
簕杜鹃花芽分化解剖与形态学研究
2019-06-05 05:33:38谢利娟王定跃
西南林业大学学报 2019年3期
聂 艳 谢利娟 王定跃
( 1. 深圳人民公园管理处,广东 深圳 518001;2. 深圳职业技术学院应用化学与生物技术学院,广东 深圳 518055;3. 深圳梧桐山风景区管理处,广东 深圳 518004)
簕杜鹃为紫茉莉科(Nyctaginaceae)叶子花属(Bougainvillea),属于攀援状藤本灌木。产自于南美洲的巴西,中国引种栽培的有簕杜鹃(Bougainvillea spectabilis)和毛簕杜鹃(Bougainvillea glabra)2种,为典型的热带植物。簕杜鹃别名三角梅,为短日照植物,长日照条件下很难完成花芽分化而开花[1],喜强光、空气流通的环境。生长适应性强,管理方便,喜水,但忌积水,对土壤要求不严,但在肥沃、排水良好、疏松的弱酸性砂质壤土条件下生长旺盛,幼株适宜的土壤pH为6.0~6.5[2]。由于其易于繁殖栽培,生长适应能力强,花型独特且花期长,从10月份开始一直到翌年6月,观赏价值极高,自引种以来,簕杜鹃就成为我国广东省珠海市、深圳市以及福建省厦门市等很多城市的市花,广泛应用于盆栽制作盆景,地栽作绿篱、围墙等装饰,其攀援性较好的种类广泛用于花架、花廊的装饰以及高架桥的垂直绿化;同时叶子花中含有花青苷和黄酮苷,具有抗肿瘤、抗氧化等多种功效[3]。
花芽分化过程复杂,受温度、光周期、水分等外界环境条件以及内部代谢平衡等多种因素的影响。花芽分化的起始标志着植株营养生长结束,生殖生长开始,是植株花期早晚、花量的多少以及观赏效果的决定因素[4-5]。在栽培生产中,通过探究植株花芽分化进程,了解植株花芽分化的特性,进而采取相应的管理措施完成花期调控,为植物的繁殖栽培以及进一步的杂交育种提供理论依据。汪晓谦等[6]通过对杨凌地区郁金香(Tulipa gesneriana)花芽分化特性的研究发现,其在杨凌地区花芽分化开始较早,因此夏季高温储藏期间要安装通风扇等降温通风设备,保证其顺利进行花芽分化;杨柳燕等[7]根据彩色马蹄莲(Zantedeschia hybrids)的花芽分化特性,制定高效的栽培措施从而达到提前花期增加花量的目的;黄凤兰等[8]通过对芍药(Paeonia lactiflora)花芽分化的显微结构研究,为其栽培学及遗传育种方面的研究提供一定参考。簕杜鹃作为深圳市的市花,由于受不定期的低温、台风等天气状况影响,使得植株出现开花不整齐,不能在特定季节开花等现象。然而国内外有关簕杜鹃的研究主要集中在繁殖栽培方面[1,9-10],Lopez等[11]仅观察了簕杜鹃花的结构以及传粉过程,有关花芽分化特性等的研究尚未见报道。为了掌握簕杜鹃的花芽分化规律,明确簕杜鹃花芽分化的时间与特点,以‘同安红’簕杜鹃为研究材料,探索其花芽分化进程,掌握簕杜鹃不同分化阶段的形态以及解剖结构特征,研究簕杜鹃的开花机制,从而为栽培生产中实现提高开花质量、增加花量以及更合理的花期调控提供参考。
1 材料与方法
1.1 研究材料
选用‘同安红’簕杜鹃(B.spectabilis‘Crimelake’)为1年生扦插苗为研究对象,株高约为52.2 cm,2016年7月25日以45 cm×60 cm的行间距盆栽摆放在深圳市公园管理中心大沙河基地,其形态特征观察见图1。
1.2 研究方法
1.2.1 取样方法
于2016年8月上旬至9月上中旬进行试验观察采样,每隔5 d连续采样。每次随机采取10盆植株上同一发育时期且大小相似的花芽,立即放入冰盒内,带回实验室对花芽的内部解剖结构以及其外部形态进行观察研究。
1.2.2 花芽超微结构观察
对一部分采回的花芽采用常规石蜡切片法[12]制作成永久性切片,室温下彻底干燥后使用电子显微镜(Nikon H600L)观察花芽内部解剖结构特征并拍照,获得簕杜鹃各个花芽分化阶段对应的内部解剖结构图。

图 1 ‘同安红’簕杜鹃花部形态特征Fig. 1 Morphological characteristics of B. spectabilis ‘Crimelake’ flower parts
1.2.3 花芽外观形态观察
从不同阶段取回的花芽中选取一部分(5个)进行观察拍照,获得相应阶段的花芽形态特征图,同时测量不同阶段花芽的长度以及直径,记录并分析每个阶段花芽的外观形态变化特征。
2 结果与分析
2.1 花芽分化过程中芽的解剖结构特征
通过对花芽不同分化阶段的显微结构观察,发现‘同安红’簕杜鹃从8月初期进入生理分化期,9月初基本完成花芽的形态分化,大约经过30 d。整个分化过程可分为未分化期、花序原基形成期、苞片原基形成期、小花原基形成期、萼片原基形成期、花瓣原基形成期、雄蕊及雌蕊原基形成期等8个时期,不同分化阶段相对应的内部解剖图以及外部形态变化见图2~3。
2.1.1 未分化期
8月初期,簕杜鹃尚未进入花芽分化阶段,这一时期主要进行植株的营养生长,花芽的分化发育受水分、温度、光周期等外界环境条件以及内部营养状况的影响,当植株自身的营养条件达到一定的标准后,随即向生殖生长过渡。通过对花芽石蜡切片的显微结构观察,发现该阶段芽体生长点发生变化,顶端呈圆锥状,组织形态狭小而尖,同时生长点两侧的叶原基也呈现出不断向内分化的趋势,且外层的组织细胞紧密排列,从而保护其内侧的生长锥(图3a)。该阶段芽体瘦小,芽鳞片包裹紧实,芽长仅为1.06 mm,芽体直径为1.75 mm(图2a)。
2.1.2 花序原基形成期
8月上旬为花序原基分化期,通过石蜡切片显微结构的观察,发现芽体分化初期,生长锥顶端逐渐变宽变平,略微向上突起,由未分化期的圆锥状变为圆弧形;随着芽体的继续分化,生长点顶端逐渐下凹,两侧形成小的凸起,之后逐渐发育为苞片原基(图3b)。此时芽长约为1.42 mm,芽直径为2.04 mm,呈现出一定的伸长和变宽,但芽长变化不显著(P>0.05),很难通过外观判断其分化进程(图2b)。
2.1.3 苞片原基形成期
8月上中旬,花芽进一步膨大变宽,内部细胞持续伸长,生长点两侧形成的小突起继续伸长、膨大,苞片原基形成。在整个花序形成过程中,苞片原基不断分化发育,并向上伸长弯曲,分化状态不规则;随着细胞不断地发育分裂,苞片原基包裹着的花序原基逐渐膨大形成凸起(图3c)。此时芽长约为1.58 mm,芽直径为2.41 mm,芽体包裹的幼叶逐渐伸长展开(图 2c)。
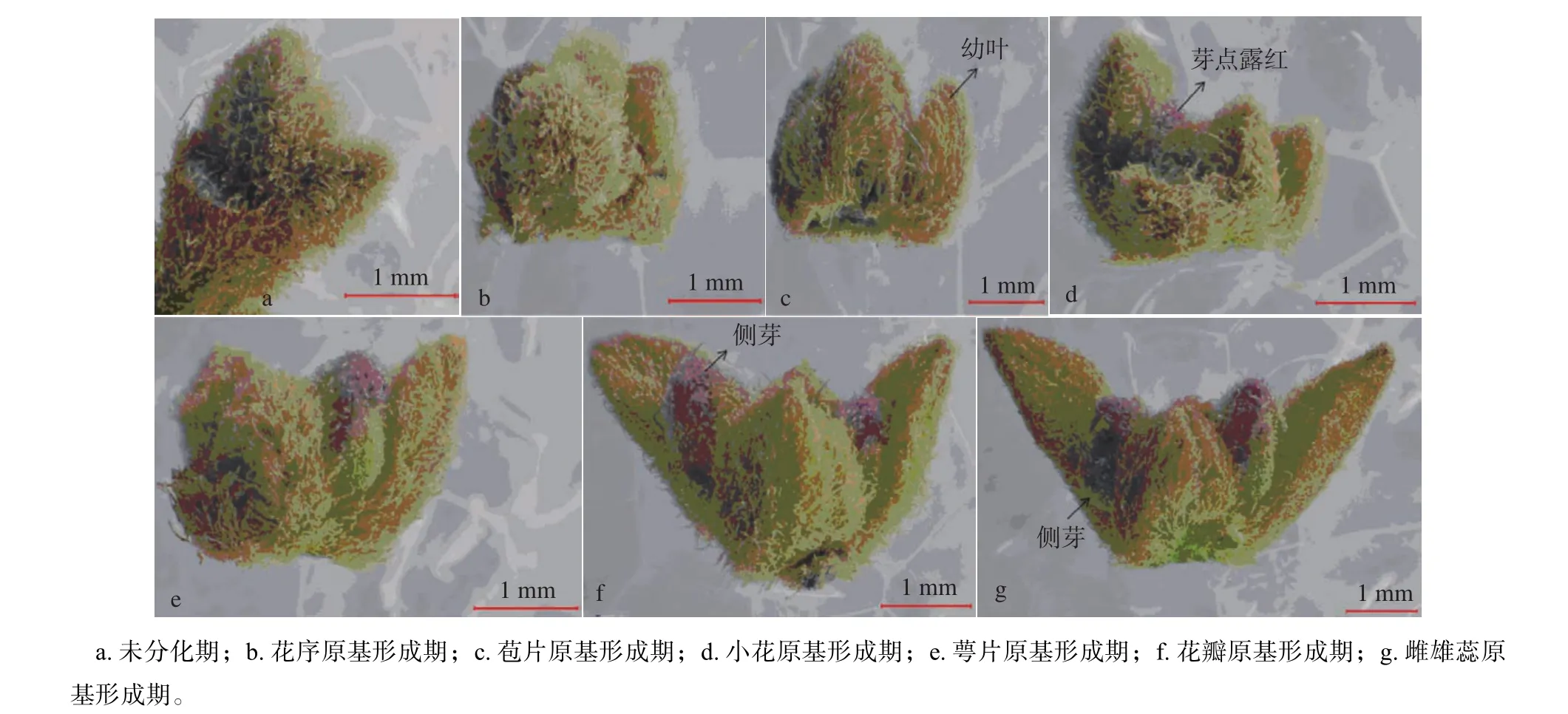
图 2 不同分化阶段花芽形态特征Fig. 2 Flower bud morphological characteristics of differentiation at different stages
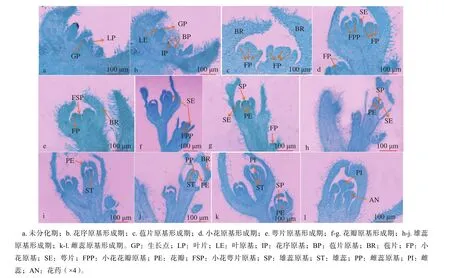
图 3 花芽分化过程Fig. 3 Flower bud differentiation process
2.1.4 小花原基形成期
8月中旬,随着生长锥的持续膨大变宽,其两侧不断形成小凸起,后期分化发育为侧花芽,因此该阶段芽体直径表现出明显的膨大变宽,通过超景深显微系统拍照测量得出芽长为1.84 mm,芽体直径为2.64 mm(图2d)。通过显微结构观察发现,花序原基不断的分化发育,形成椭圆形的小突起,即为小花原基;小花原基分化发育的同时,在其两侧分生出明显的圆柱形平滑的小突起,侧花原基形成。至此,簕杜鹃复伞形花序的结构特征基本形成(图3d)。
2.1.5 萼片原基形成期
8月中下旬左右,簕杜鹃开始进入萼片原基形成期,此时通过石蜡切片显微结构观察发现,随着芽体的持续分化发育,生长点顶端慢慢变平,并向下凹陷,逐渐在小花原基周围形成小的凸起,萼片原基形成,最终发育为花萼(图3e)。该阶段花芽继续伸长膨大,包裹的幼叶慢慢展开,芽长变化显著(P<0.05),芽长为2.13 mm,芽直径为2.95 mm(图2e)。
2.1.6 花瓣原基形成期
8月下旬,通过石蜡切片显微结构观察发现,花芽迅速分化发育,花萼原基分裂速度较快,持续伸长变宽,同时向内弯曲,在其内侧形成新的凸起,花瓣原基形成(图3f-g)。此时花瓣原基生长速度快,幼叶全部展开,芽长为2.58 mm,芽体直径为4.06 mm,花芽体积相当于未分化期芽体体积的2倍(图2f)。
2.1.7 雄蕊原基形成期
8月下旬到9月初期,花芽继续膨大发育,萼片和花瓣逐渐伸长,生长锥继续变宽,花瓣原基的外侧分化出小的凸起。雄蕊原基形成(图3hj),此时芽体宽度分化速度明显大于长度;雄蕊原基形成初期,顶端浑圆光滑,随着进一步的分化发育,顶部开始呈纵向分裂的趋势,逐渐发育形成花丝、花药等内部组织结构。
2.1.8 雌蕊原基形成期
9月初期,雄蕊原基继续发育,其内侧逐渐形成小的凸起,即雌蕊原基形成(图3k-l)。此时芽体分化速度依然很快,表现出明显的膨大与变宽,芽长为3.37 mm,芽直径为4.51 mm,芽体体积相当于营养生长阶段的3倍,具有显著性差异(P<0.05)(图2g)。至此,基本完成整个花器官的分化。分化过程中,不同阶段相互之间出现一些交叉的现象,最终形成完整的复伞形花序,这一分化特征同天竺葵(Pelargonium hortorum)、紫杏(Armeniaca.dasycarpa)等的分化特性相似[13-14]。
2.2 簕杜鹃花器官发育过程
雄蕊及雌蕊原基形成之后,内部细胞组织持续分化发育,且雄蕊原基分化速度较快,持续向上伸长,形成一条条柱状群,即为雄蕊群发育形成(图4a)。随着芽体的进一步分化发育,雄蕊原基继续分化,条形的柱状群持续伸长,下端形成细长的花丝,顶部膨大形成花药,雄蕊群在雌蕊的周围均匀分布,并在雌蕊基部联合(图4b,c)。在雄蕊持续伸长的同时,雌蕊也在不断的向上伸长,基部逐渐膨大发育为子房,子房发育内部形成胚珠原基(图4d);雄蕊持续分化发育,形成成熟的花药,通过花被管的横切面可以发现,发育成熟的花芽有8枚花药,内部含有较多花粉粒,在中心的子房周围均匀分布(图4h),此时整个雄蕊群为棒槌状(图4e),在雄蕊原基不断发育的同时,雌蕊原基也持续向上伸长,发育为细长的花柱(图4f,g)。
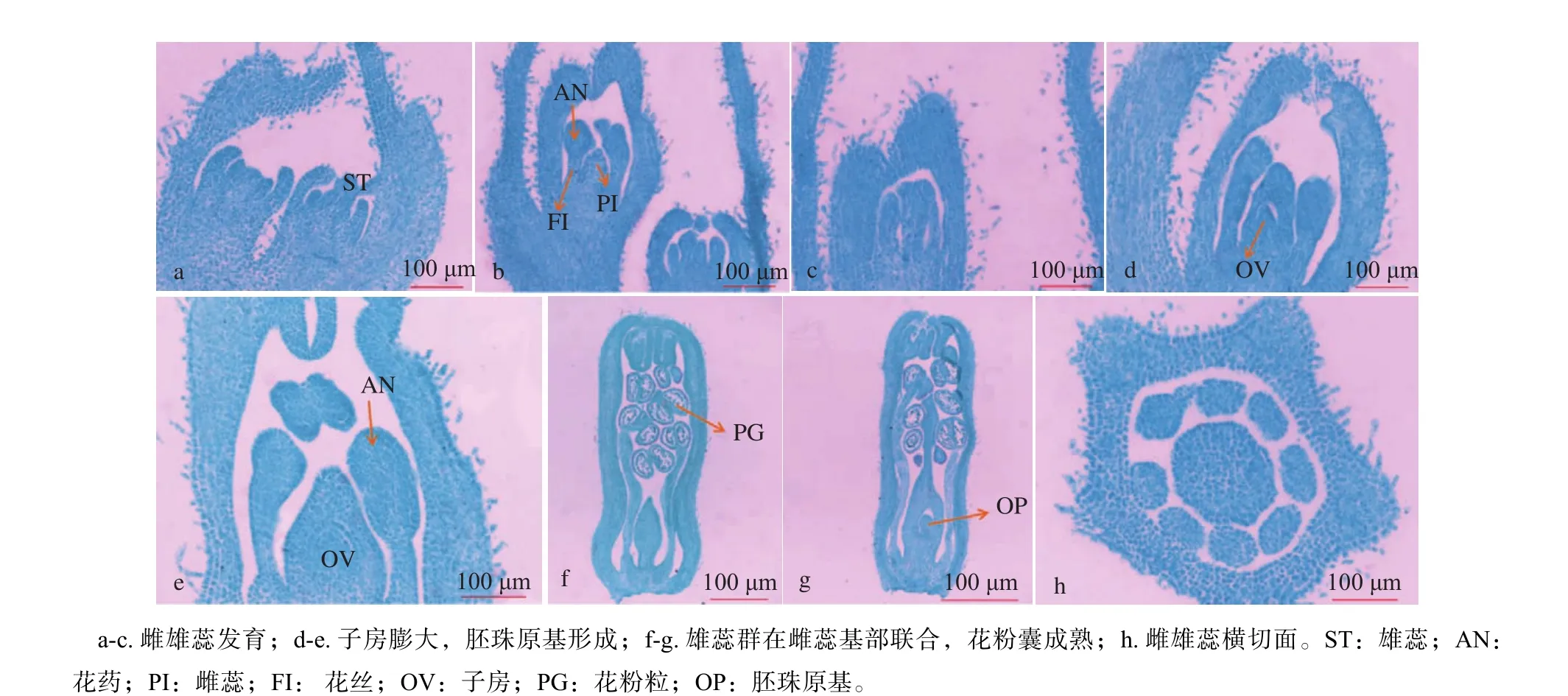
图 4 花器官发育过程Fig. 4 The development of floral organs
2.3 花芽分化过程中外部形态变化
在营养物质充足且外界环境条件适合的情况下,簕杜鹃芽体内部花器官不断的分化发育,同时外部形态也不断的生长变化,主要表现为芽体的膨大与变宽,整体过程均表现为平均芽宽大于平均芽长,呈椭圆形(表1、图2)。从表1中可知,花芽后期(花瓣原基形成期)分化速度比前期加快,芽直径明显加宽(P<0.05),基本可以通过外部形态的变化判断芽体内部的分化过程。通过芽体的解剖结构可以看出,刚进入分化期或者未分化期的花芽芽体非常瘦小,外围包裹紧密的芽鳞片(图3),芽体平均长度约为1 mm(表1),因此,很难通过肉眼观察外部形态去判断内部的分化特征,必须通过观察内部解剖结构特征确定具体的分化进程。随着营养物质的积累,芽体的进一步分化发育,芽长和芽直径不断的伸长和膨大;在顶端生长点不断发育的同时,在其两侧不断分生出新的花原基,即侧花原基,继而形成完整的复伞形花序。

表 1 簕杜鹃不同分化阶段花芽外部形态变化Table 1 Morphological changes of flower buds of B. spectabilis differentiation at the different stages
2.4 不同花芽分化期外部形态特征比较
在植物的花芽分化进程中,芽体内部解剖结构特征的变化在一定程度上和外部形态相对应,同时外部形态上的膨大与变宽也反应出内部细胞组织的分化发育。因此在实际研究过程中,可以将花芽外部形态特征的变化和内部解剖结构特征相结合,利用二者之间的相关性,建立一定的对应关系,进而更明确地判断簕杜鹃具体的花芽分化进程,从而对簕杜鹃花芽分化进程建立更清晰的认知,以便于今后开展花期调控满足生产栽培需求,具体对应关系见表2。

表 2 花芽分化进程及其外观形态Table 2 The differentiation process and external morphology
3 结论与讨论
植物从营养生长向生殖生长过渡的一个重要标志即为花芽分化,生产栽培中,花芽分化起始时间、分化的数量以及花芽分化质量的高低等因素都直接影响植株开花的效果,决定其观赏价值和生产应用。因此,很多研究学者通过探索植株的花芽分化进程指导花期调控[15-17],通常情况下,专家学者们将植物的花芽分化进程分为未分化期、萼片原基形成期、花序原基形成期、花瓣原基形成期及雌雄蕊原基形成期,不同阶段的划分通常以其对应的内部原基开始分化为依据[18-19]。植物雌雄蕊原基形成之后,内部细胞组织继续分化发育,有的研究者认为,雄蕊及雌蕊原基的继续分化发育,形成后期的花药、花丝以及成熟的花粉粒,同时子房膨大形成成熟的胚珠等过程,都是在雌蕊及雄蕊原基分化发育的基础上继续发育产生的,而且会对植株的受精结实以及果实的成熟造成很大的影响,因此该过程应该划分为花芽分化的进程范畴内[20];但也有人认为,雌雄蕊原基的进一步分化发育,形成成熟的内部细胞组织,是性细胞逐渐成熟的过程,因此应该属于性细胞发育成熟的阶段。
有关木本植物花芽分化特性的研究有很多[21-24]。探究簕杜鹃花芽分化的具体进程,以及每个分化阶段的特证,有助于更明确的认识簕杜鹃的生物学特性,为开展花期调控、指导生产以及今后的遗传育种等提供一定的参考。通过对簕杜鹃花芽解剖结构以及外部形态的观察研究,发现簕杜鹃属于腋生纯花芽,只要积累到足够的养分,且光照、水分等外界条件满足的情况下,就能进行花芽分化,完成花的发育,形成完整的复伞形花序。簕杜鹃整个花芽分化进程大致可以分为未分化期、花序原基形成期、苞片原基形成期、小花原基形成期、萼片原基形成期、花瓣原基形成期、雌雄蕊原基形成期8个时期,雌雄蕊原基产生凸起后,花药以及子房分化发育,形成成熟的花粉囊和胚珠,至此雌蕊和雄蕊完全发育成熟。从八月初期开始分化,到9月初期基本结束,持续30 d左右,这与罗汉果(Siraitia Grovenorii)[25]、龙葵(Solanum nigrum)[26]、牡丹(Paeonia suffruticosa)、芍药[27]的研究基本相同,莫长明等[25]、祁宏英等[26]没有划分苞片原基分化期、小花原基分化期,其花芽分化初期特征和簕杜鹃花序原基分化期基本一致。研究过程中,通常以花芽生长锥顶端的凸起或凹陷以及外部形态的伸长和膨大等作为花芽开始分化起始的特征[19],但由于簕杜鹃花芽分化起始时外部形态变化不显著,从开始进入形态分化期一直到小花原基形成期过程中,芽直径变化不显著(P>0.05),仅由2.04 mm膨大到2.64 mm,因此必须通过石蜡切片的显微结构观察,研究其内部解剖结构的变化,确定其花芽分化进程,这一分化特点与荷兰水仙(Narcissustazetta)[28]、彩色马蹄莲[7]相似,这可能是由于簕杜鹃花芽分化前期主要进行内部细胞组织的发育,生产中可通过对该阶段花芽生理生化特性的测定探究分化前期花芽的理化性质,从而采取针对性的措施达到推迟或提前花期的目的。此外,通过对簕杜鹃花芽石蜡切片的显微结构观察发现,在簕杜鹃花芽分化进程中,不同阶段之间存在相互交叉的现象,在生长锥顶端生长发育的过程中,其两侧也不断地形成新的小花原基,进而形成新的小花;小花原基继续生长发育,随即在其两侧分化出新的侧花原基,最终发育形成一个完整的复伞形花序,通常3枚叶状苞片组成1朵花,3朵花组成一个完整的伞形花序。生产栽培中,可根据簕杜鹃花序自身的分化发育特征,在特定时期采用一定的促进或抑制措施达到增加或减少花量的目的。
通过对簕杜鹃的花芽分化进程以及相对应的外部形态特征的观察,初步了解了簕杜鹃的花芽分化规律,将簕杜鹃的花芽分化过程划分为未分化期、花序原基形成期、苞片原基形成期、小花原基形成期、萼片原基形成期、花瓣原基形成期、雄蕊及雌蕊原基形成期等8个时期,这一结论的得出有助于人们进一步了解其花芽分化特性,认识簕杜鹃花器官形态建成的规律,从而为生产栽培中更有效合理的开展花期调控、满足生产应用提供参考。本次研究过程中,笔者尚未对簕杜鹃花芽分化时间做精确地划分,对花芽分化不同阶段花器官内部理化性质的变化未做深入研究,关于簕杜鹃各个花芽分化阶段理化性质的探索,也仅见少量粗略的研究[29],因此,下阶段希望对簕杜鹃不同花芽分化阶段生理物质的变化特性做进一步深入研究。
猜你喜欢
烟台果树(2022年2期)2022-11-26 13:01:32
果树实用技术与信息(2022年4期)2022-11-21 04:43:16
果农之友(2022年8期)2022-08-26 08:25:44
中学课程辅导·教师教育(上、下)(2018年15期)2018-10-10 07:56:54
山西农业科学(2018年9期)2018-09-12 02:25:34
生物学教学(2017年6期)2017-02-18 10:14:33
红蜻蜓·低年级(2016年9期)2016-05-14 10:50:49
中国马铃薯(2015年3期)2015-12-19 08:03:54
今传媒(2015年12期)2015-05-30 05:12:32
华南师范大学学报(自然科学版)(2014年6期)2014-08-28 00:43:38
