
白叶1号白化过程中叶绿体蛋白质组差异分析
2019-06-20 07:13:26李勤程晓梅李永迪杨培迪黄建安刘仲华
茶叶科学 2019年3期
李勤,程晓梅,李永迪,杨培迪,黄建安,3*,刘仲华*
白叶1号白化过程中叶绿体蛋白质组差异分析
李勤1,2,3,程晓梅1,李永迪1,杨培迪4,黄建安1,3*,刘仲华1,2,3*
1. 湖南农业大学/茶学教育部重点实验室,湖南 长沙 410128;2. 国家植物功能成分利用工程技术研究中心,湖南 长沙 410128;3. 湖南省植物功能成分利用协同创新中心,湖南 长沙 410128;4. 湖南省农业科学院茶叶研究所,湖南 长沙 410128
白叶1号是一种温度敏感型白化茶树品种,叶绿体的变化是其产生阶段性白化现象的关键因素。本研究以白叶1号鲜叶叶绿体为研究对象,采用双向电泳、质谱鉴定结合生物信息学分析,研究阶段性白化过程中叶绿体蛋白的表达差异,探讨白叶1号阶段性白化现象的分子机制。结果表明,在白叶1号白化前期、白化期和复绿期叶绿体中分别识别726、748、718个蛋白质,其中差异表达的蛋白59个,质谱成功鉴定22个差异表达蛋白。生物信息学分析表明,差异表达蛋白直接或间接参与了光合作用、应激响应、核酸代谢、物质代谢和未知功能等,其中与光合作用相关的差异表达蛋白最多,占31.82%,表明阶段性白化现象可能与这些生理功能相关。通过荧光定量PCR分析发现,差异蛋白的基因表达与蛋白表达存在一定差异,这可能是由于蛋白质翻译后加工及修饰造成的。上述研究为进一步揭示白叶1号阶段性白化现象产生的分子机制奠定了理论基础。
白叶1号;阶段性白化;叶绿体蛋白质组;双向电泳
白叶1号是一种低温敏感型白化茶树品种,因其新梢阶段性白化现象和白化期较高的氨基酸含量而备受关注。多年田间观察发现,白叶1号阶段性白化现象与新梢萌芽期温度密切相关[1]。白化期间,其一芽二叶氨基酸含量比普通绿茶品种高1倍,茶多酚含量仅为普通绿茶品种的1/2[2]。研究表明,白叶1号叶色突变和高氨基酸含量的形成机制非常复杂,受温度、光照等外界环境因素和基因、蛋白变化等内在因素的共同影响[3-5]。我们前期研究发现,白叶1号阶段性白化过程中,叶绿体内囊体片层膜结构被破坏、叶绿素生物合成受阻、质体发育停滞,导致白化现象的产生。因此,我们推测叶绿体的变化是白叶1号阶段性白化现象的关键因素[6]。叶绿体是植物进行光合作用的重要细胞器,同时也是植物进行多种生理代谢所必需的场所。植物亚细胞结构分离技术快速发展,亚细胞蛋白质组学已成为植物蛋白质组学研究中的一个新热点[7]。张立明等[8]比较了不同茶树叶绿体分离及叶绿体蛋白质的提取方法,并采用双向电泳技术分离了约350个茶树叶绿体蛋白质,发现叶绿体蛋白质点主要集中于等电点3~5之间。但目前关于白叶1号阶段性白化过程中叶绿体蛋白质组的变化尚未见报道,阻碍了白叶1号白化现象的分子机制研究。
本研究以白叶1号阶段性白化过程中不同阶段鲜叶叶绿体为研究对象,采用双向电泳结合质谱技术研究阶段性白化过程中叶绿体蛋白质组的变化,为白叶1号阶段性白化现象形成的分子机制研究提供理论基础。
1 材料与方法
1.1 材料
白叶1号阶段性白化期间样品取自湖南省茶叶研究所高桥试验基地茶园。根据叶片颜色变化,采集阶段性白化过程中3个典型阶段叶片,包括白化前期、白化期和复绿期。取带叶枝条插入水中,迅速带回实验室,立即进行后续样品处理。
1.2 方法
1.2.1 叶绿体制备
叶绿体粗提物[9]:取25.0 g茶树鲜叶,然后加入200 mL预冷的叶绿体提取液(330 mmoL·L-1L-山梨醇,50 mmoL·L-1HEPES,2 mmoL·L-1EDTANa2,1 mmoL·L-1MgCl2,1 mmoL·L-1MnCl2,0.5 g·L-1牛血清白蛋白,0.05%-巯基乙醇,pH 8.0),于预冷的匀浆杯中低速匀浆3次,每次10 s;4层纱布过滤,滤液于4℃,500×条件下,离心10 min,弃沉淀;取上清液于4℃,2 000×条件下,离心10 min,弃上清;沉淀重悬于预冷的叶绿体提取液中,于4℃,2 000×条件下,离心10 min,弃上清;沉淀重悬于3 mL预冷的叶绿体洗涤缓冲液(330 mmoL·L-1L-山梨醇,50 mmoL·L-1HEPES,2 mmoL·L-1EDTANa2,1 mmoL·L-1MgCl2,1 mmoL·L-1MnCl2,0.05%-巯基乙醇,pH 8.0),即得叶绿体粗提液。
叶绿体纯化[10]:取3 mL叶绿体粗体液平铺于Percoll密度梯度液(3 mL 80% Percoll,3 mL 40% Percoll)上层,4℃,18 000×,离心60 min,用注射器吸取40%和80% Percoll密度梯度液之间的绿色条带至干净的离心管中,重悬于叶绿体洗涤缓冲液,4℃,12 000×离心10 min,沉淀即为纯化的完整叶绿体。
(3)叶绿体完整性与纯度检测:叶绿体完整性采用希尔反应法检测[11];叶绿体纯度采用过氧化氢酶活性和延胡索酸酶活性检测法测定[12]。
1.2.2 叶绿体蛋白质样品制备
采用三氯乙酸丙酮沉淀法[13],将纯化的叶绿体与10%的三氯乙酸丙酮溶液按1∶15(∶)涡旋混匀,–20℃放置至少4 h,沉淀蛋白;放置后将混合液在4℃,12 000×条件下离心10 min,弃上清液,向沉淀中加入1 mL预冷的100%丙酮(含0.07%的-巯基乙醇)涡旋混匀后继续离心10 min,至少重复洗涤沉淀3次;弃上清液,向沉淀中加入1 mL 80%预冷的丙酮(含0.07%的-巯基乙醇),混匀后离心,条件为4℃,12 000×,10 min,至少重复3次,洗至上清无色透明为止;弃去上清液,将打开管口的离心管置于冰上挥发丙酮,称重,所得沉淀即为叶绿体中的蛋白质,–20℃中保存备用。
将冷冻的蛋白沉淀按1∶15(∶)加入蛋白质裂解液,充分混匀后按体积比1∶50加入蛋白抑制剂,涡旋混匀置于冰上静置2 h,每半小时震荡混匀1次,之后超声破碎6次,总共1 min(超声5 s,停5 s),让蛋白质充分溶解,4℃,12 000×离心15 min,用移液枪吸取上清液到新的离心管中,即为叶绿体蛋白质样品,置于–20℃中保存备用。
1.2.3 双向电泳及质谱鉴定
参考Li等[6]的方法,采用17 cm,pH 4~7的IPG预制胶条,上样量350 μg,上样体积400 μL,50 V主动水化12 h。等电聚焦(IEF)条件:250 V缓慢升压1 h,500 V缓慢升压1 h,1 000 V缓慢升压3 h,10 000 V线性升压5 h,10 000 V快速升压至6 h,500 V保存。聚焦后IPG预制胶条经2次平衡,12%分离胶进行SDS-PAGE电泳分离。Blue silver胶体考染[12],图像扫描,利用Bio-Rad公司PDquest软件对凝胶进行图像分析。每组样品进行3次生物学重复试验。参考Li等[6]的方法,将差异表达的蛋白质点经胶内酶解、酶解肽段抽提、ZipTip脱盐、真空浓缩、质谱检测、数据库检索对差异表达蛋白质点进行鉴定。
1.2.4 荧光定量PCR分析
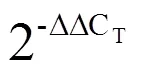
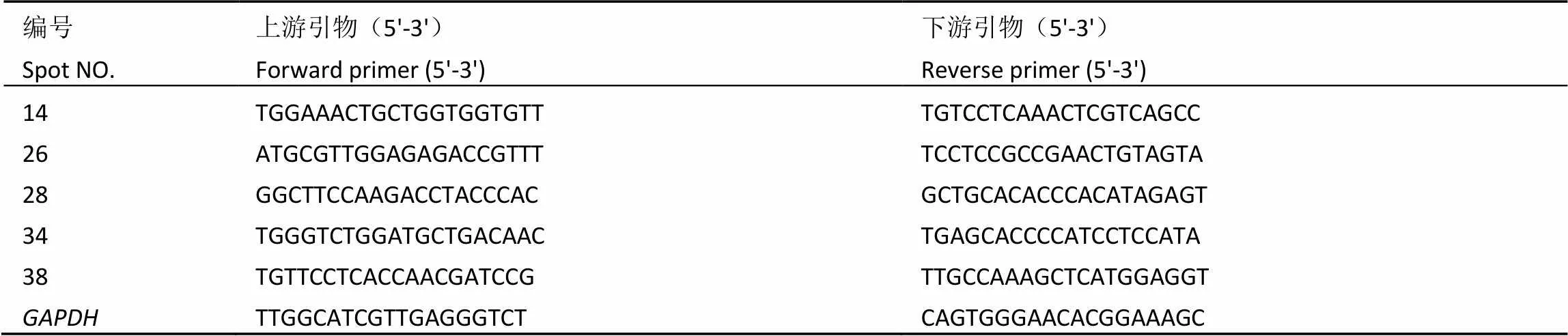
表1 实时荧光定量PCR引物序列
2 结果与分析
2.1 白叶1号叶绿体制备
采用Percoll密度梯度离心法对叶绿体进行分离纯化,梯度介质中从上至下形成两条绿色条带,纯化的完整叶绿体主要聚集在40%与80% Percoll分离介质梯度之间(图1,第二条绿色条带)。如表2所示,经Percoll密度梯度离心法纯化后,叶绿体得率为每100 g鲜叶可获得(8.31±0.03) mg叶绿体,叶绿体完整率为(73.68±0.54)%;线粒体和微粒体标志酶活性检测表明(图2),单位时间内纯化的叶绿体提取液过氧化氢酶和延胡索酸酶活性无显著性变化(>0.05),表明纯化的叶绿体提取液中无线粒体和微粒体污染,适合下一步试验要求。
2.2 叶绿体蛋白质组双向电泳
采用pH为4~7的IPG预制胶条和12%的SDS-PAGE胶对白叶1号白化前期、白化期及复绿期叶绿体蛋白质组样品进行双向电泳分离,染色后经PDQuest软件识别白化前期蛋白点726个,白化期748个,复绿期718个,其中丰度表达变化在1.5倍以上且Student’s-test检验差异显著(<0.05)、重复性好的差异蛋白质点共计59个(图3)。差异蛋白质分为4种表达类型:类型Ⅰ:在白化期下调表达的蛋白质点,共计39个,占总差异蛋白66.10%,其中7个蛋白质点在白化期未表达;类型Ⅱ:在白化期上调表达的蛋白质点,共计15个,占总差异蛋白25.42%,其中13个蛋白质点仅在白化期表达;类型Ⅲ:在白化前期、白化期和复绿期连续上调表达的蛋白质点,共计3个,占总差异蛋白5.08%;类型Ⅳ:在白化前期、白化期和复绿期连续下调表达的蛋白质点,共计2个,占总差异蛋白3.39%。

图1 叶绿体密度梯度离心

表2 叶绿体得率、完整性及纯度
注:图中数字表示差异蛋白质点编号,A:白化前期,B:白化期,C:复绿期
Note: The numbers represent the different proteins. A: Pre-albinistic stage, B: Albinistic stage, C: Regreening stage
图3 白叶1号阶段性返白过程中叶绿体蛋白质的双向电泳图谱
Fig. 3 Two-dimensional electrophoresis gel of separated chloroplast proteins in Baiye 1 during periodic albinism
2.3 差异蛋白质质谱鉴定及生物信息学分析
从上文所述的4种表达模式差异蛋白质中,选取25个不同分子质量、不同等电点、分离效果好、重复性好,且差异表达丰度大于1.5倍的差异蛋白点。经脱色、酶解、抽提和脱盐后,进行质谱鉴定。经数据库检索后,成功鉴定22个差异蛋白点,鉴定成功率为88.00%(表3)。根据差异蛋白质的功能,可将它们分为5类,分别为:与光合作用相关7个,占31.82%,白化期均呈现显著的下调表达,复绿期表达量又逐渐恢复;与应激响应相关5个,占22.73%;与核酸代谢相关3个,占13.64%;与物质代谢相关4个,占18.18%;未知功能的蛋白质3个,占13.64%。
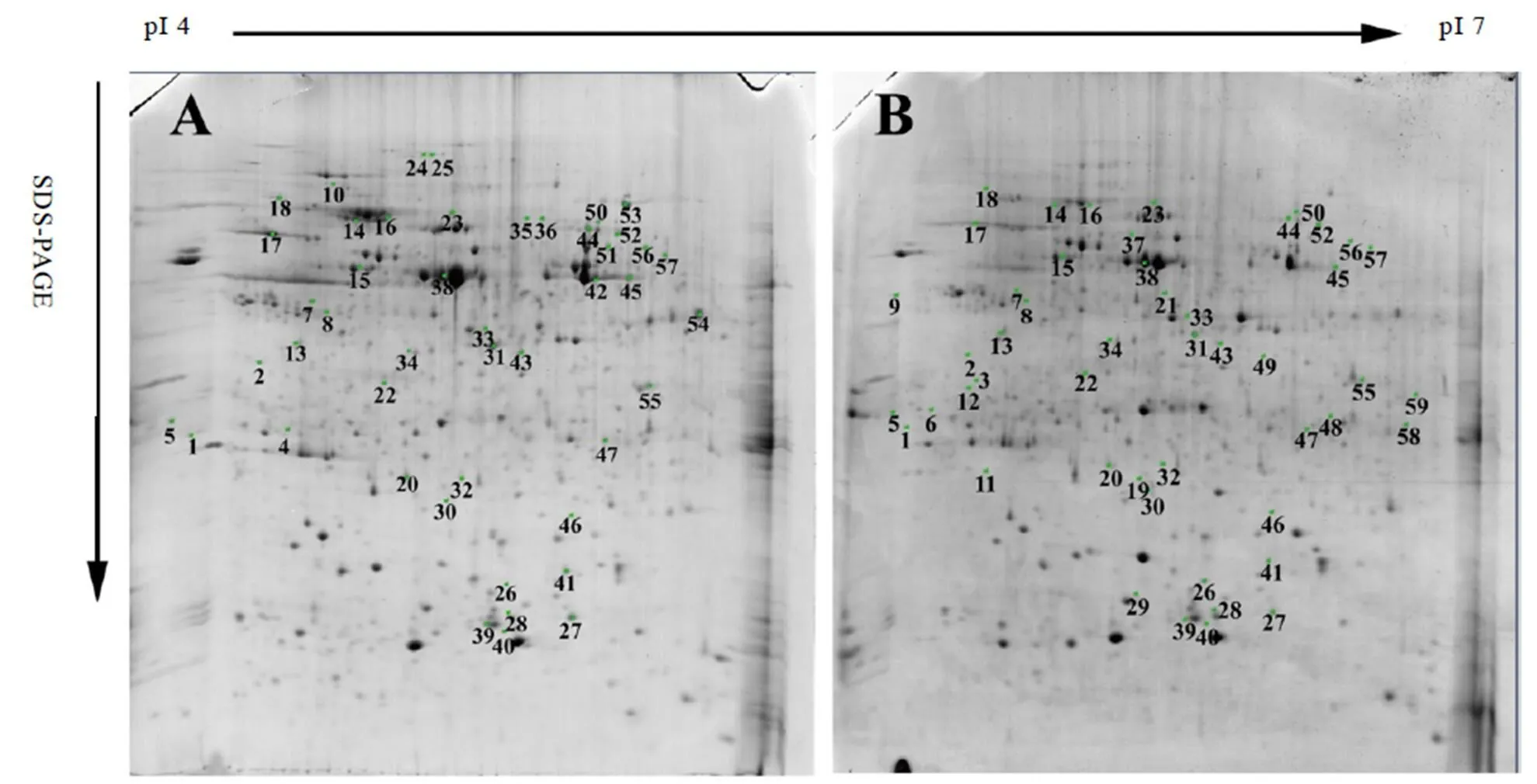
2.4 荧光定量PCR验证
从鉴定到的与光合作用、核酸代谢和应激响应功能相关的差异蛋白中挑选5个差异蛋白质对其基因表达情况进行分析。结果表明(图4),热激蛋白70、核酮糖1,5-二磷酸羧化酶/加氧酶大亚基、-腺苷甲硫氨酸合成酶和ATP合成酶CF1亚基基因表达情况与蛋白质表达情况一致。但真核起始因子5A4基因表达情况与蛋白质表达情况有一定差异;与白化期相比,真核起始因子5A4在白化前期基因水平为上调表达,而蛋白质水平为下调表达。蛋白质和基因表达的差异可能是由于蛋白质翻译的可变剪切,磷酸化和糖基化等翻译后修饰造成。
3 讨论
叶绿体是植物进行光合作用的一个重要细胞器。目前,对于叶绿体蛋白组学的研究主要分为叶绿体整体的蛋白质组学和叶绿体内不同组成部分的蛋白质组学,如叶绿体膜蛋白组学、叶绿体基质蛋白组学和叶绿体类囊体蛋白组学[15]。本研究采用蛋白质组学研究技术,研究了白叶1号阶段性白化过程中叶绿体蛋白质表达的变化,并对差异蛋白质鉴定及功能分析。
3.1 光合作用相关蛋白质
白叶1号阶段性白化过程中7个与光合作用相关的蛋白差异表达,其作用涉及光呼吸、光合磷酸化、光合电子传递等过程。核酮糖-l,5-二磷酸羧化酶/加氧酶(Rubisco)是植物卡尔文循环(C3循环)和光呼吸的关键酶[16],由8个大亚基(分子量为56 kD)和8个小亚基(分子量为14 kD)组成,叶绿体中Rubisco是在类囊体膜上合成的。Rubisco在C3循环中能够催化CO2与1,5-二磷酸核酮糖反应生成2分子3-磷酸甘油酸;在光呼吸中Rubisco可以催化O2与1,5-二磷酸核酮糖反应生成1分子3-磷酸甘油酸和1分子磷酸乙醇酸[16]。白叶1号阶段性白化过程中鉴定到2个差异表达的Rubisco大亚基蛋白质点(Spot 26,Spot 45),在白化期间发生了下调表达。有研究报道,在小麦白化阶段叶片蛋白质组分和含量显著降低,其中以Rubisco亚基的降低幅度最大[17]。翁晓燕等[18]在白化水稻突变体阶段性白化过程中发现,白化苗类囊体片层结构缺乏,Rubisco含量显著降低;随叶片转绿,类囊体片层结构逐渐恢复并增多,Rubisco含量也随之显著增加,推测白化期间Rubisco含量显著降低与叶绿体类囊体片层结构缺失有关。我们前期研究也表明,白叶1号在白化期间叶绿体类囊体膜结构被破坏[6]。因此,白化期间叶绿体Rubisco含量显著降低可能与类囊体膜片层结构缺失,导致Rubisco合成受阻有关。能量传递是光合作用的重要组成部分,而ATP合成酶是能量代谢的关键酶,其参与光合作用中的光合磷酸化及氧化磷酸化过程,将光能转化为化学能[19]。叶绿体中的ATP合酶位于类囊体膜上,由CF0和CF1两部分组成[20]。白叶1号阶段性白化过程中,ATP合酶(Spot 32,Spot 38,Spot 42)表达量随着叶片的白化而呈现下降趋势,当叶片颜色逐渐复绿,该酶的表达量逐渐增强。有文献报道,叶绿体类囊体膜上若除去CF1的亚基,ATP合酶将失去催化ATP合成的能力,当添加外源亚基,ATP合成能力就可得到恢复[21]。毛娟等[22]研究也表明,植物ATP合成酶的缺失会导致植物叶色黄化,光合电子传递受到影响。因此,推测白叶1号阶段性白化过程中,由于叶绿体类囊体膜结构破坏,影响了ATP合酶组装,导致叶色出现白化,ATP合成受阻;同时,C3受到抑制,使净光合速率下降;后期随着叶绿体类囊体膜结构逐渐恢复,ATP合成酶组装随之恢复,光合速率逐渐增强。细胞色素b5是生物体内氧化反应的电子传递组分,还能够促进膜的合成[23]。本研究中,细胞色素b5(Spot 27,Spot 39)在白叶1号白化期表达量降低,影响电子传递,阻碍了白化期叶片光合作用。
注:Spot 14:热激蛋白70,Spot 26:核酮糖1,5-二磷酸羧化酶/加氧酶大亚基,Spot 28:真核起始因子5A4,Spot 34:S-腺苷甲硫氨酸合成酶,Spot 38:ATP合成酶CF1β亚基。*表示P<0.05,**表示P<0.01
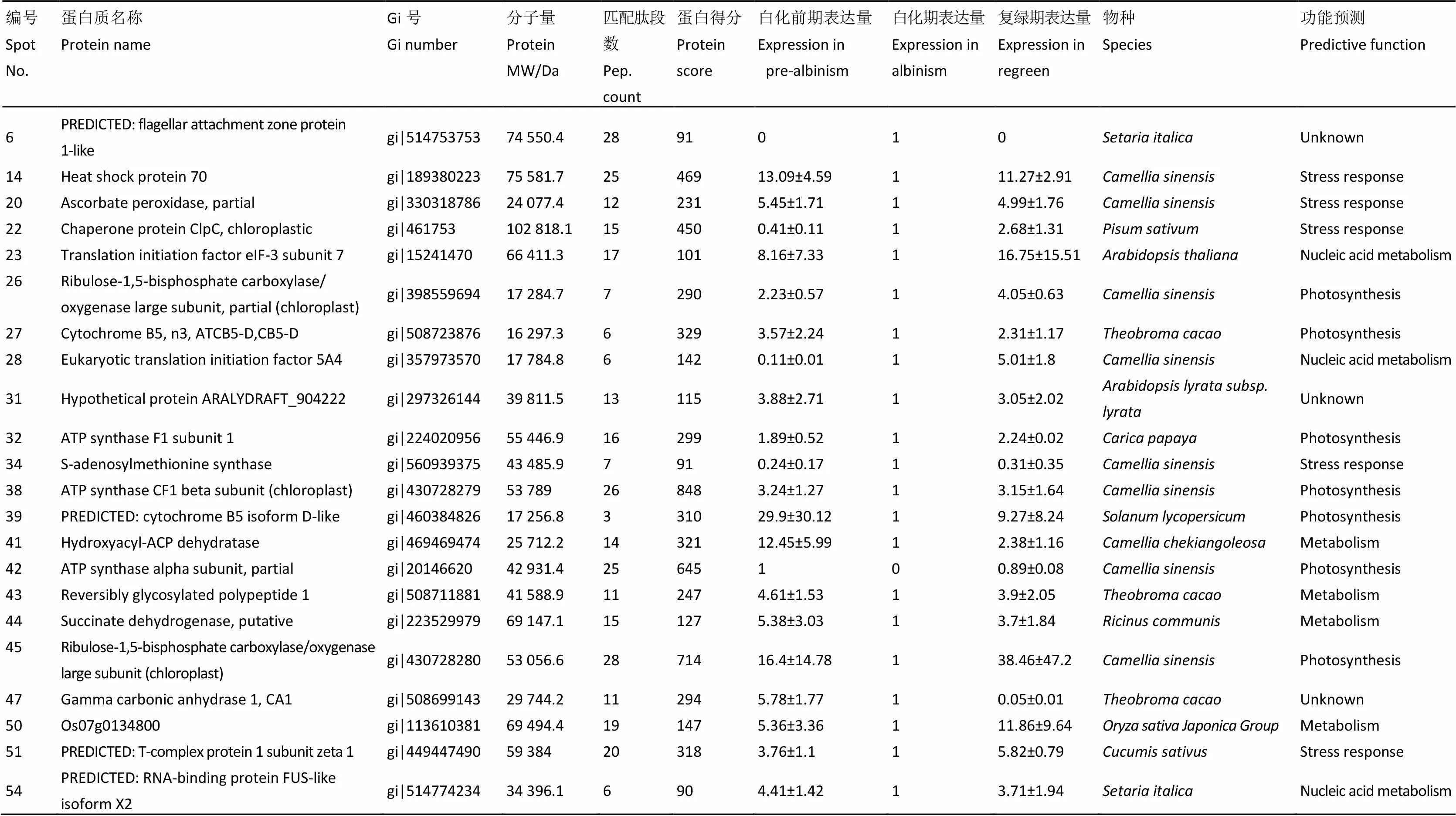
表3 叶绿体差异蛋白点质谱鉴定结果
3.2 核酸代谢相关蛋白质
叶绿体是含有自身遗传物质的一类细胞器,其基因的表达受转录水平的调控。白叶1号阶段性白化期间3个差异表达蛋白与核酸代谢及调控相关。叶绿体RNA结合蛋白(Spot 54)定位于叶绿体,与叶绿体基因转录后调控密切相关,它与细胞核的RNA结合蛋白具有相似的功能,包括维持RNA稳定、RNA翻译后调控等过程[24]。白叶1号阶段性白化期间,RNA结合蛋白表达量在白化期显著降低,随着叶片逐渐转绿其表达量又呈上升状态,推测可能与白化期叶绿体发育停滞有关。有报道指出,叶绿体RNA结合蛋白与叶绿体中核糖体复合体的装配及叶片的发育关系密切[25],叶绿体RNA结合蛋白表达水平能对光合作用产生调控作用,且叶绿体中mRNAs的稳定性会因叶绿体发育的变化而变化[26]。我们前期研究表明,白叶1号白化期叶片光合速率显著降低,可能也与白化期RNA结合蛋白表达量降低抑制了叶绿体核糖体组装有关[27]。真核起始因子(eIF)在真核生物翻译过程中具有RNA解旋酶的作用。白叶1号阶段性白化期间,eIF(Spot 23,Spot 28)表达量在白化期显著降低可能与白化期叶绿体基因翻译减少,蛋白质合成受到抑制有关。
3.3 应激响应相关蛋白质
植物在遭受逆境胁迫时,会产生应激响应,增强自身对胁迫环境的适应性。热激蛋白(HSP)在逆境胁迫下大量表达,以增强植物的抗逆性,根据分子质量可分为Hsp100、Hsp90、Hsp70、Hsp60、Hsp40等[28]。Hsp70主要通过参与机体内蛋白质的合成、折叠及组装和协助降解变性蛋白等多种过程来维持胁迫环境下植物机体内环境稳定,增强适应性[29]。Hsp70蛋白存在于植物叶绿体膜和基质,有利于维持叶绿体正常结构及叶绿体形成,是叶绿体发育的必要条件。白叶1号阶段性白化期间,Hsp70(Spot 14)表达量在白化期显著降低,复绿期逐渐升高,可能与白化期间叶绿体发育停滞,复绿期间叶绿体重新形成有关,在白化小麦突变株中也获得相同结果[30]。
S-腺苷甲硫氨酸合成酶(SAMS)催化S-腺苷甲硫氨酸(SAM)合成,SAM是合成乙烯和多胺的甲基供体。乙烯和多胺都参与了植物的抗逆反应,因而SAMS在植物抗逆中可能发挥一定的作用。本研究中,白叶1号白化期,SAMS(Spot 34)表达量显著提高,可能是因白化期间叶绿体被破坏,茶树产生了抗逆反应所致。
H2O2是叶绿体中光合电子传递和酶学反应的重要活性氧,但过量的H2O2会引起细胞损伤,因此应及时清除H2O2,维持叶绿体正常的光合作用。逆境环境下植物会产生过量的活性氧,损伤植物细胞。抗坏血酸过氧化物酶(APX)是植物体内清除H2O2的关键酶,可分为叶绿体基质APX(sAPX)、类囊体APX(tAPX)、微体APX(mbAPX)和胞质APX(cAPX)[31]。有研究报道,菠菜完整叶绿体基质中含有一种以光化还原剂为电子供体的过氧化物酶,即APX,但在不完整的叶绿体中APX活性很低[32]。也有报道,抗坏血酸过氧化物酶存在于白化突变苗中,而在绿苗中缺失[33],上述结果可能是由于不同植物中APX对各种环境因素的敏感性不一致而造成的[31]。在白叶1号白化期,APX(Spot 20)表达量的降低推测是由于叶绿体发育停滞、成熟叶绿体结构不完整导致APX表达量降低,随着复绿期叶绿体结构逐渐恢复,APX表达量也有所提高。
植物光合作用时,叶绿体中蛋白质常受到各种损伤而丧失功能,叶绿体中分子伴侣蛋白负责受损蛋白的重新折叠,蛋白酶负责清除不能修复的蛋白,蛋白酶对叶绿体的发育及结构维持具有重要作用[34]。ATP依赖型Clp蛋白酶主要位于叶绿体基质,能降解多种基质蛋白,在叶绿体发育过程中起着重要作用。Sjogren等[35]报道,抑制拟南芥ClpP6表达后,幼叶呈黄化表型,其底物主要是调控蛋白质合成、折叠等功能的基质蛋白。白叶1号白化期间,ATP依赖型Clp蛋白酶(Spot 22)表达在白化期有一定程度的上升,进入复绿阶段进一步显著上升。推测由于白化期,叶绿体结构破坏,产生功能大量变性蛋白质,诱导ATP依赖型Clp蛋白酶表达,但由于叶绿体发育停滞仅有少量叶绿体存在,因而ATP依赖型Clp蛋白酶表达量提高有限;随着叶片逐渐复绿,叶绿体结构和发育恢复正常,光合作用逐渐增强,ATP依赖型Clp蛋白酶表达量显著提升。
3.4 脂质代谢相关蛋白质
脂肪酸是植物体的重要组分,在生物膜合成、信号的转导及能量储存等方面具有重要作用。羟脂酰-ACP脱水酶(HAD)参与Ⅱ型脂肪酸合成代谢,主要作用是催化可逆反应羟酰-ACP脱水生成烯酰-ACP[36]。在植物体中饱和脂肪酸合成途径大部分发生在叶绿体中,Ferro等[37]利用液质联用技术在拟南芥中鉴定到与叶绿体脂质代谢相关的膜蛋白。白叶1号阶段性白化期间,HAD(Spot 41)在白化期含量显著降低,可能与叶绿体膜结构破坏有关,但HAD与叶片的白化是否有关仍需进一步研究。
叶绿体是植物光合作用必需场所,白叶1号阶段性白化过程中光合作用发生了显著变化,研究白叶1号阶段性白化过程中叶绿体蛋白质组的变化,对了解和发现白叶1号白化期叶色突变和高氨基酸含量具有重要意义。本研究通过双向电泳结合质谱技术在亚细胞和蛋白质水平上鉴定了白化过程中部分差异表达的叶绿体蛋白质,发现这些差异表达蛋白质直接或间接参与光合作用、能量代谢、应激响应、脂质代谢等生理生化过程,其中与光合作用相关的差异表达蛋白最多,表明这些生理生化过程的变化可能与白叶1号阶段性白化现象有关。这些结果为阐明白叶1号阶段性白化现象的分子机制提供了一定的理论基础。
[1] 李素芳. 安吉白茶返白机理的研究[J]. 中国计量学院学报, 2002, 13(3): 214-217.
[2] 杨普香, 李文金, 聂樟清, 等. 安吉白茶茶多酚和氨基酸含量初探[J]. 蚕桑茶叶通讯, 2007(5): 33-34.
[3] 成浩, 李素芳, 陈明, 等. 安吉白茶特异性状的生理生化本质[J]. 茶叶科学, 1999, 19(2): 87-92.
[4] 曾超珍, 刘仲华. 安吉白茶阶段性白化机理的研究进展[J]. 分子植物育种, 2015, 13(12): 2905-2911.
[5] 卢翠, 沈程文. 茶树白化变异研究进展[J]. 茶叶科学, 2016, 36(5): 445-451.
[6] Li Q, Huang J, Liu S, et al. Proteomic analysis of young leaves at three developmental stages in an albino tea cultivar [J]. Proteomecience, 2011, 9(1): 44. https://doi.org/10.118 6/1477-5956-9-44.
[7] 贺庭琪, 郭安平, 杜伟, 等. 植物叶绿体蛋白质组学研究进展[J]. 热带作物学报, 2011, 32(11): 2196-2203.
[8] 张立明, 刘亚军, 王云生, 等. 茶树叶绿体及其蛋白的分离研究[J]. 激光生物学报, 2011, 20(6): 802-808.
[9] 肖龙, 张彩霞, 宗泽冉, 等. 苹果叶片应答轮纹病菌胁迫的叶绿体蛋白质组学分析[J]. 果树学报, 2016, 33(11): 1357-1366.
[10] 肖龙, 张彩霞, 张利义, 等. 苹果叶片叶绿体分离及其蛋白提取、双向电泳方法的优化[J]. 果树学报, 2016, 33(6): 752-761.
[11] Lydia Macias-Rubalcava M, Claudia Garcia-Mendez M, King-Diaz B, et al. Effect of phytotoxic secondary metabolites and semisynthetic compounds from endophytic fungus Xylaria feejeensis strain SM3e-1b on spinach chloroplast photosynthesis [J]. Journal of Photochemistry and Photobiology B: Biology, 2017, 166: 35-43.
[12] Kley J, Heil M, Muck A, et al. Isolating intact chloroplasts from small Arabidopsis samples for proteomic studies [J]. Analytical Biochemistry, 2010, 398(2): 198-202.
[13] 李勤, 黄建安, 刘硕谦, 等. 茶树蛋白质双向电泳样品制备技术研究[J]. 茶叶科学, 2011, 31(3): 173-178.
[14] Chomczynski P, Mackey K. Short technical reports. Modification of the TRI reagent procedure for isolation of RNA from polysaccharide- and proteoglycan-rich sources [J]. BioTechniques, 1995, 19(6): 942-945.
[15] Baginsky S, Gruissem W. Chloroplast proteomics: potentials and challenges [J]. Journal of Experimental Botany, 2004, 55(400): 1213-1220.
[16] Spreitzer RJ, Salvucci ME. Rubisco: Structure, regulatory interactions, and possibilities for a better enzyme [J]. Annual Review of Plant Biology, 2002, 53: 449-475.
[17] 苏小静, 汪沛洪, 王永吉. 小麦突变体返白系返白机理研究-Ⅲ.返白阶段叶蛋白质、游离氨基酸变化规律的分析[J]. 西北农林科技大学学报(自然科学版), 1990, 18(4): 16-22.
[18] 翁晓燕, 蒋德安, 陆庆. 水稻转绿型白化突变系W25转绿过程中Rubisco、Rubisco活化酶活性与光合速率的变化[J]. 植物生理学报, 2000, 26(3): 213-225.
[19] Igamberdiev AU, Kleczkowski LA. Optimization of ATP synthase function in mitochondria and chloroplasts via the adenylate kinase equilibrium [J]. Frontiers in Plant Science, 2015, 6: 10. https://doi.org/10.3389/fpls.2015.00010.
[20] Rast A, Heinz S, Nickelsen J. Biogenesis of thylakoid membranes [J]. Biochimica Et Biophysica Acta-Bioenergetics, 2015, 1847(9): 821-830.
[21] 董慧. 叶绿体ATP合酶的活性调节与亚基的定点突变[D]. 上海: 中国科学院研究生院(上海生命科学研究院), 2005.
[22] 毛娟, 迟伟, 马今方, 等. 拟南芥PAB1蛋白参与叶绿体ATP合酶复合物调控机制的研究[J]. 生物物理学报, 2009, 25(S1): 242.
[23] 胡光珍. 香蒲细胞色素b5基因的克隆和功能研究[D]. 上海: 华东师范大学, 2005.
[24] 乌凤章, 王柏臣, 杨传平. 白桦叶绿体RNA结合蛋白的蛋白质组学分析(英文)[J]. 中国生物化学与分子生物学报, 2013, 29(2): 144-150.
[25] Dupont FM. Metabolic pathways of the wheat () endosperm amyloplast revealed by proteomics [J]. Bmc Plant Biology, 2008, 8(1): 39. https://doi.org/10.1186/ 1471-2229-8-39.
[26] 张在宝, 李婉杰, 李九丽, 等. 植物RNA结合蛋白研究进展[J]. 中国农业科学, 2018, 51(21): 4007-4019.
[27] 程晓梅. 安吉白茶阶段性返白过程中叶绿体蛋白质组学研究[D]. 长沙: 湖南农业大学, 2015.
[28] 栗振义, 龙瑞才, 张铁军, 等. 植物热激蛋白研究进展[J]. 生物技术通报, 2016, 32(2): 7-13.
[29] 王明强, 张道远. 植物热激蛋白70基因家族及其生物学功能研究进展[J]. 基因组学与应用生物学, 2015, 34(2): 421-428.
[30] 侯典云. 小麦返白系叶绿体基因组分析及叶绿体超微结构和差异表达蛋白质研究[D]. 杨凌: 西北农林科技大学, 2009.
[31] 孙卫红, 王伟青, 孟庆伟. 植物抗坏血酸过氧化物酶的作用机制、酶学及分子特性[J]. 植物生理学通讯, 2005, 41(2): 143-147.
[32] 孙云. 茶叶抗坏血酸过氧化物酶(APX)的生理学与分子生物学研究[D]. 福州: 福建农林大学, 2009.
[33] 董红霞. 水稻苗期叶色突变体的蛋白质组分析[D]. 武汉: 华中农业大学, 2010.
[34] Langer T. AAA proteases: cellular machines for degrading membrane proteins [J]. Trends in Biochemical Sciences, 2000, 25(5): 247-251.
[35] Sjogren LL, Stanne TM, Zheng B, et al. Structural and functional insights into the chloroplast ATP-dependent Clp protease in[J]. The Plant cell, 2006, 18(10): 2635-2649.
[36] Lu Y-J, Zhang Y-M, Rock CO. Product diversity and regulation of type II fatty acid synthases [J]. Biochemistry and Cell Biology-Biochimie et Biologie Cellulaire, 2004, 82(1): 145-155.
[37] Ferro M, Salvi D, Brugiere S, et al. Proteomics of the chloroplast envelope membranes from[J]. Molecular & Cellular Proteomics: MCP, 2003, 2(5): 325-345.
Analysis of the Chloroplast Proteome Difference of ‘Baiye 1’ [(L.) O Kuntze] during Periodic Albinism
LI Qin1,2,3, CHENG Xiaomei1, LI Yongdi1, YANG Peidi4, HUANG Jian'an1,3*, LIU Zhonghua1,2,3*
1. Tea Key Lab of the Ministry of National Teaching of Hunan Agricultural University, Changsha 410128, China;2. National Research Center of Engineering Technology for Utilization of Botanical Functional Ingredients, Changsha 410128, China;3. Hunan Co-Innovation Center for Utilization of Botanical Functional Ingredients, Changsha 410128, China;4. Tea Research Institute, Hunan Academy of Agriculture Sciences, Changsha 410128, China
‘Baiye 1’ is a kind of temperature sensitive tea cultivar. The change of chloroplast is the key factor for the periodic albinism of ‘Baiye 1’. To understand the mechanism of periodic albinism of ‘Baiye 1’, two dimensional electrophoresis (2-DE) and mass spectrometry was adopted to separate and identify the chloroplast proteins, which were significantly changed during the three developmental periods. The results show that 726, 748 and 718 protein spots were separated at the pre-albinistic, albinistic and regreen stages, respectively. The expression levels of 59 protein spots varied markedly during the three development stages. A total of 22 protein spots were successfully identified by MS, which were involved in photosynthesis, stress response, metabolism of nucleic acid, substance metabolism and unknown function. Photosynthetic proteins were the most affected proteins, which account for 31.82% in the significantly changed proteins. These results indicate that these physiological processes might playcrucial roles in the periodic albinism. The gene expression profiles of the differentially expressed proteins were alsoverified by real-time PCR analysis. The results show that the expressions of genes and proteins were not consistent, which might be related to the protein processing and post-modification. These results provide a theoretical basis for understanding the molecular mechanism of periodic albinism in ‘Baiye 1’.
Baiye 1, periodic albinism, chloroplast proteome, 2-DE
S571.1
A
1000-369X(2019)03-325-10
2018-11-05
2019-01-01
国家自然科学基金(31200522)、湖南省教育厅科研项目(15B116)、湖南省财政厅科技专项(湘财教指[2016]175号)、湖南农业大学校青年基金项目(15QN28)
李勤,男,讲师,主要从事茶叶生物化学和种质资源创新方面的研究,E-mail: liqinvip@126.com。*通信作者
猜你喜欢
热带农业科学(2020年7期)2020-08-31 07:10:08
环境与生活(2020年4期)2020-02-19 04:41:50
今日农业(2019年12期)2019-08-15 00:56:32
中国外汇(2019年22期)2019-05-21 03:15:06
生活PLUS(2018年6期)2018-05-14 11:55:31
广西林业科学(2016年1期)2016-03-20 05:33:01
茶叶(2015年3期)2015-12-13 06:31:06
人间(2015年21期)2015-03-11 15:24:49
中国茶叶加工(2015年3期)2015-02-27 07:55:25
食品科学(2013年6期)2013-03-11 18:20:13
