
圆叶玉兰叶片非结构性碳水化合物与氮、磷含量对海拔的响应
2019-07-08 07:14:34康喜坤陈小红张腾驹
生态学报 2019年11期
康喜坤,陈小红,龚 伟,张腾驹
四川农业大学林学院,成都 611130
非结构性碳水化合物(non-structural carbohydrates,NSC)主要指糖类和淀粉这两大类,是植物碳元素的主要供应者,是评价植物碳收支状况的一个重要量度指标,对植物的生长和适应性有很大影响[1-2]。光合作用不足时NSC可以起到缓冲作用,加上它又是光合作用的产物,因此影响光合作用的因子也会影响植物NSC的含量[3-4]。有研究表明,植物叶片的光合速率会随叶片氮含量的增加而显著提高,同时固定同化CO2和制造NSC的能力也增强[5]。磷是植物新陈代谢、能量和蛋白质合成的重要元素[6],因此叶片的光合能力及NSC 的合成不仅受氮含量的影响,也会受到磷含量的影响[7]。
濒危植物在地理分布有很大的局限性,仅生存在特殊的生境或有限的地方。而海拔是一个重要的环境因素,它会引起光照、温度、湿度等环境因素的剧烈变化,会极大地影响到植物的生长,从而使植物在生理生态适应性方面产生变化[8]。研究濒危植物的非结构性碳水化合物和氮、磷等元素对海拔的响应,可以了解濒危植物在分布环境中的碳水化合物供应状况,从营养角度探索濒危植物在不同海拔的生理变化及对海拔的适应机制,并进一步探讨不同海拔中濒危植物的碳收支状况。目前对植物NSC与氮、磷随海拔的变化研究较少,对于可利用性碳在植物生长过程中所起的作用及其与氮、磷元素之间关系还缺乏足够认识[9]。
圆叶玉兰(Magnoliasinensis)为木兰科木兰属落叶灌木或小乔木,为国家Ⅱ级重点保护野生植物、四川省特有珍稀濒危植物和优先拯救保护的极小种群野生植物[10]。圆叶玉兰零星散生于四川中部以及北部的常绿落叶阔叶混交林中,仅在芦山县和彭州市发现有成片分布,主要分布在海拔1800—2400 m的林缘和灌丛中。目前仅有曾洪和陈小红[11]对圆叶玉兰进行过种群生态学研究,其研究基础还很薄弱。本文研究了生长在芦山县不同海拔的圆叶玉兰的叶片NSC、氮、磷含量及其相互关系,拟回答下述问题(1)NSC、N、P含量随海拔升高的变化规律如何?它们之间有何关系?(2)NSC与N、P对圆叶玉兰的生长有无限制作用?(3)通过NSC含量变化,是否可以了解圆叶玉兰的碳收支状况?对这些问题的回答,可以帮助人们了解圆叶玉兰的生理生态特征,理解其在不同海拔的生长表现,探讨碳供应水平情况,并揭示这些指标在生理生态适应机制中发挥的作用。
1 试验地概况
试验地位于四川省西部的雅安市芦山县境内大川镇大熊猫基因交流走廊带(102°52′—103°11′E,30°01′—30°49′N),地势北高南低,土壤为山地黄壤,呈微酸性;属亚热带湿润气候,气候温和,年平均气温为14.1—17.9℃;雨量充沛,年降雨量达到1000—1800 mm以上。圆叶玉兰集中分布在马桑坪-汇源电站、小牛井公路沿线两旁、及黄水河支流小沟、磨坊沟、鹿厂沟沟谷两旁[11],海拔在1800—2400 m范围内,多生长在陡坡、崖壁处,其生境环境复杂、条件恶劣且地质活动频繁。圆叶玉兰分布较集中,为优势植物,仅有少数的铁杉(Tsugachinensis)、卵叶钓樟(Linderalimprichtii)、领春木(Eupteleapleiospermum)、槭属(Acer-spp.)、柳属(Salix-spp.)等乔木伴生,灌木主要有川莓(Rubussetchuenensis)、贴毛箭竹(Fargesiaadpressa)、红花悬钩子(Rubusinopertus)等木本植物,草本则有蟹甲草(Parasenecioforrestii)、野艾蒿(Artemisialavandulaefolia)、冷水花(Pileanotata)、繁缕(Stellariamedia)等。对该区内圆叶玉兰实地调查后,选取了间隔约100 m的6个海拔点作为采样点,每个采样点设置2个采样地,共计12个采样地,基本情况见表1。
2 研究方法
2.1 样品采集
2016年8月在每个采样地随机筛选3株生长健康、大小基本一致的圆叶玉兰植株,每株选取树冠中部4个方向的生长发育良好、健康的成熟叶片20枚混合取样,先在105℃下杀青30 min,再在80℃中烘干至恒重,备用。

表1 采样地基本情况Table 1 Basic situation of Magnolia sinensis
2.2 NSC组分及N、P含量的测定
将烘干样品研磨并过2 mm筛,作为测定样品。称取一部分样品采用蒽酮比色法[12]测定NSC,包括可溶性糖(葡萄糖、果糖、蔗糖)和淀粉。
称量一部分样品采用硫酸-高氯酸消煮法对样品进行处理,之后采用全自动间断化学分析仪(Smartchem 200,AMS集团)测定叶片氮含量,采用钼锑抗紫外可见分光光度法测定叶片磷含量[13]。
2.3 数据处理
NSC含量=可溶性糖含量+淀粉含量。
数据统计分析在SPSS 20.0 软件中完成,采用One-Way ANOVA分析和Duncan检验法进行多重比较,进行Pearson相关分析,相关图表制作在Microsoft Excel 2016中完成。
3 结果与分析
3.1 不同海拔圆叶玉兰叶片NSC组分和N、P含量变化
如表2所示,圆叶玉兰叶片蔗糖含量、果糖含量、可溶性糖、淀粉含量、NSC和可溶性糖/淀粉比值在不同海拔间差异极显著。圆叶玉兰叶片的蔗糖含量和淀粉含量均随海拔变化表现为“W”型的变化趋势,果糖含量、可溶性糖含量和可溶性糖/淀粉比值均随海拔升高呈现“N”型变化趋势,叶片NSC含量随海拔的增加呈现先上升后下降的变化,在P4处达最大值。圆叶玉兰叶片的蔗糖、果糖、可溶性糖、可溶性糖均在P6处显著高于P1处。
圆叶玉兰叶片的N含量、P含量、N/P比值的范围分别为10.41—18.26 mg/g、1.42—1.85 mg/g、6.70—12.91,且N含量和N/P比值在不同海拔间差异极显著。氮含量整体随海拔增加呈先下降变化趋势;磷含量随海拔变化表现为“M”型变化趋势;N/P比值随海拔的增加呈降低的变化趋势。
3.2 不同海拔圆叶玉兰叶片NSC与N、P化学计量特征的变化
由图1可以看出,可溶性糖/N、淀粉/N、NSC/N、淀粉/P在不同海拔间差异极显著,可溶性糖/P和NSC/P的比值在不同海拔间差异显著。圆叶玉兰叶片可溶性糖/N比值呈显著上升趋势;而淀粉/N和NSC/N比值呈先上升后下降的变化趋势,均在P5处达到最大值;可溶性糖/P比值随海拔的增加呈先上升后下降的变化,在P4处达到最大值。而淀粉/P和NSC/P的比值均随海拔的增加呈现“W”型的变化趋势,分别在P6和P2处达到最低值。

表2 不同海拔圆叶玉兰叶片的非结构性碳水化合物(NSC)变化Table 2 Characteristics of non-structural carbohydrates(NSC)in leaves of Magnolia sinensis
同行不同小写字母表示在0.05水平间的存在显著性差异
3.3 不同海拔圆叶玉兰叶片NSC与N、P的相关关系
对圆叶玉兰叶片的NSC与N、P含量进行Pearson分析可知(表3):蔗糖含量与果糖含量、可溶性糖含量、NSC、可溶性糖/淀粉比值均呈极显著正相关关系;果糖含量与可溶性糖含量、NSC、可溶性糖/淀粉比值呈极显著正相关关系,而与N含量、N/P比值呈显著负相关关系;可溶性糖含量与NSC、可溶性糖/淀粉比值呈极显著正相关关系,与N/P比值呈显著负相关关系;淀粉含量与NSC呈极显著正相关关系,与P含量呈显著的负相关关系,与可溶性糖/淀粉比值呈极显著负相关关系;N/P比值与P含量呈极显著负相关,与可溶性糖/淀粉比值呈显著负相关,而与N含量呈极显著正相关。
4 讨论
植物叶片的NSC组分含量通常能反映植物整体的碳供应水平,还能体现植物生长状况及其对环境的适应策略[14-15]。植物NSC组分中的淀粉因具有不可移动的特性被普遍认为是储备物质,而可溶性糖可随时被调动参与植物的各种生理代谢活动[15-16]。本研究发现,圆叶玉兰的叶片可溶性糖含量在P6处显著大于P1处,与周永斌等[12]、Li等[17]的研究结果一致,表明低温等恶劣环境对可溶性糖的有效性没有影响,也不限制高海拔植物的持续发展[12,18-22]。有研究表明,冬季树木存活与生长季末期树木组织中的可溶性糖含量水平密切相关,高的可溶性糖可以提高组织细胞液渗透压,防止细胞内结冰从而保护植物顺利度过冬天[19]。不同海拔的圆叶玉兰叶片NSC含量差异极显著,随海拔的升高表现为“低-高-低”的单峰变化过程,生长在P4处的圆叶玉兰,叶片NSC含量最高,碳水化合物供应充足,而分布于其上及其下的圆叶玉兰,因面临不同的水分、温度等环境,导致NSC含量降低,与吴杰等[23]对油竹的研究结果相似。随着海拔升高,温度降低,会导致植物的淀粉向可溶性糖转变,而本研究中圆叶玉兰叶片淀粉含量呈“W”型的变化趋势,其变化趋势较特别,除受低温影响外,还可能是受到磷元素的影响,由于圆叶玉兰叶片P含量随海拔升高呈“M”变化趋势,且不同海拔圆叶玉兰叶片的P含量与叶片淀粉呈显著的负相关关系,表明P含量对淀粉含量有抑制作用。但圆叶玉兰叶片的可溶性糖/淀粉比值随海拔升高(温度降低)而逐渐增大,表明可溶性糖/淀粉比值在圆叶玉兰抵御低温胁迫中起着重要的作用[17,22]。可溶性糖与淀粉的相互转化被公认为是一种有效的植物抗逆胁迫机制,要有足够高的糖/淀粉比值,植物才能顺利越冬,是植物对环境条件的一种积极响应[17,24],这与Li等[17]提出的在高海拔处的树木不仅依赖于总的NSC含量,而且需要高的可溶性糖/淀粉比值的观点相一致。
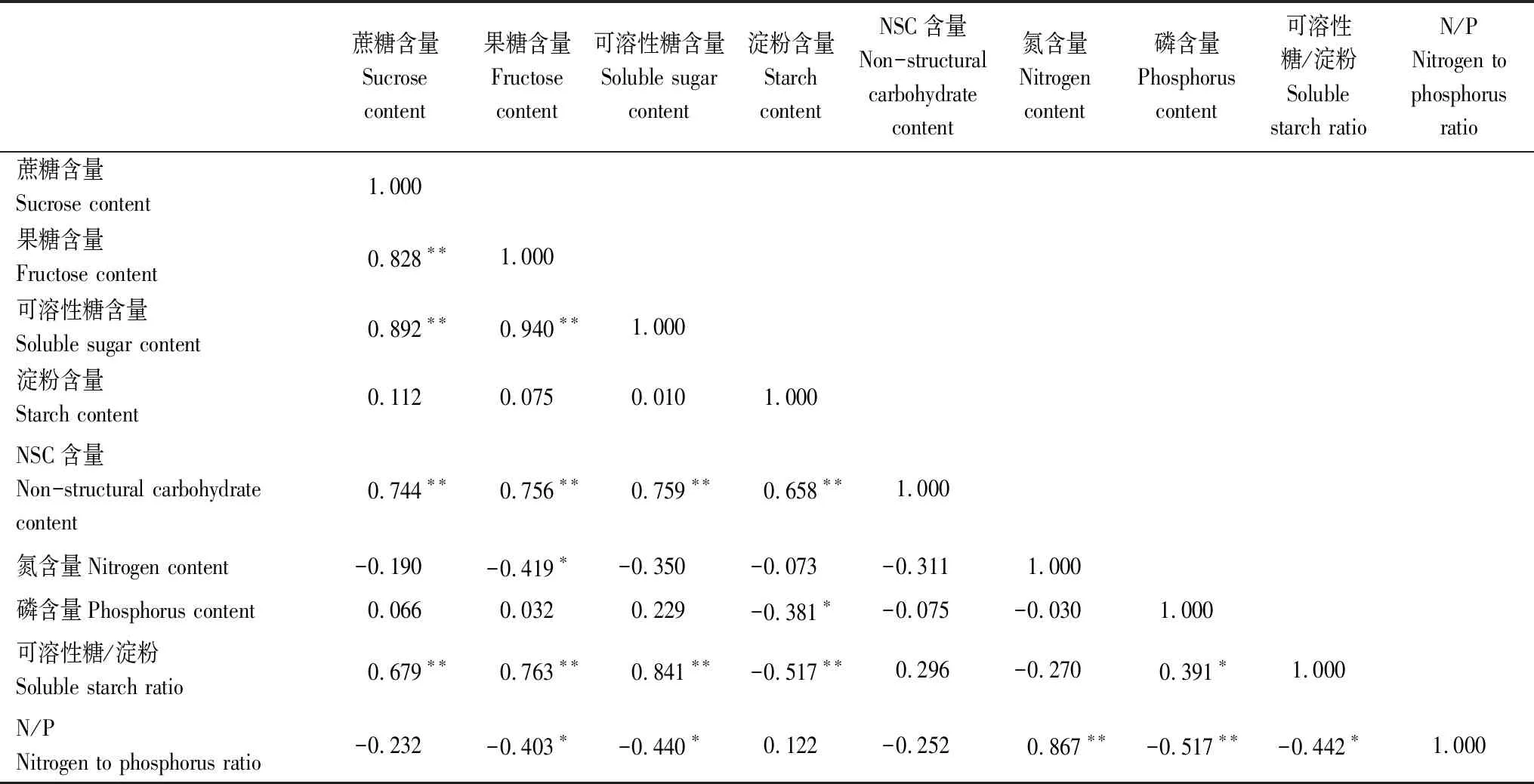
表3 不同海拔圆叶玉兰叶片NSC与N、P的相关系数Table 3 Correlation coefficients of the contents of NSC,soluble sugar,N and P in the leaves of Magnolia sinensis at different altitudes
*P<0.05,相关性显著;**P<0.01,相关性极显著
植物叶片N、P含量及其化学计量关系不但反映了植物对环境条件的适应[25],还是评估生态系统限制因子的主要指标[26-27]。有研究表明[28],N/P比值在15时被认为是临界状况;当N/P比大于16,表明其生长主要受到磷元素的限制;当N/P比小于14,则其生长主要受到氮元素的限制。本研究中,圆叶玉兰叶片的N含量和N/P比都随海拔上升而下降,且N/P比值在各海拔处均小于14,表明圆叶玉兰的生长主要受氮元素限制。圆叶玉兰叶片P含量随海拔变化表现为“M”型变化趋势,则可能是受到其生长环境土壤中P含量的影响。圆叶玉兰叶片的N含量与可溶性糖、淀粉含量和NSC均呈负相关关系,可能由于N含量增加会促进叶片中NSC组分转移到库器官,参与形态建成从而导致可溶性糖、淀粉和NSC含量下降[29];而N含量减少则会抑制NSC组分转移到库器官,使叶片中可溶性糖、淀粉和NSC含量较多。植物体NSC是植物重要的代谢能量来源,植物体NSC、N和P含量及其比例,在很大程度上反映了可供植物生长利用的物质水平、NSC 产出与N、P的投入比例关系及利用效率,而探究NSC与N、P的比例关系还可以了解N、P在植物碳代谢过程中对NSC组分含量与动态平衡的影响[17,29]。本研究中,不同海拔圆叶玉兰叶片可溶性糖/N比值显著升高,淀粉/N比值则呈“∧”型的变化趋势,即P5处叶片淀粉含量达到最高值,N含量达到最低值,淀粉/N达到最大值,这表明不同海拔圆叶玉兰叶片N含量减少主要用于可溶性糖的合成,抑制叶片淀粉的增加。可溶性糖/P比值在总体上呈升高的变化趋势,这表明圆叶玉兰叶片P含量可促进可溶性糖的累积,加速淀粉的消耗;而淀粉/P比值呈“W”型的变化趋势,P含量随海拔呈“M”型变化趋势,是由于不同海拔圆叶玉兰叶片P含量与淀粉含量变化趋势相反。NSC/N比在P6处显著高于P1处,高海拔处叶片NSC含量高意味着碳供应充足,进一步说明碳含量没有限制圆叶玉兰的生长和发育。
综上所述,随着圆叶玉兰生长地的海拔变化,其叶片的非结构性碳水化合物和氮、磷含量表现出不同的适应策略,其NSC含量较高,这表明圆叶玉兰生长可能不受碳限制。但本研究仅仅分析了不同海拔处圆叶玉兰叶片的非结构性碳水化合物和氮、磷含量,今后应结合环境因子和其他生理指标,深入探讨圆叶玉兰的生态适应机理和濒危机制,提出更好的保护策略促进圆叶玉兰的种群繁衍。
猜你喜欢
中外葡萄与葡萄酒(2022年1期)2022-01-21 09:29:10
散文诗(2021年22期)2022-01-12 06:14:04
红蜻蜓·中年级(2021年3期)2021-09-10 07:22:44
锦绣·上旬刊(2019年12期)2019-09-10 22:41:30
环球时报(2019-05-23)2019-05-23 06:17:36
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
中老年健康(2016年6期)2016-06-30 10:50:16
茶叶学报(2016年4期)2016-03-29 03:24:48
河北遥感(2014年4期)2014-07-10 13:54:58
体育教学(2009年7期)2009-09-07 10:06:12
