
不同光周期环境对谷子农艺性状的影响
2019-06-25 02:14:42贾小平全建章王永芳董志平袁玺垒李剑峰
作物学报 2019年7期
贾小平 全建章 王永芳 董志平 袁玺垒 张 博 李剑峰
不同光周期环境对谷子农艺性状的影响
贾小平1,*全建章2王永芳2董志平2袁玺垒1张 博1李剑峰1
1河南科技大学农学院, 河南洛阳 471023;2河北省农林科学院谷子研究所/ 国家谷子改良中心, 河北石家庄 050035
本研究连续2年在短日照(海南)、中等日照(河南)、长日照(吉林) 3个不同光周期环境调查160份谷子资源的抽穗期、株高、叶片数、穗长、穗粗、穗码数、码粒数、穗重、穗粒重、千粒重10个主要性状, 利用SPSS 19.0软件进行多因素方差分析及多重比较, 揭示光周期对谷子各性状的影响, 评价160份谷子资源对光周期的敏感性。结果表明, 3个不同光周期环境间谷子株高、叶片数、穗长、穗码数、抽穗期、穗粗、穗重、穗粒重、码粒数9个性状均表现出极显著差异(<0.01), 千粒重表现出显著差异(<0.05)。株高、叶片数、穗长、穗码数、抽穗期5个性状随着日照的延长呈现出递增趋势。品种对谷子的10个性状有极显著影响(<0.01), 年份对千粒重以外的9个性状有极显著影响(<0.01)。光周期与品种互作效应对10个性状均有极显著影响(<0.01), 光周期与年份互作效应对抽穗期以外的9个性状有极显著影响(<0.01), 年份与品种互作效应对株高、叶片数、穗长、穗粗、穗重、穗粒重、穗码数、码粒数8个性状有极显著影响(<0.01)。160份谷子资源中对光周期表现中、低敏感性的材料没有明显的地域特性, 而对光周期表现强敏感性的材料主要是来自春谷区的农家品种。筛选出光周期极不敏感材料小早谷和极端敏感材料呼和浩特大毛谷、然谷、红钙谷、茄谷、二白谷等, 为选育谷子光周期钝感品种及开展光周期敏感性形成机理研究奠定了基础。
谷子; 光周期; 农艺性状; 抽穗期; 互作效应
谷子(Beauv.)属于短日照喜温作物, 对光周期和温度反应比较敏感是导致其生态适应性狭窄、生产上跨区域种植的主栽品种较少的重要原因。充分了解谷子主要性状对光周期反应敏感性的强弱, 合理评价谷子资源的光周期敏感性, 对开展谷子广生态适应性育种具有重要指导意义, 同时也为开展谷子光周期敏感性遗传规律研究以及谷子光周期敏感性相关性状QTL定位研究奠定基础。目前有关农艺性状光周期反应方面的研究多集中于模式作物大豆和禾本科作物玉米。对大豆的研究表明, 光周期反应存在于出苗至开花的全过程[1-2]; 不同生态类型大豆光周期不敏感性为北方春豆>南方春豆和黄淮春豆>黄淮夏豆>南方夏豆>南方秋豆[3]; 广适应性大豆品种中黄13的生育期性状和株高、主茎节数、分枝数、顶端花序荚数、单株粒数、粒重等性状光周期敏感性弱于中黄24和凤交66-12[4]; 光周期和温度对大豆的发育存在明显的互作, 随着温度的升高, 短日照促进大豆发育的效应有所增强, 随着日照的缩短, 高温加快发育的作用增强[5]。对玉米的研究表明, 针对抽雄期、开花期和总叶片数3个性状, 光周期反应的敏感程度为热带玉米>亚热带玉米>高原玉米>温带玉米[6]; 玉米植株性状的光周期敏感程度为雄穗分枝数>穗位高>棒三叶面积>叶片数>株高>雄穗长度>茎粗, 生育期性状的敏感程度依次为ASI>吐丝期>抽雄期>散粉期, 产量性状的敏感程度为穗重>穗粒重>行粒数>出粒率>穗粗>穗行数>千粒重>穗长[7]; 吐丝期、散粉期、ASI、全株叶片数、穗位下叶片数与品种的光周期反应有显著或极显著相关性, 穗位下叶片数和吐丝期可作为玉米光周期敏感性的鉴定指标[8]。谷子作为一个C4作物, 基因组比玉米、高粱小, 且全基因组序列已经由美国和中国测定[9-10], 这极大地方便了基因组水平研究的开展。目前虽然国内外开展了谷子光周期敏感性QTL定位方面的研究, 但都是以开花期作为光周期敏感性指标性状, 而对其他主要性状的光周期敏感性研究极少[11-13]。本研究利用方差分析比较3个不同日照长度生态区间10个性状的差异性, 揭示各性状光周期敏感性强弱, 综合评价160份谷子资源光周期敏感性, 旨在为开展谷子广生态适应性育种、光周期敏感性遗传学分析及QTL定位研究奠定基础。
1 材料与方法
1.1 谷子材料及种植方法
选用的160份谷子材料包括来自河南、河北、山东、山西、陕西、黑龙江、吉林、辽宁、新疆、内蒙古、甘肃、青海、宁夏、西藏等地区的品种资源及部分国外种质。2015年5月中旬至10月中旬、2016年5月中旬至10月中旬分别将材料种植于河南洛阳河南科技大学试验田(34°37¢N, 112°26¢E)、吉林市农业科学院试验田(42°31¢N, 125°40¢E)、吉林省农业科学院试验田(43°11¢N, 124°02¢E)。2015年11月中旬至2016年2月中旬、2016年11月中旬至2017年2月中旬将材料种植于海南乐东县九所镇(18°45¢N, 109°10¢E)。种植每品种(系) 1行, 行长2 m, 行距60 cm, 株距3~5 cm, 地两头设3行保护行, 按当地常规方法管理。表1是3个种植地的温度、降雨、日照时数信息, 吉林属于长日照地区, 河南属于中等日照地区, 海南属于短日照地区。
1.2 统计分析
1.2.1 谷子农艺性状测定 测定株高、叶片数、穗长、穗粗、穗码数、码粒数、穗重、穗粒重、千粒重、抽穗期10个性状。以每个品种(系)从出苗至植株超过50%抽穗的天数表示抽穗期, 每个品种(系)行中部10株测量其他各性状, 测量方法见贾小平等[14-15]。
1.2.2 数据分析 利用SPSS 19.0软件对10个性状进行多因素方差分析, 因素主要考虑光周期[长日照(吉林)、中日照(河南)、短日照(海南)], 年份(2015年、2016年)和品种(160个), 并用邓肯多重比较进行多重检验。选择抽穗期、叶片数、穗长、穗码数4个性状计算光周期相对敏感度(PS)。PS河南= (某性状河南测量值–某性状海南测量值)/某性状海南测量值; PS吉林= (某性状吉林测量值−某性状海南测量值)/某性状海南测量值。求出每个品种4个性状相对敏感度之和作为该品种的光周期敏感度, 数值越小代表该品种光周期敏感性越低。
2 结果与分析
2.1 谷子各性状的多因素方差分析
由表2可以看出, 谷子的株高(10.60~245.00 cm)、穗重(0.20~58.37 g)、穗粒重(0.03~46.06 g)、穗码数(3.00~306.00个)、码粒数(1.13~637.20粒)的测定值变异幅度比较大, 说明所选的谷子材料存在较广的遗传变异, 另外也说明年份、光周期条件对谷子农艺性状可能存在较大的影响。
由表3可以看出, 光周期对谷子的株高、叶片数、穗长、穗粗、穗重、穗粒重、穗码数、码粒数、抽穗期有极显著影响(<0.01), 对千粒重影响显著(<0.05)。品种对谷子的株高、叶片数、穗长、穗粗、穗重、穗粒重、穗码数、码粒数、抽穗期、千粒重有极显著影响(<0.01)。年份对谷子的株高、叶片数、穗长、穗粗、穗重、穗粒重、穗码数、码粒数、抽穗期有极显著影响(<0.01), 对千粒重无显著影响(>0.05)。光周期与品种互作效应对以上10个性状影响极显著(<0.01)。光周期与年份互作效应除了对抽穗期无显著影响外(>0.05), 对其他性状有极显著影响(<0.01)。年份与品种互作效应对抽穗期无显著影响(>0.05), 对千粒重影响显著(<0.05), 对其他农艺性状有极显著影响(<0.01)。
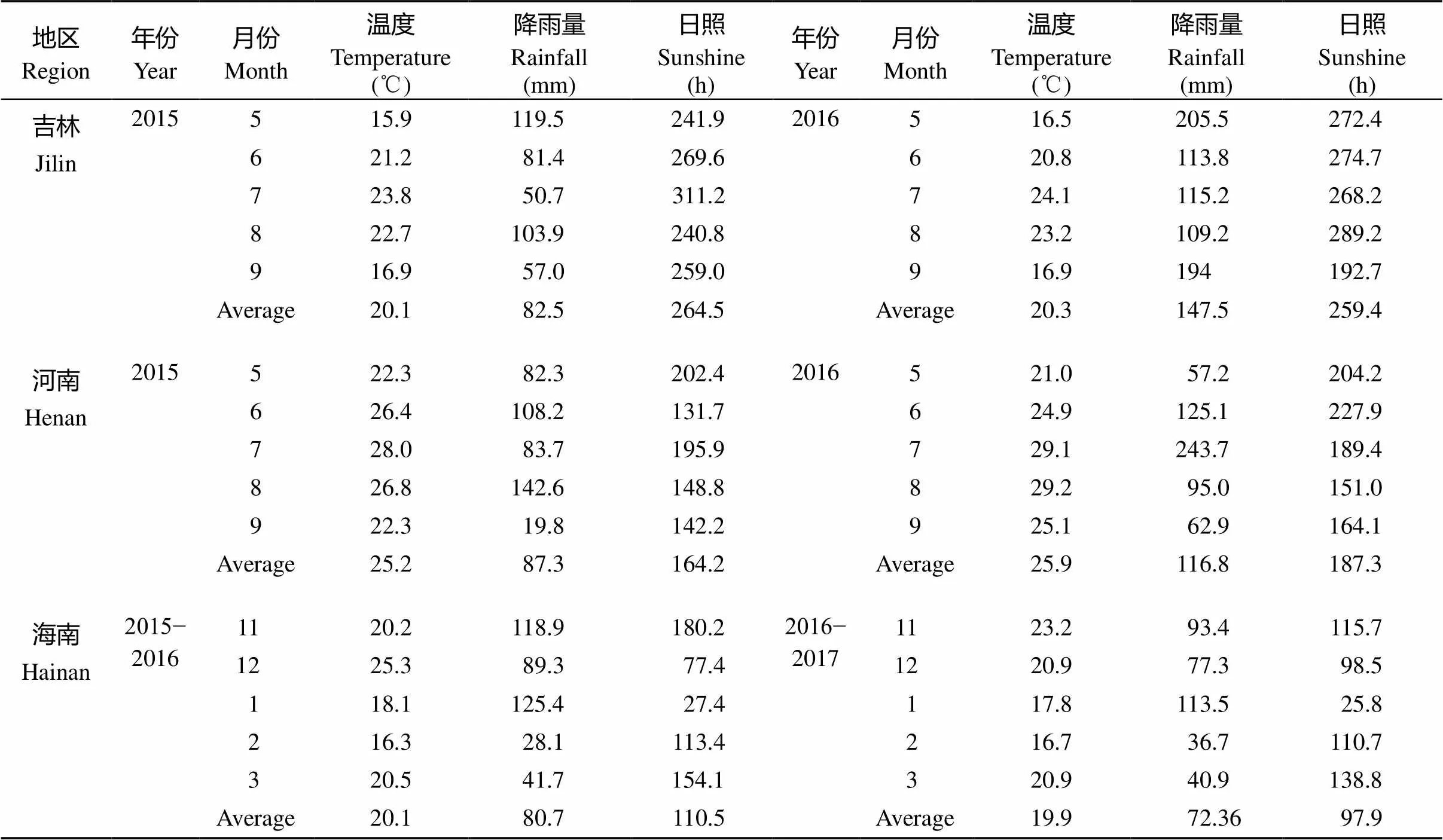
表1 不同地方的温度、降雨、日照时数

表2 谷子各性状的描述性统计量

表3 谷子各性状的多因素方差分析
(续表3)
因素Factor性状Trait平方和Sum of squares自由度df均方Mean squareF值F-value显著性P值SignificanceP-value 年份Year株高Plant height822174.381822174.388562.680.00 叶片数Number of leaf13075.28113075.2816900.060.00 穗长Panicle length3650.4013650.40547.170.00 穗粗Panicle diameter470.021470.02608.610.00 穗重Panicle weight9374.7919374.79586.300.00 穗粒重Grain weight per panicle2602.8112602.81234.880.00 穗码数Branch number per panicle41449.53141449.53183.180.00 码粒数Grain number per branch48833.25148833.25120.080.00 抽穗期Heading stage3330.3513330.35188.890.00 千粒重1000-grain weight1.0211.026.140.07 品种Variety株高Plant height2089521.592249328.2297.150.00 叶片数Number of leaf20789.9022492.81119.960.00 穗长Panicle length71359.45224318.5747.750.00 穗粗Panicle diameter9826.3422443.8756.800.00 穗重Panicle weight101343.40224452.4328.300.00 穗粒重Grain weight per panicle67909.59224303.1727.360.00 穗码数Branch number per panicle2321605.2922410364.3145.800.00 码粒数Grain number per branch2955378.7722413193.6632.440.00 抽穗期Heading stage32774.39165198.6311.270.00 千粒重1000-grain weight105.231650.643.850.00 光周期× 年份Photoperiod ×Year株高Plant height1445246.922722623.467525.890.00 叶片数Number of leaf20229.57210114.7813073.560.00 穗长Panicle length1124.722562.3684.290.00 穗粗Panicle diameter1032.202516.10668.280.00 穗重Panicle weight22351.24211175.62698.930.00 穗粒重Grain weight per panicle11356.6925678.34512.420.00 穗码数Branch number per panicle96841.08248420.54213.990.00 码粒数Grain number per branch205935.592102967.79253.190.00 抽穗期Heading stage18.3129.150.520.60 千粒重1000-grain weight6.2923.1418.960.00 光周期× 品种Photoperiod × Variety株高Plant height1266340.693973189.7833.220.00 叶片数Number of leaf6387.0039716.0920.790.00 穗长Panicle length62606.81397157.7023.640.00 穗粗Panicle diameter4102.8639710.3413.380.00 穗重Panicle weight81735.03397205.8812.880.00 穗粒重Grain weight per panicle58855.72397148.2513.380.00 穗码数Branch number per panicle1562876.893973936.7217.400.00 码粒数Grain number per branch1457601.443973671.549.030.00 抽穗期Heading stage12186.6830739.702.250.00 千粒重1000-grain weight70.393070.231.380.01
(续表3)
因素Factor性状Trait平方和Sum of squares自由度df均方Mean squareF值F-value显著性P值SignificanceP-value 年份 × 品种Year × Variety株高Plant height274965.272001374.8314.320.00 叶片数Number of leaf1927.472009.6412.460.00 穗长Panicle length9554.1020047.777.160.00 穗粗Panicle diameter1080.202005.406.990.00 穗重Panicle weight19149.7620095.755.990.00 穗粒重Grain weight per panicle13821.3520069.116.240.00 穗码数Branch number per panicle379445.962001897.238.390.00 码粒数Grain number per branch1234865.352006174.3315.180.00 抽穗期Heading stage3465.7415921.801.240.08 千粒重1000-grain weight36.431590.231.380.02 误差Error株高Plant height948949.07988396.02 叶片数Number of leaf7646.3098830.77 穗长Panicle length65934.2698836.67 穗粗Panicle diameter7632.4598830.77 穗重Panicle weight158025.71988315.99 穗粒重Grain weight per panicle109517.24988311.08 穗码数Branch number per panicle2236284.889883226.28 码粒数Grain number per branch4019193.469883406.68 抽穗期Heading stage3138.3617817.63 千粒重1000-grain weight29.511780.17 总计Total株高Plant height14200000.0010710 叶片数Number of leaf201143.4310710 穗长Panicle length557746.8510710 穗粗Panicle diameter47231.6210710 穗重Panicle weight728103.1010710 穗粒重Grain weight per panicle428015.8610710 穗码数Branch number per panicle12780000.0010710 码粒数Grain number per branch11510000.0010710 抽穗期Heading stage263089.69816 千粒重1000-grain weight326.93816
2.2 不同光周期环境对谷子各性状的影响
从图1可以看出, 株高、叶片数、穗长、穗码数均按短日照、中日照、长日照的顺序呈现递增趋势, 长日照条件下极显著高于中日照和短日照(<0.01), 中日照条件下的株高、叶片数、穗长、穗码数极显著高于短日照(<0.01); 穗重、穗粒重、穗粗、码粒数这4个性状的测定值在长日照条件最大, 在中日照条件下最小, 长日照条件下极显著高于中日照和短日照(<0.01), 中日照条件下极显著低于短日照(<0.01); 抽穗期按照短日照、中日照、长日照顺序呈现递增趋势, 长日照条件下极显著高于中日照和短日照(<0.01), 中日照条件下极显著高于短日照(<0.01); 千粒重以短日照条件下显著高于长日照(<0.05), 而长日照条件下显著高于中日照(<0.05)。
2.3 年份对谷子性状的影响
在受年份影响达到极显著水平的株高、叶片数、穗长、穗粗、穗重、穗粒重、穗码数、码粒数、抽穗期9个性状中, 年份对株高的影响较大, 2016年谷子株高为135.45 cm, 2015年谷子株高113.70 cm (图2)。
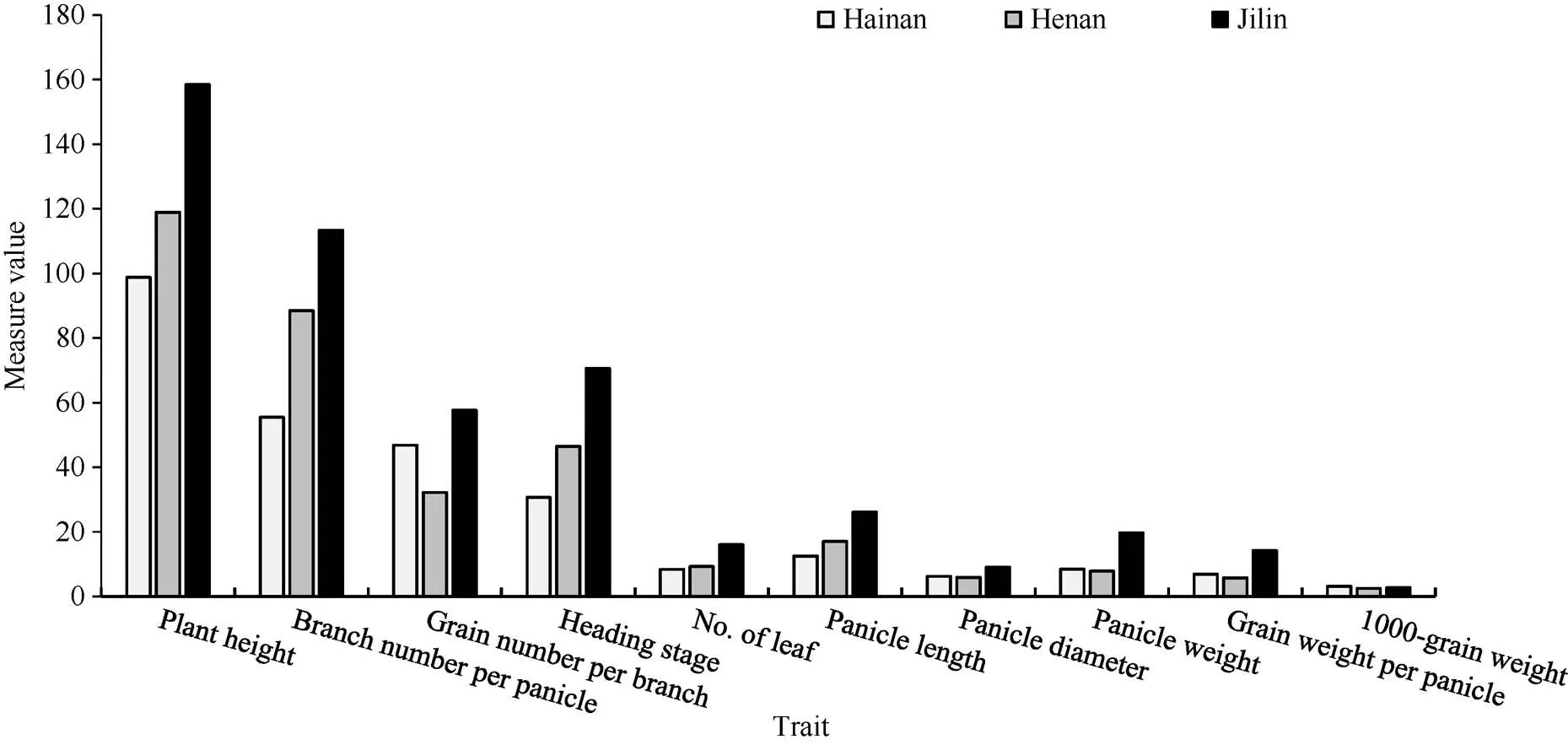
图1 不同光周期环境对谷子农艺性状的影响
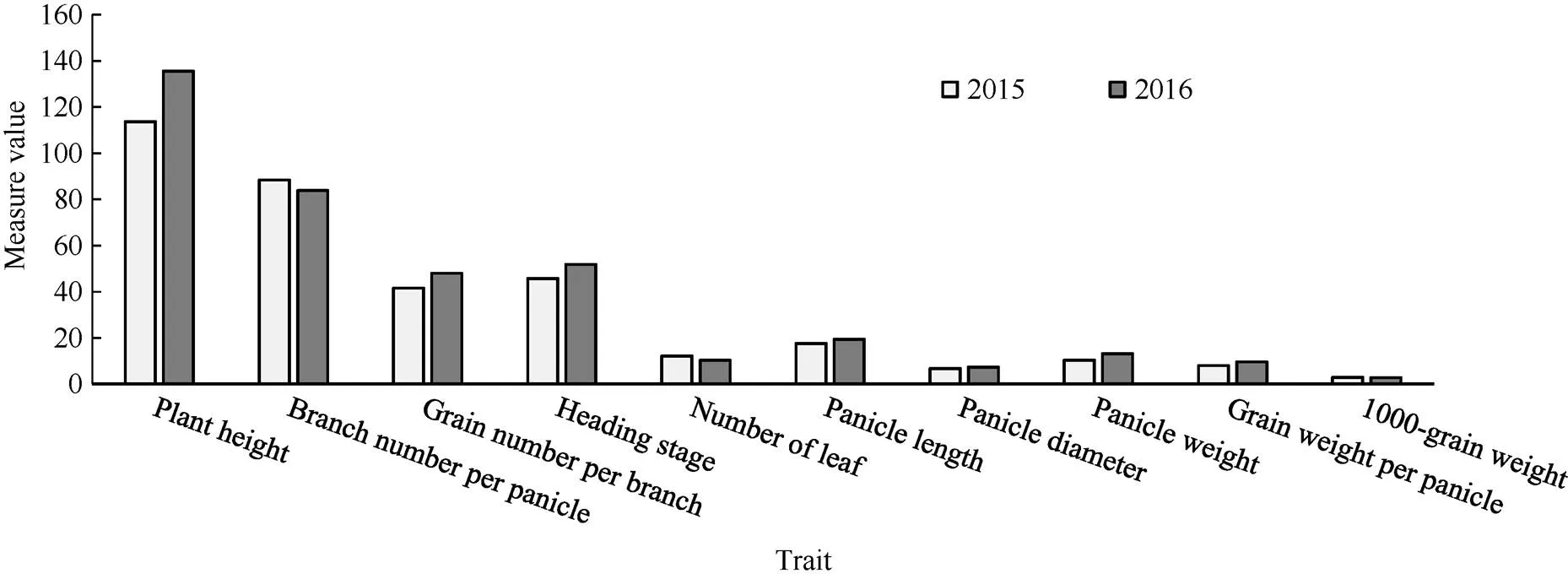
图2 2个年份间谷子农艺性状的比较
2.4 160份谷子资源的光周期敏感性评价
以对光周期敏感性反应较强的抽穗期、叶片数、穗长、穗码数4个性状的相对光周期敏感度之和作为评价指标, 评价160份谷子资源在河南、吉林两地的光周期敏感性强弱。在河南, 谷子资源光周期敏感度变化范围在-1.48~ 4.89之间, 其中小早谷和SET3/80两个品种对光周期反应特殊, 敏感度均小于0, 说明由海南短日照环境变为河南中日照环境各性状并未表现出增长趋势, 甚至略有降低。其余的品种可以划分为光周期低敏感材料(0.33~1.40)、光周期中等敏感材料(1.41~2.00)、光周期高度敏感材料(2.01~4.89)。从材料来源看, 光周期敏感度和地理来源并没有明显的对应关系, 中、低敏感度的材料来自全国各生态区, 还包含了部分国外种质, 无论育成品种还是地方品种, 都没有表现出明显的地域性, 一些来自河北、山东、吉林、山西、甘肃的地方品种如老绳头、白罗砂、齐头白、拔谷、钱串子、白杆白沙、叩根、红腿谷、小金苗等表现较低的光周期敏感性, 来自美国、德国、俄罗斯、南非的金德、Set64/82、K-3606、Red manna几个国外种质材料也表现较低的光周期敏感度, 这些资源可以作为中日照地区光周期钝感品种选育的亲本材料。生产上广泛种植的光温不敏感品种豫谷18号光周期敏感度为1.32, 在本研究中属于光周期低敏感度品种, 但排在靠后位置, 说明适度光周期敏感性是作物对新光温环境做出的适应性调节, 有利于增强其适应性。
在吉林, 谷子资源光周期敏感度变化范围在0.23~ 11.17之间, 根据光周期敏感度变化范围可以将所有谷子资源划分为光周期低敏感型(0.23~4.00)、光周期中敏感型(4.02~5.00)、光周期高敏感型(5.02~11.17)。在河南光周期敏感度为负值的2个品种(小早谷和SET3/80)均变为正值, 说明在长日照条件下这2个品种光周期敏感度有所增加, 但小早谷敏感度仍最低, 为0.23, 而SET3/80敏感度达到6.53, 由河南中度日照环境的不敏感变为吉林长日照环境的高度敏感。从品种来源分析, 发现和河南中日照条件一样, 低敏感度材料和生态环境没有明显的对应关系, 包括河南、吉林、河北、山西、辽宁、湖南、广西等地品种, 如郑315、公矮3号、15HN-138、晋谷35、铁7924、黄粟、大头糯。还有一些来自印度、日本、南非的国外种质, 如ISE-245、六十日、Red manna。比较河南和吉林两地, 发现一些材料均稳定表现出对光周期的低敏感性, 如小早谷、公矮3号、黄粟、15HN-138、Red manna、郑315、安4117、龙谷26等, 而豫谷18、532、矮88、铁谷4等虽然在河南和吉林两地都属于对光周期低敏感品种, 但在河南中日照环境的敏感度排序要比吉林长日照环境靠后。还有一些品种在河南光周期敏感度低, 到了吉林却变成对光周期高度敏感, 如SET3/80、K-3606、济叶冲4等。本研究还鉴定出一些河南、吉林两地均表现高度光周期敏感性品种资源, 如陇谷10号、黄毛谷、竹叶青、坝谷81、二白谷、茄谷、红钙谷、然谷、呼和浩特大毛谷等。从海南短日照环境到吉林长日照环境160份谷子资源光周期敏感度都有所增加(附表1)。
3 讨论
谷子属于短日照作物, 对光周期反应较敏感, 但是有关光周期对谷子主要农艺性状影响方面的报道极少, 有研究者设置不同短日照条件处理不同叶龄的谷子, 发现8 h短日照处理能够促进五叶期谷子出叶速度, 提早抽穗, 缩短育种进程[16]。在对谷子近缘种狗尾草光周期敏感性研究中发现, 长日照狗尾草A10表现明显的营养生长过剩和生育期推迟, 长日照敏感阶段要长于短日照敏感阶段, 狗尾草从出苗开始就进入光周期敏感期[17]。最近国外学者研究发现, 光周期条件对谷子开花期的影响最显著, 而对株高、生物量影响较小[18]。上述研究所用的谷子材料有限, 且调查的性状较少, 不能充分揭示谷子资源对光周期敏感性反应的总体表现和遗传差异。本研究选择代表不同生态区的160份谷子资源在日照长度存在明显差异的海南、河南、吉林3个生态区调查10个主要农艺性状, 发现光周期除对千粒重影响达到显著水平(<0.05)外, 对其他9个性状的影响均达到极显著水平(<0.01), 充分说明光周期对谷子的影响是全方位的, 包括了生育性状和产量性状。从海南短日照环境到河南中日照环境、吉林长日照环境株高、叶片数、穗长、抽穗期、穗码数均表现出规律性递增趋势, 与光周期的变化趋势表现出一致性, 因此这5个性状适合用来评价谷子的光周期敏感性。而已报道的谷子光周期敏感性QTL定位研究多以开花期作为评价指标[11-13], 谷子花器官较小, 在调查大量材料时, 存在工作量大、容易产生视觉误差等缺点, 而株高、抽穗期、叶片数、穗长则相对容易准确判断, 且省时省力, 在谷子资源光周期敏感性评价及谷子光周期敏感性QTL定位中可能会发挥更大作用。
目前对谷子资源光周期敏感性鉴定的研究还未见报道, 传统习惯上认为春谷材料光周期敏感性要强于夏谷, 本研究发现对光周期表现低敏感性或中度敏感性的谷子材料并没有明显的地域分布特点, 来自春谷区、夏谷区以及一些国外的种质均对光周期敏感性较弱, 而对光周期表现强敏感性的品种则多数来自春谷区, 且多为农家品种, 说明多数育成品种无论来自夏谷区还是春谷区, 经过选育明显降低了对光周期的敏感性, 春谷区、夏谷区的农家品种中也存在一些对光周期表现弱敏感性的资源, 尚待充分挖掘。生产上广泛种植的光温不敏感品种豫谷18号在本研究中虽然也属于光周期低敏感品种, 但是排在靠后的位置, 特别是在河南中日照环境, 事实上, 虽然豫谷18号从河南到吉林光周期敏感度值由1.32增加到3.38, 光周期敏感性增强了, 但是增加程度较低, 在河南豫谷18号光周期敏感度排在53位, 而在吉林则上升到21位, 和其他品种相比, 光周期敏感度反而降低了不少。说明在中日照条件下保持一定低敏感度而转至长日照条件下仍尽量降低光周期敏感度变化幅度可能是豫谷18号品种广生态适应性的原因。本研究发现了一些极端材料, 如小早谷, 在河南、吉林两地均表现出最弱的光周期敏感性, 生育期较短, 可以用来改良光周期高度敏感、在长日照条件下无法正常成熟的品种; 呼和浩特大毛谷、然谷、红钙谷、茄谷、谷子(来自青海)、二白谷、坝91-0130等材料在河南和吉林都表现出对光周期极端敏感性, 这些材料可以进一步用于研究谷子光周期敏感性形成的分子机制。
本研究所选的海南、河南、吉林3个环境日照长度差别较大, 适合开展谷子光周期敏感性研究, 但是温度、湿度、降水等其他环境因素也对农艺性状有一定影响, 同一个年份内3个环境的月均温海南与吉林接近, 为20~21℃左右, 但河南为25℃, 明显高于海南、吉林两地。降雨量在3个环境间和年份间存在较大差异, 吉林和河南2015年月降雨量明显低于2016年, 而海南则相反, 2015年明显高于河南和吉林, 2016年则是吉林最高, 河南次之, 海南最低。因此, 如何确定只对光周期敏感而受其他环境因子影响小的性状是评价谷子光周期敏感性和开展相关基因定位的关键。参考水稻、玉米、大豆的光周期敏感性研究成果, 发现水稻、大豆主要以生育期(抽穗期、开花期和成熟期)作为光周期敏感性指标, 定位了大量光周期途径的关键基因[19-21]; 玉米则以抽雄期、吐丝期和叶片数开展光周期敏感性研究, 获得了理想的结果[22-25]。借鉴水稻和玉米相关研究, 应以抽穗期和叶片数作为谷子光周期敏感性的主要评价指标, 结合穗长、穗码数2个性状开展研究应该会取得较好的结果。
附表 请见网络版: 1) 本刊网站http://zwxb.chinacrops.org/; 2) 中国知网http://www.cnki.net/; 3) 万方数据http://c.wanfangdata.com.cn/Periodical-zuowxb. aspx。
[1] 韩天富, 王金陵. 大豆开花后光周期反应的研究. 植物学报, 1995, 37: 863–869. Han T F, Wang J L. Studies on the post-flowering photoperiodic responses in soybean., 1995, 37: 863–869 (in Chinese with English abstract).
[2] Sun H B, Jia Z, Cao D, Jiang B J, Wu C X, Hou W S, Liu Y K, Fei Z H, Zhao D Z, Han T F., a soybean homolog of flowering locus T, is involved in flowering transition and maintenance., 2011, 6: e29238.
[3] 韩天富, 王金陵. 中国大豆不同生态类型开花至成熟期对光周期的反应. 作物学报, 1996, 22: 21–26. Han T F, Wang J L. A study on the responses of different ecotypes of Chinese soybeans to post-flowering photoperiod., 1996, 22: 21–26 (in Chinese with English abstract).
[4] 姜妍, 冷建田, 费志宏, 冯涛, 祖伟, 王连铮, 韩天富, 吴存祥. 广适应大豆品种中黄13的光周期反应. 大豆科学, 2009, 28: 377–381. Jiang Y, Leng J T, Fei Z H, Feng T, Zu W, Wang L Z, Han T F, Wu C X. Photoperiod responses of a widely-adapted soybean cultivar of Zhonghuang 13., 2009, 28: 377–381 (in Chinese with English abstract).
[5] 孙洪波. 大豆光温互作新模式的验证及PEBP家族基因的克隆和功能分析. 中国农业科学院研究生院博士后报告, 北京, 2008. Sun H B. Testify a New Soybean Model of Photo-thermal Interaction and Clone PEBP Family Genes. Postdoctoral Report of Graduate School of Chinese Academy of Agricultural Sciences, Beijing, China, 2008 (in Chinese with English abstract).
[6] 张凤路, Mugo S. 不同玉米种质对长光周期反应的初步研究. 玉米科学, 2001, 9(4): 54–56. Zhang F L, Mugo S. Primary study on the effect of long photo- period on different maize germplasm., 2001, 9(4): 54–56 (in Chinese with English abstract).
[7] 陈彦惠, 吴连成, 吴建宇, 席章营. 两种纬度生态条件下热带、亚热带玉米种质群体的鉴定. 中国农业科学, 2000, 33(增刊): 40–48. Chen Y H, Wu L C, Wu J Y, Xi Z Y. Identification of tropical, subtropical populations of maize in different ecological conditions of two latitudes., 2000, 33(suppl): 40–48 (in Chinese with English abstract).
[8] 梁文科. 热带温带玉米群体育种价值评估及光周期反应敏感性指标研究. 华中农业大学博士学位论文, 湖北武汉, 2008. Liang W K. Breeding Value Estimation and Photoperiod Sensitivity Index Study of Tropical, Temperate Maize Populations. PhD Dissertation of Huazhong Agricultural University, Wuhan, Hubei, China, 2008 (in Chinese with English abstract).
[9] Bennetzen J L, Schmutz J, Wang H, Percifield R, Hawkins J, Pontaroli A C, Estep M, Feng L, Vaughn J N, Grimwood J, Jenkins J, Barry K, Lindquist E, Hellsten U, Deshpande S, Wang X W, Wu X M, Therese M T, Triplett J, Yang X H, Ye C Y, Mauro- Herrera M, Wang L, Li P H, Sharma M, Sharma R, Ronald P C, Panaud O, Kellogg E A, Brutnell T P, Doust A N, Tuskan G A, Rokhsar D, Devos K M. Reference genome sequence of the model plant setaria., 2012, 30: 555–564.
[10] Zhang G Y, Liu X, Quan Z W, Cheng S F, Xu X, Pan S K, Xie M, Zeng P, Yue Z, Wang W L, Tao Y, Bian C, Han C L, Xia Q J, Peng X H, Cao R, Yang X H, Zhan D L, Hu J C, Zhang Y X, Li H N, Li H, Li N, Wang J Y, Wang C C, Wang R Y, Guo T, Cai Y J, Liu C Z, Xiang H T, Shi Q X, Huang P, Chen Q C, Li Y R, Wang J, Zhao Z H, Wang J. Genome sequence of foxtail millet () provides insights into grass evolution and biofuel potential., 2012, 30: 549–556.
[11] 谢丽莉. 谷子光周期敏感相关性状的QTL定位与分析. 河南农业大学硕士学位论文, 河南郑州, 2012.Xie L L. QTL Mapping and Analysis of the Photoperiod-sensitive Traits in Foxtail Millet. MS Thesis of Henan Agricultural University, Zhengzhou, Henan, China, 2012 (in Chinese with English abstract).
[12] Margarita M H,Wang X W, Barbier H, Brutnell T P, Devos K M, Doust A N. Genetic control and comparative genomic analysis of flowering time in Setaria (Poaceae)., 2013, 3: 283–295.
[13] Doust A N, Mauro-Herrera M, Hodgeand J G, Stromsk J. The C4model grass Setaria is a short day plant with secondary long day genetic regulation., 2017, 8: 1–10.
[14] 贾小平, 董普辉, 张红晓, 孔祥生. 不同谷子品种(系)生长发育及抗倒性分析. 河南农业科学, 2015, 44(8): 27–31. Jia X P, Dong P H, Zhang H X, Kong X S. Analysis of growth and development characteristics and lodging resistance of different foxtail millet cultivars (strains)., 2015, 44(8): 27–31 (in Chinese with English abstract).
[15] 贾小平, 董普辉, 张红晓, 全建章, 董志平. 谷子抗倒伏性和株高、穗部性状的相关性研究. 植物遗传资源学报, 2015, 16: 1188–1193. Jia X P, Dong P H, Zhang H X, Quan J Z, Dong Z P. Correlation study of lodging resistance and plant height, panicle traits in foxtail millet., 2015, 16: 1188–1193 (in Chinese with English abstract).
[16] 许寅生, 赵治海, 米连明, 孙玉山, 白有生. 不同短日照对不同叶龄谷子生长发育的影响. 张家口农专学报, 1999, 15(3): 19–22. Xu Y S, Zhao Z H, Mi L M, Sun Y S, Bai Y S. Effects of different leaf age and short-day on growth and development of foxtail millet., 1999, 15(3): 19–22 (in Chinese with English abstract).
[17] 谢丽莉, 吴连成, 库丽霞, 吴刘记, 张君, 王新涛, 陈彦惠. 玉米和狗尾草光周期敏感性的比较研究. 玉米科学, 2012, 20(2): 82–86.Xie L L, Wu L C, Ku L X, Wu L J, Zhang J, Wang X T, Chen Y H. Comparative study on the photoperiod-sensitive between maize and., 2012, 20(2): 82–86 (in Chinese with English abstract).
[18] Doust A N, Diao X M. Genetics and Genomics of Setaria. German: Springer-Verlag Publisher, 2017. pp 197–210.
[19] Doi K, Izawa T, Fuse T, Yamanouchi U, Kubo T, Shimatani Z, Yano M, Yoshimura A., a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of., 2004, 18: 926–936.
[20] Xue W Y, Xing Y Z, Weng X Y, Zhao Y, Tang W J, Wang L, Zhou H J, Yu S B, Xu C G, Li X H, Zhang Q F. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice., 2009, 40: 761–767.
[21] 夏正俊. 大豆光与生育期基因研究进展. 作物学报, 2013, 39: 571–579. Xia Z J. Research progresses on photoperiodic flowering and maturity genes in soybean (Merr.)., 2013, 39: 571–579 (in Chinese with English abstract).
[22] 孙朝辉. 玉米光周期敏感相关性状的QTL定位与分析. 河南农业大学硕士学位论文, 河南郑州, 2009.Sun Z H. QTL Mapping and Analysis for the Relevant Traits of Photoperiod Sensitivity in Maize. MS Thesis of Henan Agricultural University, Zhengzhou, Henan, China, 2009 (in Chinese with English abstract).
[23] 库丽霞, 孙朝辉, 王翠玲, 张君, 张伟强, 陈彦惠. 玉米光周期敏感相关性状发育动态QTL定位. 作物学报, 2010, 36: 602–611. Ku L X, Sun Z H, Wang C L, Zhang J, Zhang W Q, Chen Y H. QTL analysis of the photoperiod sensitivity-related traits at different developmental stages in maize (L.)., 2010, 36: 602–611 (in Chinese with English abstract).
[24] Yang Q, Li Z, Li W Q, Ku L X, Wang C, Ye J R, Li K, Yang N, Li Y P, Zhong T, Li J S, Chen Y H, Yan J B, Yang X H, Xu M L. CACTA-like transposable element in ZmCCT attenuated photoperiod sensitivity and accelerated the postdomestication spread of maize., 2013, 110: 16969–16974.
[25] Cui M, Jia B, Liu H H, Kan X, Zhang Y, Zhou R H, Li Z P, Yang L, Deng D X, Yin Z T. Genetic mapping of the leaf number above the primary ear and its relationship with plant height and flowering time in maize., 2017, 8: 1437, doi: 10.3389/fpls.2017.01437.
Effects of different photoperiod conditions on agronomic traits of foxtail millet
JIA Xiao-Ping1,*, QUAN Jian-Zhang2, WANG Yong-Fang2, DONG Zhi-Ping2, YUAN Xi-Lei1, ZHANG Bo1, andLI Jian-Feng1
1College of Agriculture, Henan University of Science and Technology, Luoyang 471023, Henan, China;2Institute of Millet, Hebei Academy of Agriculture and Forestry Sciences / National Millet Improvement Center, Shijiazhuang 050035, Hebei, China
One hundred and sixty foxtail millet materials were selected to investigate heading stage, plant height, leaf number, panicle length, panicle diameter, branch number per panicle, kernel number per branch, panicle weight, grain weight per panicle and 1000-grain weight under short-day (Hainan), middle-day (Henan), long-day (Jilin), three different photoperiod conditions in two consecutive years. The multiple factor variance analysis and multiple comparisons were performed by SPSS software (19.0 version) to explore the effects of photoperiod on the ten traits, and evaluate the photoperiod sensitivity of 160 foxtail millet materials. There were highly significant differences in heading stage, plant height, leaf number, panicle length, panicle diameter, branch number per panicle, kernel number per branch, panicle weight, grain weight per panicle (< 0.01), and significant difference in 1000-grain weight (< 0.05) among three photoperiod conditions. Five traits (heading stage, plant height, leaf number, panicle length, branch number per panicle) had increasing tendency with the prolonging of day time. The variety had extremely significant effect on all of the ten traits (< 0.01), and the year had extremely significant effect on the nine traits except for 1000-grain weight (< 0.01). The interaction between photoperiod and variety showed extremely significant effect on all of the ten traits (< 0.01), and that between photoperiod and year had extremely significant effect on the remaining nine traits except for heading stage (< 0.01). The interaction between year and variety showed extremely significant effect on plant height, leaf number, panicle length, panicle diameter, panicle weight, grain weight per panicle, branch number per panicle and kernel number per branch (< 0.01). The varieties with low or moderate sensitivity to photoperiod exhibited no clear regional characteristics. While most of the materials with strong sensitivity to photoperiod were landrances mainly from spring millet regions. The very insensitive material (Xiaozaogu) and the very strong sensitivity materials (Huhehaotedamaogu, Rangu, Honggaigu, Qiegu, Er’baigu and so on) screened out in this study provide a foundation for breeding cultivars insensitive to photoperiod and carrying out studies about formation mechanism of photoperiod sensitivity in foxtail millet.
foxtail millet; photoperiod; agronomic traits; heading stage; interaction effect
2018-10-13;
2019-04-15;
2019-04-23.
10.3724/SP.J.1006.2019.84128
贾小平, E-mail: jiaxiaoping2007@163.com
本研究由国家自然科学基金项目(31471569)资助。
This study was supported by the National Natural Science Foundation of China (31471569).
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20190422.1714.002.html
猜你喜欢
种子(2023年9期)2023-11-22 13:10:56
遗传(2023年9期)2023-09-25 09:31:44
农业科技通讯(2023年1期)2023-02-12 07:08:50
麦类作物学报(2022年7期)2022-08-09 08:04:46
麦类作物学报(2021年3期)2021-05-25 07:03:32
北京农学院学报(2020年1期)2020-03-09 07:18:28
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
江西农业学报(2016年5期)2016-06-21 05:45:20
江苏农业科学(2016年4期)2016-06-14 01:41:21
