
敲低钙调磷酸酶结合蛋白1引起肾小管上皮细胞线粒体损伤
2019-06-18 02:19:08吴汪丽温跃强
广州医药 2019年3期
吴汪丽 温跃强
广州医科大学附属第二医院 (广州 510260)
肾小管上皮细胞(renal tubular epithelial cells,RTECs)损伤是肾脏疾病发生发展的重要因素。研究表明,血管紧张素II和TGF-β等因子引起RTECs慢性损伤,促进肾小管-间质纤维化,导致肾功能恶化和慢性肾脏病进展[1]。同时,线粒体功能障碍是RTECs损伤的关键环节。据报道,化疗药物等损伤因素通过降低RTECs线粒体膜电位和减少线粒体DNA复制,诱导RTECs产生线粒体自噬和凋亡[2]。钙调磷酸酶结合蛋白1(calcineurin binding protein 1, Cabin1)广泛存在于心、脑、肝等器官中,参与多种细胞功能的调节。我们的前期研究发现,Cabin1在足细胞和RTECs线粒体损伤时表达升高[3- 4],但具体的分子机制尚未阐明。本研究旨在观察敲低Cabin1对RTECs线粒体功能的影响,阐明其在RTECs线粒体损伤中的作用机制,以期为延缓肾脏纤维化提供新靶点。
1 材料与方法
1.1 RTECs培养
采用质量浓度为100 g/L的胎牛血清和100 U/mL青-链霉素的DMEM培养基,37 ℃、5% CO2培养箱,培养大鼠肾小管上皮细胞(NRK-52E,中山六院姜宗培教授惠赠)。
1.2 采用siRNA敲低Cabin1的表达
待细胞长至80%左右汇合度时,用质量浓度为0.5 g/L的trypsin-EDTA消化传代,并接种到6孔板中。待细胞汇合度达60%左右时,采用无血清的DMEM培养基培养12 h,使其生长同步化。进而将细胞分成3组: ①对照组: 采用无血清DMEM培养48 h;②阴性对照组: 采用阴性对照的siRNA转染NRK-52E细胞,6 h后更换为无血清DMEM培养基,共培养48 h;③敲低组: 采用敲低Cabin1蛋白的siRNA(正义链: 5′-CCGGAGGAGAUAAAUCUAATT-3′;反义链: 5′-UUAGAUUUAUCUCCUCCGGTT-3′)(GenePharma, China)转染NRK-52E细胞,6 h后更换为无血清DMEM培养基,共培养48 h。转染试剂采用Lipofectamine®RNAiMAX(invitrogen,USA),转染过程按说明书进行。48 h后收集细胞,Western Blot检测细胞中Cabin1蛋白表达。
1.3 WB检测Cabin1蛋白的表达
取等量蛋白行电泳、转膜、封闭,加入一抗(GAPDH, Kangchen, China ;Cabin1, Abcam, UK)4 ℃ 孵育过夜,接着用相应二抗(Anti-rabbit IgG, Cell Signaling Technology, USA)室温孵育1 h,最后加入ECL发光试剂,曝光、显影、定影,采用Image J图像分析软件进行分析。
1.4 统计学分析
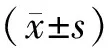
2 结果
2.1 采用siRNA敲低RTECs中Cabin1蛋白表达
在正常对照组和阴性对照组,Cabin1蛋白在RTECs中具有高的表达,采用siRNA干预RTECs后,Cabin1蛋白的表达量较对照组和阴性对照组降低50%以上,见图1。
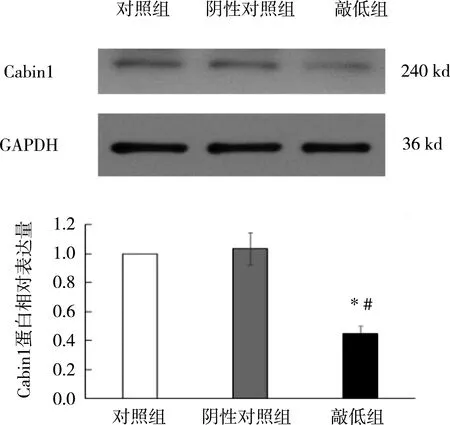
与对照组相比,*P<0.05;与阴性对照组相比,#P<0.05。图1 采用siRNA敲低RTECs中Cabin1蛋白表达
2.2 敲低RTECs中Cabin1引起线粒体形态学改变
对照组与阴性对照组细胞中,线粒体形态规则,呈圆形或椭圆形,线粒体膜完整,线粒体嵴清晰可见(白色箭头所示)。敲低组细胞中,线粒体肿胀,呈长条形或不规则形,线粒体膜、线粒体嵴结构模糊甚至消失(白色箭头所示),见图2。
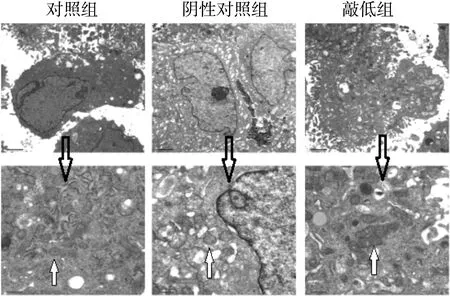
图2 敲低RTECs中Cabin1引起线粒体形态学改变
3 讨 论
RTECs中富含线粒体,后者通过氧化磷酸化为前者提供ATP,供给细胞正常代谢的能量。线粒体功能障碍引起氧化应激和炎症反应,诱导RTECs损伤,进而导致急慢性肾脏疾病的发生发展[5- 6]。本研究采用siRNA敲低RTECs中Cabin1表达,发现敲低Cabin1引起RTECs线粒体形态学改变,提示Cabin1与RTECs线粒体功能障碍密切相关。
我们的研究结果发现,敲低Cabin1引起RTECs线粒体呈长条形或不规则形,出现线粒体肿胀,线粒体膜、线粒体嵴结构模糊甚至消失。线粒体形态学的改变是线粒体功能障碍的重要指标,与肾脏细胞损伤密切相关[7]。研究者发现在阿霉素肾病大鼠模型中,模型组足细胞线粒体由圆形或椭圆形变成细长而不规则型[8]。此外,Gucer等在局灶性节段性肾小球硬化的患者肾组织中同样发现足细胞线粒体的形态学改变[9]。因此,我们推断Cabin1是维持RTECs线粒体正常功能的重要分子。
我们的前期研究发现AngII引起线粒体膜电位下调及细胞色素c蛋白的上调,同时Cabin1蛋白的表达明显升高;敲低足细胞中Cabin1引起细胞骨架蛋白F-actin的破化和细胞色素c蛋白的高表达[3]。此外,我们的体内外研究,同样证实Cabin1在RTECs线粒体损伤时表达升高[4]。我们的结果充分说明Cabin1是维持肾脏细胞线粒体功能的关键分子。那Cabin1通过什么具体的分子机制调控肾脏细胞线粒体功能呢?Cabin1通过调控p53参与多种细胞功能的调节,参与足细胞的损伤调节[10]。Peng[11]等认为p53的磷酸化引起Bax转位到线粒体膜上,继而加速细胞色素c自线粒体释放到细胞浆,导致线粒体功能障碍。Cabin1在细胞受损时,通过调控p53的表达,稳定线粒体膜上的细胞色素c,维持线粒体的正常功能。Cabin1在RTECs损伤中的具体分子机制需要我们下一步的研究来证实。
综上所述,本研究采用siRNA敲低RTECs中Cabin1表达,发现敲低Cabin1引起RTECs线粒体损伤,提示Cabin1是RTECs线粒体功能障碍的关键分子。但本研究尚未阐明Cabin1在RTECs线粒体损伤中的具体分子机制,将在下一步研究中深入探讨。
猜你喜欢
海洋通报(2021年1期)2021-07-23 01:55:14
生物学通报(2021年4期)2021-03-16 05:41:26
中国生殖健康(2019年12期)2019-01-07 01:54:30
中国继续医学教育(2015年2期)2016-01-06 01:36:16
分子影像学杂志(2015年3期)2015-12-04 03:29:03
西南军医(2015年3期)2015-04-23 07:28:34
癌变·畸变·突变(2014年1期)2014-03-01 04:39:36
振动、测试与诊断(2014年6期)2014-03-01 01:14:50
现代检验医学杂志(2014年1期)2014-02-06 01:29:31
现代检验医学杂志(2014年5期)2014-02-02 02:51:35
