
DrwH类信号肽序列对其抗氧化功能的影响
2019-06-04 07:04:56郭磊周韩佳慧唐殷李江黄程代其林王劲平淑珍江世杰
生物技术通报 2019年5期
郭磊周 韩佳慧 唐殷 李江, 黄程 代其林 王劲 平淑珍江世杰
(1. 西南科技大学生命科学与工程学院,绵阳 621010;2. 中国农业科学院生物技术研究所,北京 100081)
异常球菌属细菌是地球上迄今为止发现的最具辐射抗性的微生物,其中耐辐射异常球菌(Deinococcus radiodurans)R1是 1956年首次被鉴定的异常球菌属菌株,该菌对多种非生物胁迫具有超强的抵抗能力,包括γ-辐射、UV辐射、干燥、高温以及氧化等[1-3]。在耐辐射异常球菌中存在dr1372(命名为drwH)基因位点,其编码产物属于植物干燥抗性相关的胚胎发育晚期丰富蛋白(LEA)第5C家族成员[1,4-5]。LEA5C蛋白具有较高疏水性、较低序列重复性、不稳定系数及小分子氨基酸含量低、热稳定性差等特性,因此是一类非典型LEA蛋白[6-8]。值得注意的是,几乎所有已发现的LEA5C蛋白都包含一个“水胁迫和超敏反应结构域(Water stress and hypersensitive response,WHy)”,该结构域与生物体耐受干旱等非生物胁迫密切相关[9]。
到目前为止,有关细菌WHy蛋白的实验性研究仅有两个,即耐辐射异常球菌DrwH蛋白[5]和来源于南极沙漠土壤宏基因组文库的dWHy1蛋白[10]。研究报道,dWHy1蛋白N端含有26个氨基酸组成的信号肽序列,完整野生型dWHy1蛋白在大肠杆菌中异源表达,其存活率高于只含有空载体的对照菌株;而缺失信号肽的截短蛋白dWHy1ΔSP能显著增强宿主细胞耐受冷冻胁迫的能力[10]。本研究前期对来源于耐辐射异常球菌的DrwH核心结构域蛋白Dr-WHy在大肠杆菌中的抗逆功能进行初步研究,其在大肠杆菌中表达能够增强宿主的氧化胁迫抗性。此外,氧化胁迫(H2O2)条件下,Dr-WHy蛋白对苹果酸脱氢酶(MDH)和乳酸脱氢酶(LDH)的酶活性有保护作用[5]。DrwH从耐辐射异常球菌中发现,对其非生物胁迫条件下功能的研究显得尤为重要。
大量来源于植物或微生物的LEA/WHy蛋白都是以大肠杆菌为底盘生物进行研究[11-12]。Liu等[13]采用分段截短及人工合成的方法研究来源于大豆的LEA3家族蛋白PM2的抗逆功能区域,确定了22-mer的重复序列在PM2蛋白增强大肠杆菌耐受高盐胁迫过程中起主要作用。Zou等[14]通过类似的分段截短方法对大豆LEA1家族蛋白Em进行分析,分别构建了N末端区域(Em-N)、中间区域(Em-2M)及C末端区域(Em-C)的截短蛋白,研究证明了在体外冷冻胁迫条件下不同结构域对LDH的保护能力(Em-C >Em-2M >Em-N)。尽管前期已对 DrwH 蛋白进行了初步研究,但其一级结构上某些特征序列是否具有类似于dWHy1中信号肽的功能,对核心结构域蛋白WHy的表达产生影响值得探讨。本研究以大肠杆菌为宿主菌株,比较分析drwH基因不同区域构建的重组菌株抗氧化能力的强弱,进一步揭示DrwH类信号肽序列对其抗氧化功能的影响。DrwH类信号肽功能的初步研究,将为进一步探索耐辐射异常球菌的氧化胁迫抗性分子机制提供新的思路。
1 材料与方法
1.1 材料
1.1.1 菌株、质粒与培养条件 本研究所需要的菌株与质粒如表1所示。大肠杆菌于LB培养基(Tryptone 10 g/L,Yeast Extract 5 g/L,NaCl 10 g/L,pH 7.0,固体培养基中加入agar 15 g/L)中37℃条件培养。含外源质粒的重组大肠杆菌,培养过程中加入相应的抗生素(如Kan 50 μg/mL)。
1.1.2 主要试剂 常规PCR反应试剂(PrimeSTAR HS DNA polymerase、dNTPs、Ex Taq酶)购自TaKa-Ra公司;限制性核酸内切酶、T4 DNA连接酶等购自NEB公司;细菌DNA提取试剂盒、质粒小量快速抽提试剂盒、琼脂糖凝胶DNA纯化试剂盒均购自Magen公司;细菌RNA提取试剂盒购自Promega公司;30% H2O2,所用其他试剂均为分析纯。引物合成与测序均由华大基因公司完成。
1.2 方法
1.2.1 生物信息学分析 使用SignalP 4.1、Phobius等在线软件进行蛋白信号肽预测分析。
1.2.2 重组大肠杆菌构建 本研究采用常规PCR克隆、酶切、连接、转化方法构建重组大肠杆菌菌株,将目的基因连接于pET28a表达载体,转化E. coliBL21(DE3)获得重组菌株。具体方法参照文献[5]。重组质粒构建所需引物如表2所示,SP-WHy基因由金唯智生物科技公司合成。
1.2.3 重组大肠杆菌抗氧化胁迫实验 将实验菌株在LB平板(50 μg/mL Kan)上分别划线过夜培养,从活化的平板上分别挑取单菌落于3 mL 新鲜液体培养基(50 μg/mL Kan)中37℃过夜培养,按初始OD600为0.05分别接种于20 mL的新鲜LB液体(50 μg/mL Kan)培养基中,培养30 min后加入终浓度为0.1 mmol/L IPTG于培养基中,连续培养至菌液OD600约为0.5,分别取出1 mL进行氧化胁迫处理。向上述1 mL菌液中加入15 mmol/L H2O2,处理10 min后立即10倍梯度稀释(10-1-10-4),每个稀释梯度各取10 μL点在LB固体培养基表面,经37℃恒温培养16 h后观察菌落形成情况。

表2 本研究所用引物列表
1.2.4 重组大肠杆菌生长曲线测定 从过夜培养的平板上挑取重组菌株BL21-DrwH、BL21-WHy及对照菌株BL21-pET单菌落,接种于新鲜的20 mL LB液体培养基中(50 μg/mL Kan),37℃、220 r/min过夜培养。按初始OD600为0.05分别将种子液接种于50 mL新鲜LB液体培养基中(每个样品3个重复),37℃、220 r/min摇床培养,每隔1 h取样,测定OD600值,连续测定至细菌生长稳定期。分别以培养时间和OD600吸光值为横纵坐标,绘制菌株的生长曲线。
1.2.5 qRT-PCR分析基因转录水平 接种活化的BL21-DrwH、BL21-WHy菌株于新鲜的20 mL LB液体培养基中,37℃、220 r/min摇床培养30 min后加入终浓度为0.1 mmol/L IPTG于培养基中(对照不加IPTG),连续培养至菌液OD600约为0.5,离心收集菌体,使用Promega RNA试剂盒提取菌体总RNA,并反转录为cDNA,通过qRT-PCR分析基因的表达水平。每个实验3次重复,取平均值。
2 结果
2.1 DrwH蛋白信号肽预测分析
耐辐射异常球菌基因组数据分析显示,drwH基因位于耐辐射异常球菌R1基因组Ⅰ号染色体上,该基因全长495 bp,编码164个氨基酸。通过SignalP 4.1和Phobius在线软件对蛋白信号肽进行预测显示(图1),DrwH蛋白的N端含有19个氨基酸组成的信号肽类似序列,该类信号肽序列组成一个α螺旋结构。
使用DNAMAN对来自植物、细菌和古细菌的48种LEA5C家族蛋白序列比较分析,结果显示耐辐射异常球菌DrwH蛋白的WHy结构域与Deinococcus属同源蛋白的WHy序列一致性最高(41.9%-61.6%),而与其他物种来源的WHy序列一致性仅为20%左右。同时对这48种LEA5C家族蛋白进行信号肽预测,发现有18种蛋白存在预测的信号肽,其中所有Deinococcus属来源的蛋白中都存在预测的信号肽序列(11个),所有细菌和部分古细菌中总共有7个含有预测的信号肽,而植物来源的LEA5C蛋白中均不含预测的信号肽(表3)。
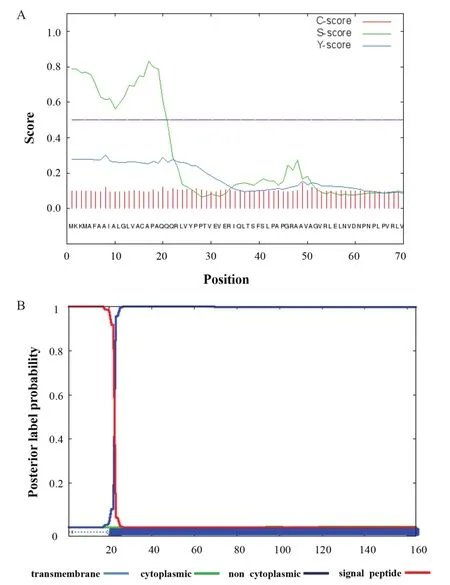
图1 采用SignalP 4.1和Phobius软件预测DrwH信号肽
2.2 重组大肠杆菌的构建
根据信号肽和结构域预测显示,DrwH蛋白N端含有19个氨基酸组成的类信号肽和一个WHy核心结构域,因此对DrwH蛋白的不同结构区域进行分段研究(图3-A),分别构建了野生型全长蛋白DrwH、C端截短蛋白DrwHΔC、N端截短蛋白DrwHΔN、核心结构域蛋白WHy、类信号肽SP、类信号肽SP缺失蛋白DrwHΔSP及类信号肽与WHy的重组蛋白SP-WHy,对其编码基因片段通过BamHI/HindIII双酶切消化后,分别连接于pET28a质粒,获得重组载体,转化E. coliBL21(DE3),构建的重组菌株分别命名为BL21-DrwH、BL21-DrwHΔC、BL21-DrwHΔN、BL21-WHy、BL21-SP、BL21-DrwHΔSP和BL21-SP-WHy(表1),重组菌株的构建策略如图2-A所示,重组载体通过PCR、酶切及测序验证正确(图2-B显示28a-drwH和28a-WHy的验证结果)。
2.3 氧化胁迫抗性分析
为探索DrwH蛋白不同区域对其抗氧化功能的影响,本研究对构建的BL21-DrwH、BL21-DrwHΔC、BL21-DrwHΔN、BL21-WHy重组大肠杆菌菌株和BL21-pET对照菌株分别进行氧化胁迫处理(图3-B)。结果显示,15 mmol/L H2O2胁迫处理10 min后,表达截短蛋白DrwHΔN和结构域蛋白WHy的重组菌株与对照菌株BL21-pET相比,存活能力提高了近3个数量级,而表达全长蛋白DrwH和截短蛋白DrwHΔC的重组菌株在H2O2处理后生存能力与对照菌株都对氧化冲击极为敏感。值得注意的是,在未胁迫处理的实验组,表达DrwH和DrwHΔC蛋白的重组菌株生存能力与其他菌株相差至少一个数量级,而这两种菌株均含有类信号肽序列SP。
结合生物信息学预测结果分析,由于全长蛋白DrwH和截短蛋白DrwHΔC的N端都含有19个氨基酸的类信号肽,为进一步验证类信号肽与大肠杆菌生长受抑制之间的关系,本研究构建了BL21-SP、BL21-DrwHΔSP及BL21-SP-WHy(图3)。对构建的重组大肠杆菌菌株进行15 mmol/L H2O2冲击10 min,结果显示,只要含有类信号肽SP重组蛋白的菌株,生长能力均受到不同程度抑制,表明DrwH蛋白的N端19个氨基酸组成的类信号肽序列是影响重组大肠杆菌生长的主要原因。此外,进一步表明DrwH发挥抗氧化功能的主要区段是核心结构域WHy。
2.4 重组菌株生长曲线测定
通过在正常培养条件下(LB培养基),测定重组菌株BL21-DrwH、BL21-WHy及对照菌株BL21-pET的生长曲线,进一步分析类信号肽SP对重组蛋白的影响。结果显示,所有的实验菌株在0-2 h生
长时期没有变化,随着培养时间的延长,OD600大于0.5时,表达结构域蛋白WHy的BL21-WHy与对照菌株没有明显差异,而过量表达全长蛋白DrwH的BL21-DrwH菌株生长明显受到抑制,最大OD600仅为0.8左右(图4)。实验结果表明在大肠杆菌中只要表达包含N端35个氨基酸(其中含有类信号肽序列SP)的重组蛋白,就会影响大肠杆菌菌株的正常生长。
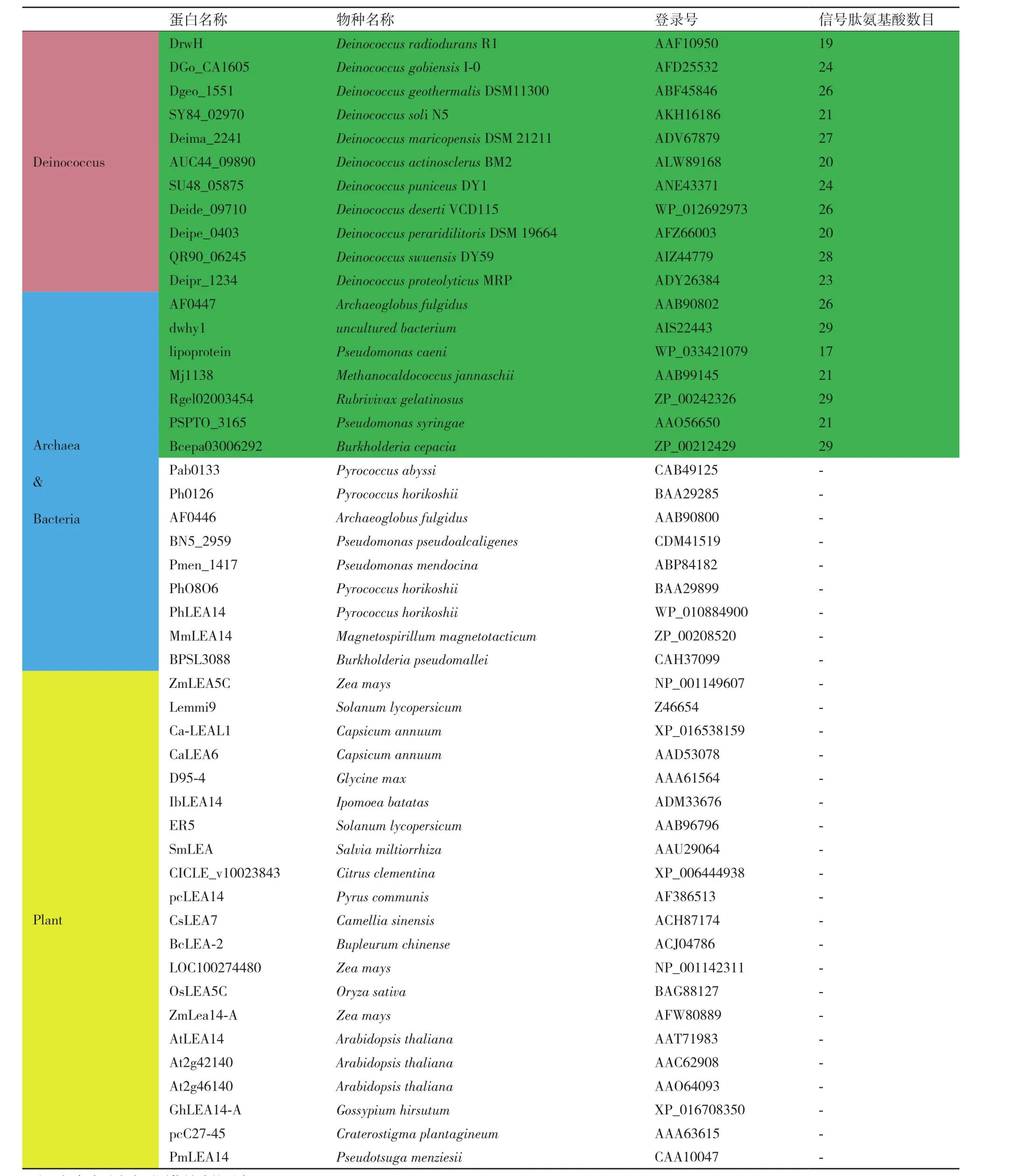
表3 LEA5C家族蛋白信号肽预测分析
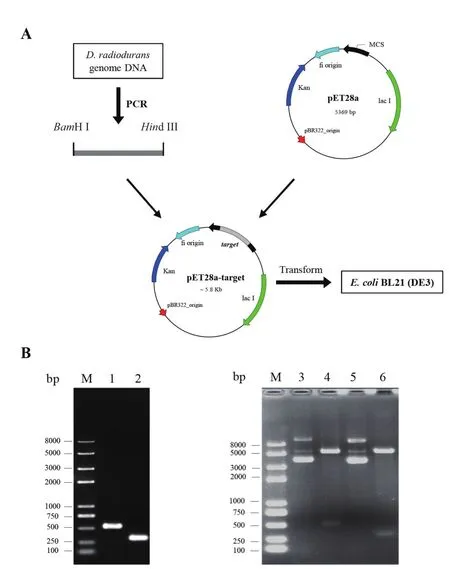
图2 重组大肠杆菌构建(A)与验证(B)
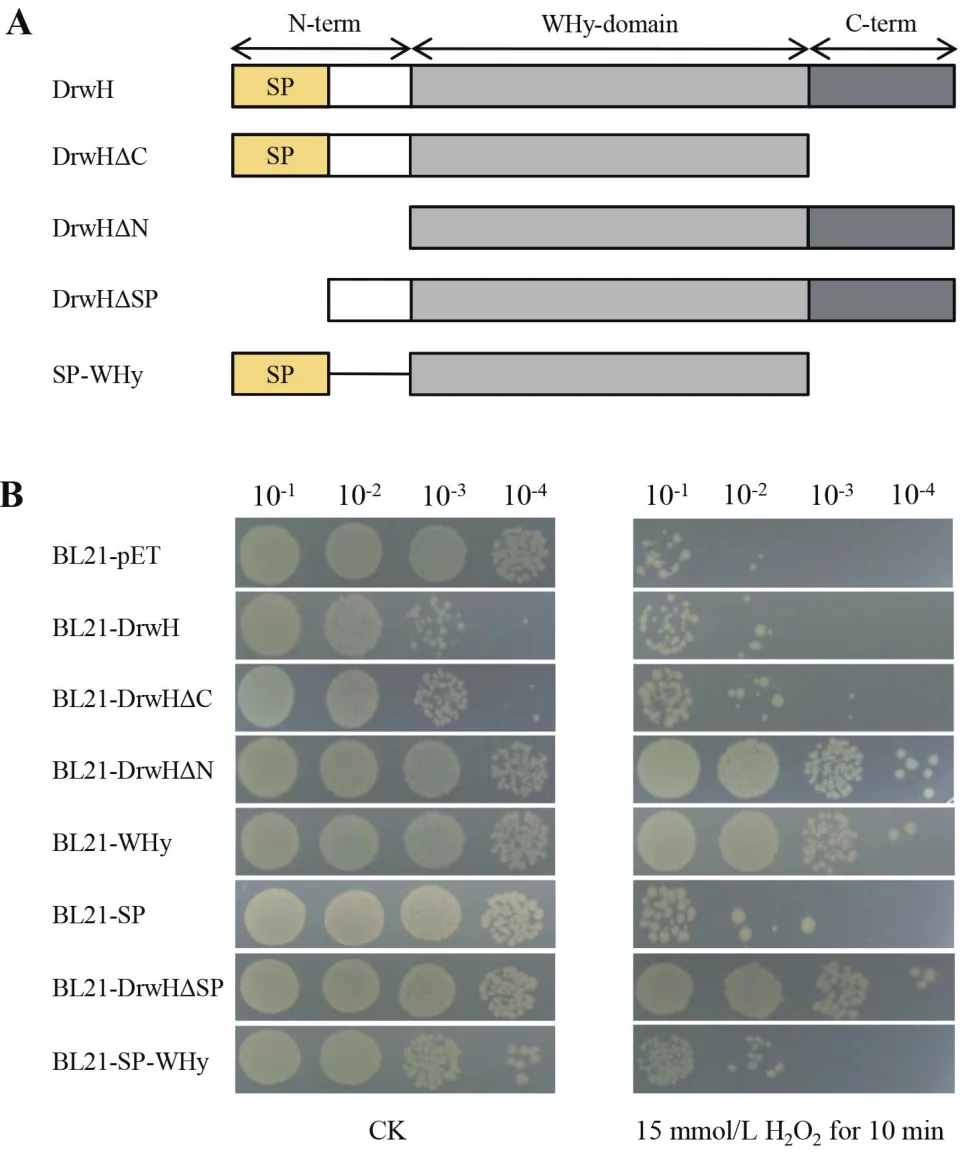
图3 表达DrwH不同截短蛋白重组菌株的抗氧表型分析
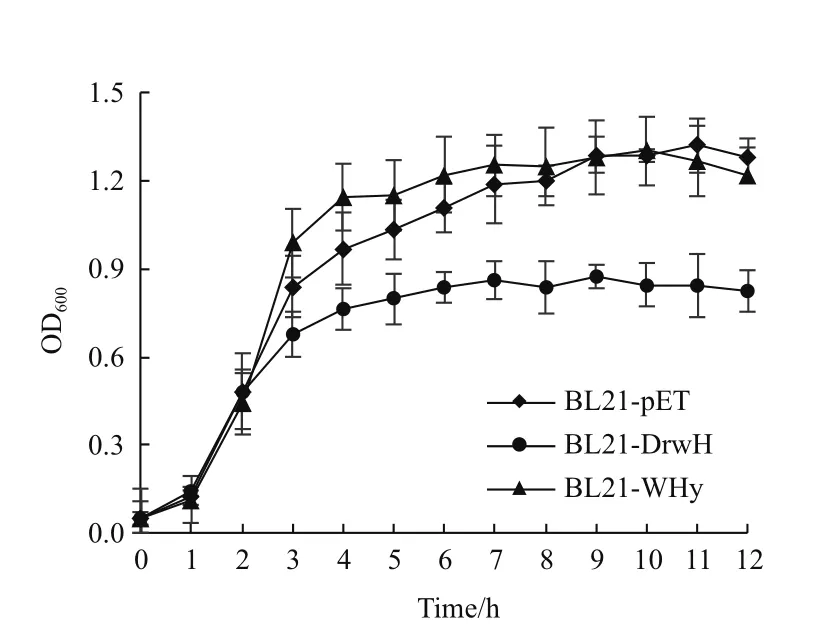
图4 过量表达全长DrwH和核心结构域WHy蛋白的重组大肠杆菌生长曲线
2.5 蛋白表达量对重组菌株抗氧化能力的影响
此外,本研究对未诱导(未添加IPTG)的重组菌株BL21-DrwH、BL21-WHy及对照菌株BL21-pET使用15 mmol/L H2O2处理10 min,结果显示,氧化胁迫处理后菌株生长能力均下降3个数量级,且所有菌株在正常培养和胁迫处理条件下均无差异,对氧化胁迫的抵抗能力也没有差别(图5-A),表明蛋白的表达量对重组大肠杆菌菌株的生长及抗氧化能力有显著的影响,细胞中蛋白量累积到一定水平才能发挥生物学功能,同时也证明了类信号肽SP的毒性效应也是在蛋白过量积累后出现的。通过qRTPCR分析了全长DrwH和核心结构域WHy编码基因在mRNA水平上的表达量(图5-B),结果显示经0.1 mmol/L IPTG诱导后,drwH和WHy基因的mRNA表达量比未经IPTG诱导的表达量均显著上调。以上结果表明,DrwH和WHy蛋白的表达量积累到较高水平才能发挥功能,同时也说明核心结构域N端的类信号肽序列SP对细菌的生长有影响。
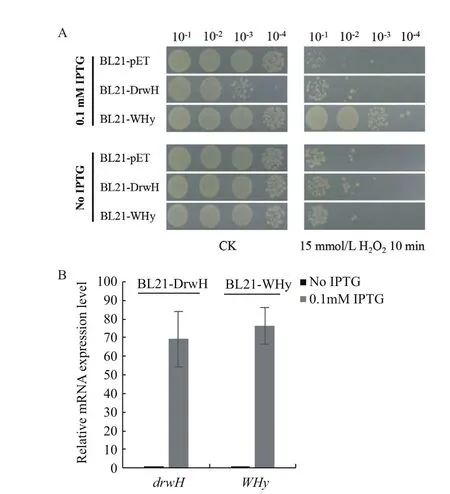
图5 蛋白表达量对重组菌株抗氧化能力的影响
3 讨论
本研究对耐辐射异常球菌DrwH蛋白信号肽预测分析发现,该蛋白N端含有19个氨基酸组成的信号肽序列,基于细菌中存在信号肽的说法目前还有待考证,本研究提出类信号肽序列SP的命名。通过对DrwH蛋白分段截短并构建含有不同区段的重组大肠杆菌,比较分析重组菌株在正常培养和氧化胁迫下的生长能力,发现含有类信号肽序列SP的全长DrwH蛋白过量表达对大肠杆菌的生长具有明显抑制作用,类似于一种毒性效应,这与Anderson等[10]对dWHy1信号肽的研究结果一致。利用qRTPCR分析重组菌株BL21-DrwH中drwH基因转录水平的变化发现,与未诱导的对照菌株相比,其表达量上调近70倍,表明drwH基因在mRNA水平受诱导。因此我们推测,DrwH在蛋白水平过量积累会对宿主细胞产生毒性,抑制其生长。
预测分析显示植物来源的LEA5C家族蛋白均不含信号肽序列,古细菌中也只有少数几个蛋白存在,而在所有的细菌LEA5C蛋白中都包含19-29个氨基酸组成的类信号肽,相比植物庞大的基因组而言,类信号肽序列在细菌相对紧凑的基因组中存在更显其重要性。研究显示,几乎所有LEA5C家族蛋白都含有WHy结构域,且WHy广泛存在于植物中,在少数细菌和古细菌中也有发现[9]。这个零散的分布可能是由于进化过程中,微生物通过结构域的选择性保留以应对胁迫环境而发挥功能[4],或通过从植物到原核生物的水平转移获得[9]。结合WHy结构域进化的可能性,类信号肽在细菌和少数古菌LEA5C蛋白中存在,我们推测这些具有特殊功能的序列是在微生物进化过程中选择性保留以应对环境胁迫[9],而植物因其基因组庞大,结构复杂,在进化过程中并不需要这样的类信号肽序列。另外,预测的19个氨基酸的短肽也可能作为调控序列,对细胞内DrwH蛋白的表达水平进行调节。根据以上分析,结合全长drwH基因突变导致耐辐射异常球菌对氧化和盐胁迫极为敏感的现象,我们可以确定,在耐辐射异常球菌中必然存在一种DrwH蛋白的剪切机制,在转录水平或翻译水平将未成熟蛋白加工修饰为成熟蛋白,从而发挥抗逆功能。
到目前为止,研究者对信号肽或类信号肽的存在与基因功能的相关性开展了一些研究。徐妙云等[15]对大豆24 kD油体蛋白基因在大肠杆菌中的表达研究表明,前22个氨基酸组成的信号肽可能影响了油体蛋白基因在细菌中的表达。Nguyen等[16]在P.pastorisX33中克隆了一个去除信号肽的内皮几丁质酶基因mchit1,并使其表达分泌,发现信号肽缺失能够提高酶的产率。Karlsson等[17]对革兰氏阴性菌中TonB蛋白N端的信号肽序列研究表明,信号肽序列与蛋白TonB在内外膜之间的连接有关。彭蔚宇等[18]对鸡白细胞介素10的信号肽体外表达分析发现,去除信号肽的重组质粒在胞浆中表达大量较强的绿色荧光,而含有信号肽的质粒荧光较弱且含量较少。Liu等[19]发现载脂蛋白M(apoM)未被切割的信号肽延缓了apoM向细胞外的转运。Wang等[20]研究发现信号肽的优化可以提高1,4-β-甘露糖苷酶活性。同样,本研究类信号肽序列过量表达对DrwH蛋白发挥功能产生不利影响,但对该类信号肽抑制DrwH蛋白发挥功能的分子机制仍不清楚。
4 结论
本研究分析显示耐辐射异常球菌DrwH蛋白含有19个氨基酸组成的类信号肽序列,对DrwH蛋白进行分段截短及重新组装,构建重组大肠杆菌菌株,并观察不同菌株在正常培养及氧化胁迫条件下的生长情况。结果表明,含有类信号肽序列的融合蛋白过量表达不仅显著抑制了大肠杆菌的生长,而且使WHy核心结构域失去抗氧化功能。qRT-PCR结果显示含类信号肽全长蛋白DrwH和结构域蛋白WHy在转录水平大量累积,诱导表达实验表明蛋白在过量积累条件下发挥其生物学功能。
猜你喜欢
天津农学院学报(2024年1期)2024-04-07 11:20:56
河北科技大学学报(2023年5期)2023-11-09 01:44:44
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
国际呼吸杂志(2019年1期)2019-01-28 09:37:06
浙江农业学报(2017年3期)2017-04-08 02:39:02
现代检验医学杂志(2016年2期)2016-11-14 02:38:02
环境科技(2016年3期)2016-11-08 12:14:10
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
华东理工大学学报(自然科学版)(2015年4期)2015-12-01 04:00:23
