
香樟天然次生林种群空间分布格局及其关联性分析
2019-06-13 00:55:18靳少非许鲁东廖晓丽范胜煌
武夷学院学报 2019年3期
靳少非,许鲁东,廖晓丽,范胜煌
(1.闽江学院 地理科学系,福建 福州 350108;2.福建省林业勘察设计院,福建 福州 350001)
种群的空间分布格局是研究种群特征,种内和种间相互作用及与环境关系的重要手段,可反映种群个体在水平空间分配和分布状况[1-2],对于揭示种群的形成和维持机制、探讨种内种间关系和种群环境关系有着重要理论意义[3-4]。种群空间分布格局的基本理论始于20世纪20年代[5],Gleason和Svedburg各自最早独立地用方差均值比率和卡方检验法进行空间分布格局研究[6-7]。我国在20世纪60年代陆续出现一些有关天然林空间分布格局的研究报道[8]。而点格局分析法作为研究种群的空间分布类型,种间的关联性和空间尺度关系的常用方法[9],在1977年由Ripley首先提出[10],20世纪90年代开始,我国学者开始运用点格局法进行乔木及草本等种群空间分布格局与空间关联性的研究[11]。近些年来,国内对林木空间分布格局的研究,多采用 RipleyK-函数,在北方地区,田慧霞[12]、赵丽琼[13]、陈列[14]等学者运用RipleyK-函数分别对山西太岳山白桦种群结构和空间分布格局、甘肃青海云杉种群空间分布格局、椴树红松林空间分布及空间关联进行研究。在南方地区,张忠华[15]、何增林[16]、徐化成[17]等人同样运用RipleyK-函数分别对桂林岩溶石山阴香种群的空间分布格局、海南热带山地夹竹桃科植物种群空间分布格局、对兴安落叶松不同年龄结构类型的种群空间分布格局进行了研究。香樟 (Cinuomomum camphoro),别名芳樟,木樟等,樟科樟属常绿乔木。是广泛分布于长江以南各省区的樟科常绿乔木,目前对香樟的研究主要集中于个体栽培、密度调控、城市绿化、叶片黄化、土壤微生物、精油提取等方面[18-21]。而对于香樟天然次生林的空间结构研究则鲜见报道,本研究以福建闽侯香樟次生天然林为研究对象,采用点格局分析方法对群落内香樟与各优势树种间、香樟种群内不同发育阶段间的空间关联进行分析,为香樟天然次生林的保护与利用提供理论依据。
1 材料与方法
1.1 研究地概况
研究区处于福建省福州市闽侯县白沙镇,位于闽江之畔、闽侯县西北部,得名于闽江大量石英砂冲积,地处东经 118°51'~119°25'、北纬 25°47'~26°37',白沙镇植被属亚热带常绿阔叶林,植被群落以香樟、马尾松、杉木、阔叶树、五节芒、芒萁骨等为主。该区域地处自然条件优越,气候温和,雨量充沛,土壤比较肥沃,有利于林木的生长,适宜建立以香樟为主的用材林基地。
1.2 调查方法
调查对象为香樟天然次生林群落,研究样地位于低山丘陵,样地海拔250~350 m,坡度26°,坡面向北,土壤为红壤,在该林分中采用相邻格子法设置两块50 m×50m样地,分别记为样地1及样地2,样地各边界按5m间距进行分割,均匀的划分出100个5m×5m的正方形调查小样方,每个单元格的编号依据在样地中对应的行列,例如:单元格1-2,即为位于第1行、第2列的单元格,行列值介于1-10之间。坐标转换:根据每个单元格在样地中的相应位置,把每株林木记录的坐标换算为以整个样地为一体的坐标。统一定义西南角为坐标原点,以此作为绘制林木空间分布图的基础。
2 研究方法
2.1 空间分布格局研究方法
采用点格局分析法进行空间分布格局研究,点格局分析法是以个体在空间的坐标为基础数据,每个个体可视为二维平面上的一个点,对点之间进行空间格局分析的一种方法,点格局分析法考虑了点的空间位置,是真正意义上的空间分析。
2.2 数据处理和分析
采用单变量O-ring函数并以完全空间随机模型为零模型分析香樟群落及各优势种群个体的空间分布格局[22],采用双变量O-ring函数分析香樟种群不同发育阶段的空间关联性及与其它主要树种的空间关联性[23]。空间格局分析过程使用生态学软件Programita2014完成,空间尺度 0~25 m,步长 1 m,将植物种群个体视为二维平面上一个点,形成空间分布点图。为了提高种群空间分布的分析精度,经199次Monte Carlo simulation循环,得到置信区间99%的包迹线以检验点格局分析的显著性。对于单变量O-ring函数计算,实际数据计算得到的种群在不同尺度下O(r)值若在下包迹线以下,则为均匀分布;若在包迹线之间,则为随机分布;若在上包迹线以上,则为聚集分布。对于双变量O-ring函数计算,实际函数O(r)值若高于置信区间上限,则在r尺度处正关联;若O(r)值低于置信区间下限,则在r尺度处负关联;若O(r)值在置信区间内,则表明种内和种间二者相互独立即无显著关联。
单变量O-ring计算公式:

式中:n1是对象种1中的个体数量;表示在种群1中以第i个点为圆心,以r为半径,以w为宽的圆环;Po int s2是对象种 2 在区域内的个体数;Area是样地面积;(xi,yi)是对象种 1 第 i个点的坐标;S(x,y)为变量,(x,y)视是否在研究区域而定,在则值取 1,否则值取 0;P2(x,y)是指每个小格中包含对象种 2 的点数;Ir(xi,yi,x,y)也是一个变量,随对象种1中第i个点为中心、以r为半径的圆而变;z2表示小格的面积。
3 结果与分析
3.1 香樟次生天然林群落的空间分布格局研究
3.1.1 香樟群落整体空间分布格局
应用上述点格局分析法分别计算样地1与样地2群落空间分布结果如下:
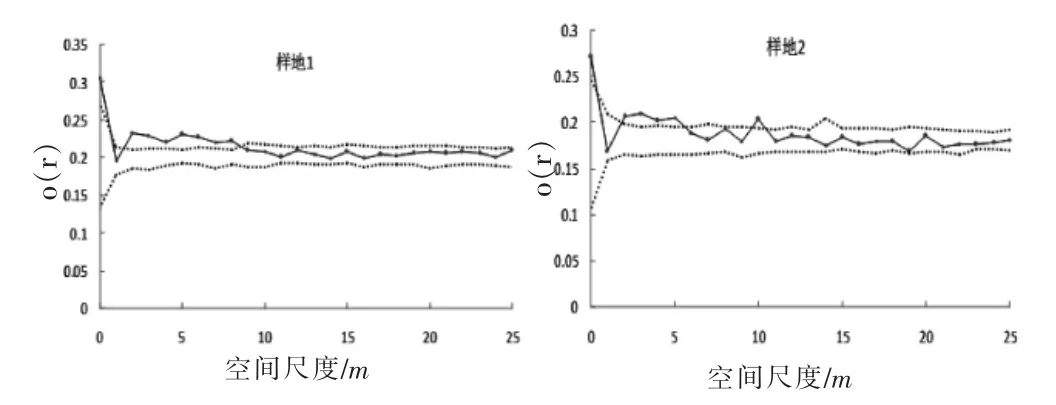
图1 群落空间分布格局Figure 1 The spatial distribution pattern of camphor community
由图1与图2可见,样地1在3~9 m尺度下呈聚集分布,其他尺度随机分布;样地2在3~6 m和10m下呈聚集分布,其他尺度随机分布。两个样地均表现为在小尺度下呈聚集分布,随着尺度增大,在大尺度下逐渐趋向于随机分布。两块样地整体空间分布格局变化趋势相似,这与香樟不同发育阶段数量特征统计分析结果相一致。

图2 样地内林木空间分布点图Figure 2 Spatial distribution of forest trees in plots
3.1.2 香樟次生天然林群落优势树种空间分布格局
进一步应用点格局分析法分析计算样地1与样地2中优势树种空间分布结果如下
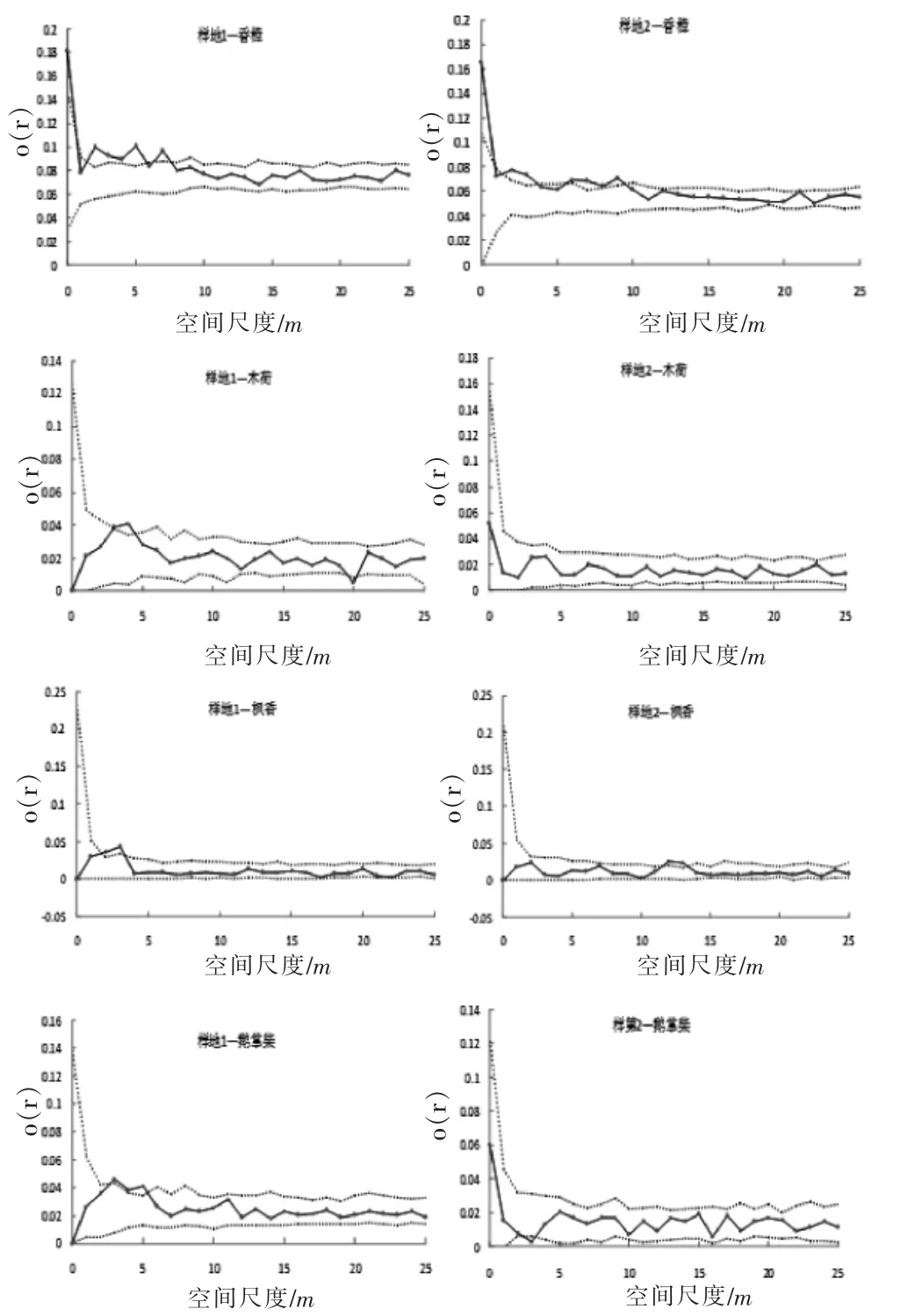
图3 各主要优势树种空间分布格局Figure 3 Spatial distribution pattern of themain dominant species
由图3可见,样地1在2~5 m和7 m呈聚集分布,样地2在2~3 m和6~9m呈聚集分布,大尺度上局部有些不同,但不影响整体大尺度上的随机分布的格局,其空间分布格局与群落整体相似,体现了香樟在群落中的优势地位;杨桐在样地1中除13~15m尺度下呈聚集分布,其他尺度呈随机分布,在样地2中的除10m和15m尺度下呈均匀分布,其他尺度均呈随机分布,杨桐多尺度下的随机分布及小部分均匀分布可能与动物取食有关,植物种子是啮齿动物的取食对象,动物在取食过程中的搬运和储藏均可能导致杨桐在大尺度下的随机分布;青冈种群在样地1中5、7~9、14m 尺度下呈聚集分布,在样地 2 中 5、7、10~11、15、20m尺度下呈聚集分布,相较于其他树种,其聚集尺度范围更大,这可能与其果实种子个体较大,掉落较易聚于母株边形成聚集分布;木荷种群在样地2中呈随机分布,在样地1中3~4 m尺度下呈明显聚集分布;枫香在样地1中3~4 m尺度内呈聚集分布,其余呈随机分布,样地2内12~13 m尺度下呈聚集,其余尺度内随机分布;鹅掌柴在样地1内3~5m尺度内呈聚集分布,其余尺度为随机分布,在样地2内2~3 m尺度下为均匀分布,其余尺度呈随机分布。大部分优势树种统一在大尺度下表现随机分布,在小尺度下呈现局部聚集,这与小尺度上的种内竞争、种子扩散机制及大尺度上的生境有关。
3.2 香樟与其他优势树种的空间关联性分析
通过香樟与其他优势树种间的空间关联性分析以了解天然香樟次生林对于生境与空间的资源的适应性与依赖性,结果如下:
由表1可见,样地1中,香樟与木荷在 3~4、10~11、17、19~20 m尺度上存在的正关联,大部分尺度下表现为相互独立;香樟与枫香在5~6m和16m尺度上表现为正关联;香樟与鹅掌柴在22m尺度上呈正关联,其它尺度为无关联;香樟与青冈在5~7、15~17m尺度上呈正关联,在其它尺度呈相互独立;香樟与杨桐在2、8~9 m尺度上呈正关联,在6 m尺度呈负关联,其他尺度呈相互独立特点。通过表2可见,样地2中,香樟与木荷在 2、5~6、12、16~17m 尺度上呈正关联;与枫香在3~4、11~12 m及19 m尺度上呈正关联;与鹅掌柴在5~6 m尺度上呈正关联;与青冈在4、6、15~16、18~19m 尺度上呈正关联;与杨桐在 17~18 m尺度上呈正关联,在3~4 m小尺度内呈负关联;香樟与各主要优势树种其他大部分尺度上呈相互独立的特征。综合两个样地来看,香樟与木荷、枫香、青冈具有更多尺度上的正关联,与杨桐存在的一定尺度的负关联,但在大部分尺度上偏向于相互独立的无显著关联性。

表1 样地1主要树种和香樟的种间关联性Table 1 Inter specific association of major tree species and cinna momum camphora in plot1
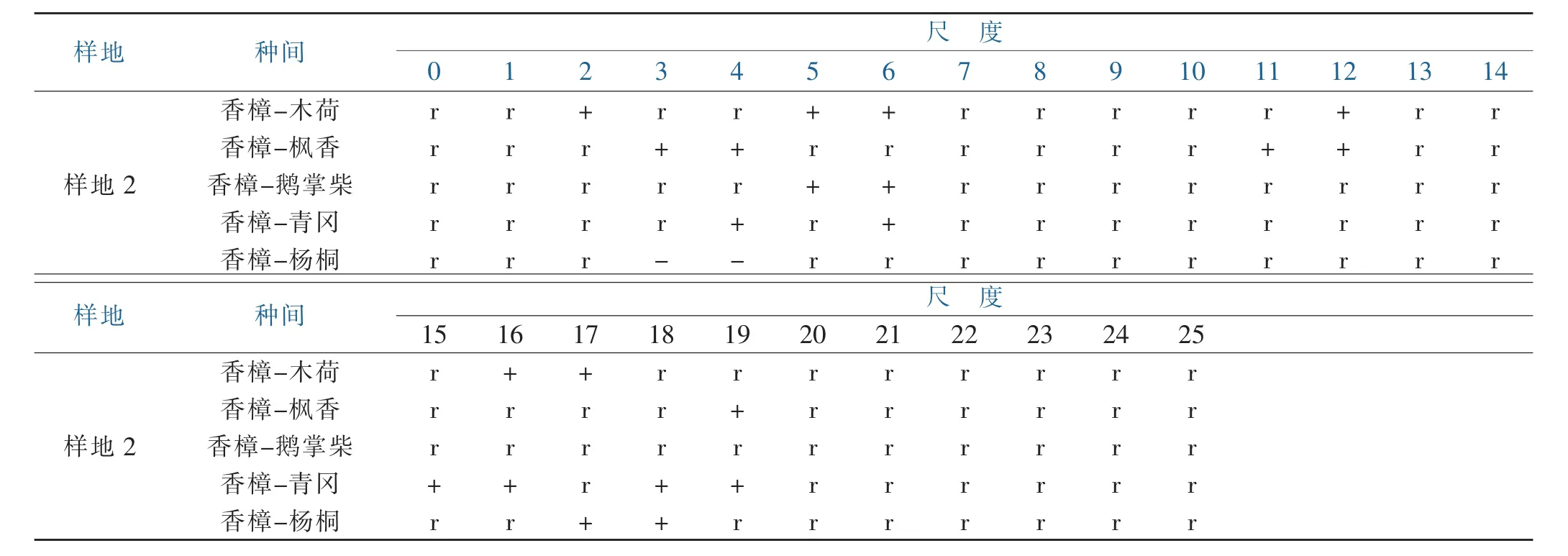
表2 样地2主要树种和香樟的种间关联性Table 2 Interspecific association ofmajor tree species and cinnamomum camphora in plot2
3.3 香樟不同发育阶段间的空间关联性分析
依据胸径大小将香樟划分为幼树DBH<7.5 cm,中树7.5 cm ≤DBH<22.5 cm,大树DBH≥22.5 cm,三个发育阶段。使用完全随机零假设模型,假设高龄级对低龄级个体生长有影响,低龄级对高龄级个体生长没影响。
由表3分析香樟不同发育阶段空间关联性变化。在样地1中,香樟幼树和中树在3、6~7、9~10 m尺度上呈显著正关联,其他尺度上呈无关联性;幼树和大树在 1~3、6、11、16~17 m 尺度上呈正关联,其他尺度上呈无关联;中树和大树在2m尺度处呈负关联。在样地 2 上,幼树和中树在 2~4、6、8~9m 上呈显著正关联,其他尺度呈无关联;幼树和大树在 2~3、5、10、17m尺度呈显著正关联,其他尺度呈无显著关联;中树和大树在3~4m尺度上呈负关联性,其他尺度呈无关联性。在样地 2 上,香樟幼树和中树在 2~4、6、8~9m 尺度上呈显著正关联,其他尺度呈无关联;幼树和大树在4~5、10、17m尺度上呈显著正关联,在其他尺度呈无关联性;中树和大树在3~4 m尺度上呈显著负关联,在其他尺度上呈无关联性。综合两块样地可以看出,香樟不同发育阶段间的空间关联性随尺度的变化有所不同,幼树和中树、大树在小尺度上均呈一定显著正关联,随尺度增大关联性逐渐减弱,表明幼树在小尺度上对中树、大树有着明显依赖性,这符合幼树寻求中树和大树保护的生存特性;同时,两块样地中树和大树间均存在个别尺度上的负关联性,表明样地内一定尺度下存在着激烈竞争。
4 讨论和结论
香樟次生天然林群落空间分布格局总体上表现为小尺度上局部聚集分布,随尺度的增加由聚集分布到随机分布的特征,聚集程度随空间尺度增大而降低,这与大多数学者研究成果一致[1]。群落优势树种空间分布格局从整体上看多尺度上表现为随机分布尤其是大尺度上,小尺度有聚集分布的现象如青冈、鹅掌柴、木荷,这与小尺度上的种内竞争、种子扩散限制及大尺度上的生境有关;香樟优势树种种群的空间分布格局与群落整体空间分布格局相似,在群落中表现出绝对的优势;青冈种群小尺度聚集现象明显,与其种子扩散有关,种粒大且重,不易受风力影响,分布与母株周边,聚集生长,这与前人研究成果一致[24];杨桐树种的随机与均匀分布则与啮齿动物取食密不可分。
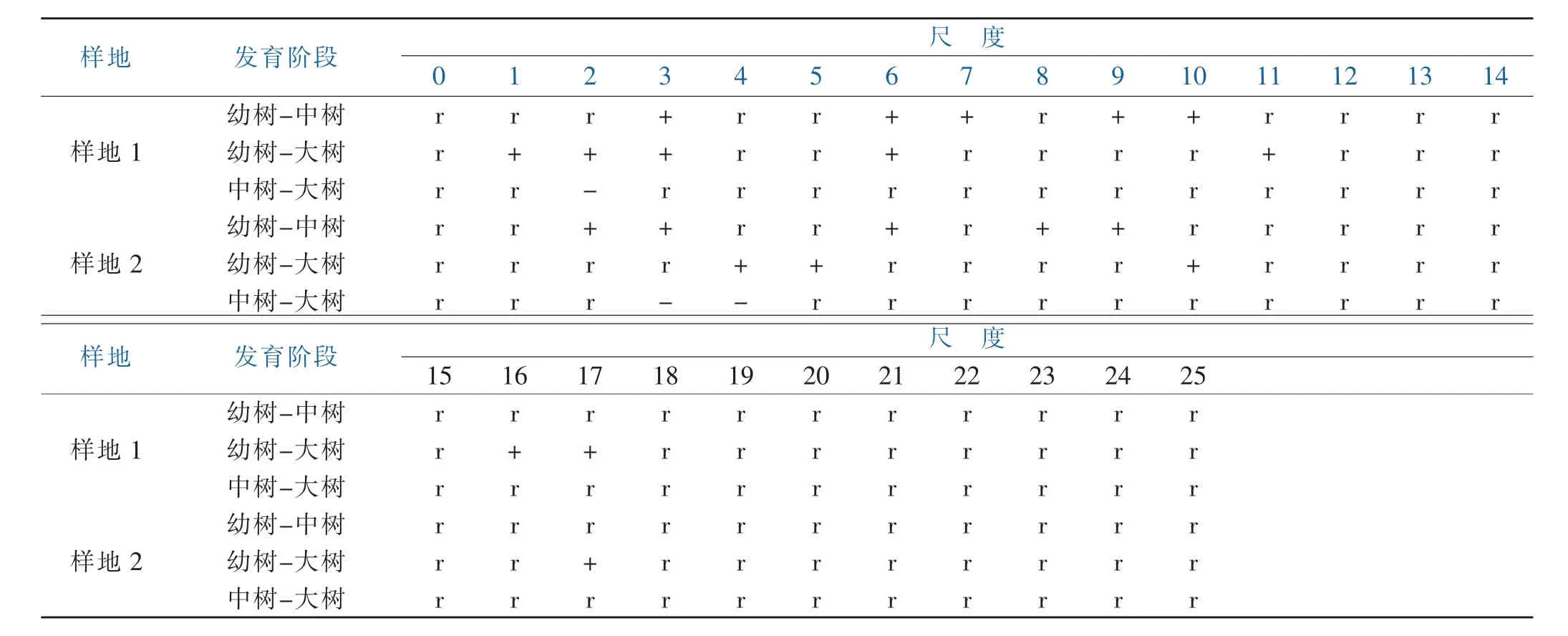
表3 不同发育阶段香樟空间关联性Table 3 Spatial correlation of cinnamomum camphora at different developmental stages
树种间关联性是对环境和空间资源利用相适应的结果,对生境的依赖性往往决定种间关联性[25]。香樟与其他优势树种之间大部分尺度上表现为相互独立的无关联性,局部有正关联,这主要与优势树种生态习性各不相同有关,一定程度上说明香樟与这些优势树种可以共存,空间竞争较小,形成一定空间上的依赖。香樟与杨桐树种在样地1和样地2中小尺度上均存在负关联性,表明杨桐与香樟存在局部尺度上的激烈竞争,可能是在香樟所形成的主林层中存在一定数量杨桐,与其竞争空间资源较大所造成的。香樟与不同树种间空间关联性的不同,充分说明了不同树种间空间关联的复杂性,在今后更深入的研究中需做进一步的探讨。
对香樟幼树、中树、大树三个发育阶段间的空间关联性进行分析,两块样地中香樟三个发育阶段间绝大部分尺度上呈无显著关联性,幼树和中树、大树间小部分呈显著正关联,结果表明,香樟幼苗为获得更好的生长空间资源,出现在中树和大树斑块间隙,尽量减少中树和大树对其自身生长的影响,但在局部形成对中树和大树的依赖,表现出幼苗寻求中树和大树保护的生物特性。香樟中树和大树在样地1中2m尺度上和样地2中3~4m尺上呈现负关联性,表明在小尺度上香樟中树和大树间存在一定空间资源上的种内竞争,这一定程度上影响了中树生长为大树的速度。
一般而言,随着群落演替的发展,种间的正关联性程度逐渐加大,当群落演替到顶级群落时,种间关系表现为明显正关联[26]。本次研究的香樟次生天然林群落中优势种在大多数尺度上表现为相互独立的无关联性,说明香樟次生天然林群落的结构未达到很稳定的状态,处于相对稳定状态,群落演替发展处于演替初期群落地过渡阶段,在接下来的群落演替发展中,种间竞争竞争将越发激烈,物种更替频繁,种群更替速率加快,群落整体更新速率将处于较高水平。
猜你喜欢
现代园艺(2021年23期)2021-12-01 07:47:44
西安交通大学学报(社会科学版)(2021年2期)2021-04-06 03:46:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
北方音乐(2019年6期)2019-07-16 07:50:53
现代园艺(2017年23期)2018-01-18 06:58:00
中成药(2017年3期)2017-05-17 06:09:05
中国环境监察(2016年12期)2016-10-24 05:29:18
中国卫生标准管理(2015年6期)2016-01-14 05:17:08
小小说月刊·下半月(2015年12期)2015-05-14 15:23:21
