sardinia)as in an older phylogeographic study that resulted in quite different conclusions about refugia locations,notably along the Italian peninsula(Fineschi et al.2000).This example is a good illustration of the difficulties that still exist in drawing up robust biogeographical histories of cultivated trees who are indigenous somewhere in the Mediterranean region.
Intensive reforestation carried out in the North-Mediterranean region generates the translocation of forest species outside their natural range of distribution,and these practices may alter the phylogeographic structures for widely used taxa(e.g.Pinus brutia,Pinus halepensis,Pinus nigra,Pinus pinaster,Pinus pinea,Quercus suber).It happens also for much less used and unsuspected tree species such as the manna ash(Fraxinusornus):in Southeastern France,thistree isnative in the Maritime Alps but invasive further west,along some rivers of the Hérault department(Thébaud and Debussche 1991).Sometimes,a depauperate low genetic diversity together an intense human-induced movement for reforestation preclude any phylogeographic signal,as for the emblematic and widespread Pinus pinea(Vendramin et al.2008).These frequent reforestation practices can blur the biogeographic signature of forest stands at the regional scale,with detrimental consequences for the conservation of local genetic resources.
Conservation issues
As many trees are threatened around the world(BGCI 2018),it is necessary to get an accurate assessment of the conservation status of the different tree taxa in a comprehensive biogeographical scheme and using the most robust data available,i.e.from the IUCN(2018).However,the IUCN Red List contains only a limited number of Mediterranean trees and some old or inadequate assessments should be reviewed again.The presence of only 19 Mediterranean trees considered to be threatened(15 CR+EN+VU)or near threatened(4 NT)is probably not representative of the global level of threat that these woody contingent experiences in a region that has been managed,modified,and,in some places,heavily degraded by human activities for millennia(e.g.Thirgood 1981;Blondel et al.2010).Indeed,only 7.7%of the 245 species and subspecies we identified are considered threatened according to the IUCN database,despite that the global contingent of the North Mediterranean trees includes 46 endemic trees that have a narrow distribution range.This is not surprising because at the world scale only a tiny percentage(11%)of trees has been assessed by the IUCN Red List criteria,compared with other major species groups(Fauna and Flora International 2013).
Some rare Mediterranean endemic taxa(e.g.Abies pinsapo,Phoenix theophrasti),or trees with a patchy distribution along the coasts(e.g.Juniperus macrocarpa on sand dunes)face dramatic habitat destruction due to the exponential development of human infrastructure,often linked to mass tourism.Several rare and narrow endemic trees linked to wet habitats are not included in the current IUCN database,such as the Sardinian endemic Salix arrigonii,whereas another tree,Tamarix minoa,occurring in a unique locality of NW Crete on sandy river banks close to the sea(Villar et al.2015),is quoted Data Deficient(DD).Among the other DD taxa,at least two characteristic taxa(Platanus orientalis,Vitex agnus-castus)currently experience a severe reduction of their populations due to the artificialization of their habitats,even if they still count a quite large number of populations.
Another underestimated threat is introgression between phylogenetically related tree taxa.This is the case of the group of black pines(Pinus nigra)where introgression risk is likely between Pinus nigra subsp.nigra,frequently planted for reforestation,and two other subspecies having a more restricted distribution range:subsp.dalmatica in Croatia,and subsp.salzmannii in France and Spain.Although infrequent and potentially beneficial for some traits,such gene flow from planted to native trees to could contribute to worsen local adaptation,for example to diseases such as Dothistroma sp.(Scotti-Saintagne et al.2018).
From a geographical point of view,we must emphasize the role of the large Mediterranean islands,which include 11 of the 19 tree taxa currently present in the IUCN database.This shows the importance of these island ecosystems for the differentiation and persistence of taxonomically singular trees,but also the severe threats in these spatially restricted systems.This is the case of Sicily which includes three rare narrow endemics(Abies nebrodensis,Cytisus aeolicus,Zelkova sicula)among which the two latter are only present on some small islands of the Aeolian archipelago(Troia 2012).Some other narrow endemic trees(Genista tyrrhena from Aeolian and Pontine archipelagos in Italy,and Quercus alnifolia restricted to the Troodos range in Cyprus)are classified as least concern(LC),but due to their restricted range and population size these evaluations are probably too optimistic.A more precise risk analysis by region would also probably highlight the high level of threat to locally several Mediterranean trees.For example,the tyrrhenian insular endemic Genista etnensis which is ranked globally as LC is classified as critically endangered(CR)in Corsica.This is also the case of several trees with a patchy distribution in specialized mountain habitats such as Juniperus drupacea,J.foetidissima,J.thurifera,and Pinus heldreichii which are classified as LC as well.Some other locally threatened taxa(Buxus balearica,Rhododendron ponticum)with a highly restricted range in the N.Mediterranean region are not even considered in the IUCN database.
Conclusion
This study provides the first comprehensive assessment of tree distributions in the Mediterranean-European region.The present checklist includes an unsuspectedly high number of tree taxa(245 speciesand subspecies),with at least 44 taxa that are generally bushy but can express a true tree architecture under certain conditions(“cryptic trees”).This means that the Mediterranean-European region has almost 200 trees more than the Central European region.This taxonomic biodiversity is highest in the central-eastern part of the Mediterranean region,notably in the Apennino-Balkan province.We must also emphasize the role of some large Tyrrhenian islands(especially Sicily and Sardinia)that shelter a significant number of endemic trees.These areas are particularly important from the point of view of functional conservation because tree species richness is positively linked to higher levels of multiple ecosystem services(Gamfeldt et al.2013),and rare species-like most of the Mediterranean endemic trees-support original and vulnerable function(Mouillot et al.2013).But threats to this woody biodiversity are often significant and may especially affect trees with narrow distributions.However,the IUCN Red list includes so far only 19 Mediterranean trees considered to be threatened(CR+EN+VU)or near threatened(NT).Therefore,the vulnerability of several of these trees is probably underestimated and further assessments are needed in the near future.
For Mediterranean trees favored or cultivated by man,sometimes for millennia,it is necessary to develop approaches combining phylogeography and paleoecology,at the scale of the distribution area specific to each taxon.This will allow a better estimate of the native status for each biogeographical area,and to consider the evolutionary dimension in the preservation of these trees.
This checklist represents the first step towards a better understanding of the diversity of trees in the Mediterranean region.The next step should be to improve the coverage in occurrences records for these tree taxa in the region in the existing tree species database(Noce et al.2016;Serra-Diaz et al.2017).These data will help to explore the key scientific questions associated to the drivers of the distribution pattern and to the shape of the species-area relationship.Quantitative investigations of ecological(e.g.,climate,soil,disturbance)versus historical(e.g.,past climate,genetic originality)drivers of current tree biodiversity can therefore be considered.This checklist will constitute also an essential background for functional and phylogenetic analyses of this key biological contingent for terrestrial ecosystems.The practical issues concern a better assessment of the threatened status of some these unappreciated trees within a highly human-modified region.We also hope such work will catalyze further assessment of tree diversity at the scale of the whole Mediterranean region(i.e.also North Africa and Middle-East)even if the data are still very complex to synthesize.
Additional file
Additional file 1:Table S1.Checklist of the tree taxa(species and subspecies)occuring in theMediterranean-European region,from Portugal to Cyprus.The definition of the families followsthe Angiosperm Phylogeny Group(APGIII2009;APGIV 2016).Theretained nomenclature of the tree taxa is compared with those of the Euro+Med database(http://www.emplantbase.org/home.html),the volume series Chorology of treesand shrubsin south-westAsia and adjacent regions(Browicz 1982-1996)and the World Checklist Kew.For the occurence of eachtree taxa in a country or a large island,we indicate the endemism status,the cultivated status,theautochthonous(native:N)or allochthonous(introduced:I)status.Taxa for which the presence is notconfirmed but possible are quoted as"putative presence»,i.e.“?”.(XLSX 71 kb)
Acknowledgments
The authors thank Aggeliki Doxa(Institute of Applied and Computational Mathematics,Heraklion),Laetitia Hugot and Paula Spinosi(Conservatoire botanique national de Corse/Office de l'environnement de la Corse),Virgile Noble(Conservatoire botanique national méditerranéen),Salvatore Pasta(CNRPalermo),Estefanía Santos Barea(Universidad de Sevilla)for their help in acquiring the data or for their comments on the distribution or status of the different tree species.
Funding
This work was funded by the French Foundation for Research on Biodiversity(FRB)through its Centre for Synthesis and Analysis of Biodiversity data(CESAB)programme,aspart of the WOODIVresearch project.ACMand MCB were funded by Labex OT-Med(n°ANR-11-LABX-0061).The fundershad no role in study design,data collection and analysis,decision to publish,or preparation of the manuscript.
Availability of data and materials
After acceptance data will be made publicly available as an additional file.
Authors'contributions
Conceived and designed the database:FM,DP,ACM,GB,PD,TNand AL.Analyzed the data:ACM and FM.Wrote the paper:FM,ACM and TN.All authors participated to the discussions leading to the framing of the manuscript during WOODIVworkshops.All authorsread,contributed suggestions and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Author details
1Aix Marseille Univ,Avignon Univ,CNRS,IRD,IMBE.Technopôle de l'Arbois-Méditerranée,cedex 4,BP 80,13 545 Aix-en-Provence,France.
2Faculty of Science,University of Zagreb,Horvatovac 102a,HR-10000 Zagreb,Croatia.3Department of Biology,Division of Plant Biology,Laboratory of Botany,University of Patras,University Campus,26504 Rio,Greece.4Centro Conservazione Biodiversità,Dipartimento di Scienze della Vita e dell'Ambiente,Universitàdegli Studidi Cagliari,Viale S.Ignazio da Laconi,13,09123 Cagliari,Italy.5Departamento de Biologia Vegetal y Ecologia,Universidad de Sevilla,Apartado 1095,41080 Sevilla,Spain.6Department of Botany,Hungarian Natural History Museum,Pf.137,Budapest 1431,Hungary.
7Sorbonne University,IEESBât 44-34,cc 237.4,Place Jussieu,Paris,France.
8Department of Agriculture,Food and Forest Sciences,University of Palermo,Viale delle Scienze bldg.4,90128 Palermo,Italy.9INRA,UR629,Ecologie des forêts méditerranéennes,Avignon,France.10Macedonian Academy of Sciences and Arts,Krste Misirkov 2,1000 Skopje,Republic of Macedonia.
11EcoGozo,Regional Development Directorate-Ministry for Gozo,Flat 6,Sunset Court B,Triq Marsalforn,Xaghra,Gozo,Malta.
Received:17 December 2018 Accepted:28 February 2019

- Forest Ecosystems的其它文章
- Improved simulation of carbon and water fluxes by assimilating multi-layer soil temperature and moisture into process-based biogeochemical model
- Potential gains from spatially-explicit coordinated planning between two large public ownerships in Minnesota
- South Carolina forestland owners'willingness to accept compensations for carbon sequestration
- Climate forcing of tree growth in dry Afromontane forest fragments of Northern Ethiopia:evidence from multi-species responses
- Wood density and carbon concentration of coarse woody debrisin native forests,Brazil