
PPtase高表达菌株Streptomyces ghanaensis的发酵及其产物cytoxazone的发现
2019-06-01 07:03:36黄晓伟闫晓丽田文雅瞿旭东
中国抗生素杂志 2019年5期
黄晓伟 闫晓丽 田文雅 瞿旭东
(1 华中科技大学同济医学院附属同济医院,武汉 430030;2 武汉大学药学院,武汉 430072)
天然产物对新药的发现和发展至关重要。过去30年间,国际上被批准的小分子药物中有50%以上来源于天然产物及其衍生物,抗肿瘤和抗感染(抗细菌、真菌、寄生虫和病毒)的比率更是高达79.8%和69.8%[1]。然而由于长期缺乏新的研究方法和研究思路,新结构的活性天然产物发现率愈来愈低;加之新的代谢性疾病及多重耐药菌的发展快于新活性化合物的临床应用,医药领域对新的活性天然产物愈发迫切的需求,已成为当前药物研发领域最需解决的瓶颈之一[2]。另外,从天然产物中发现新的活性化合物的方式也是化学合成所不能替代的[3-4]。因此,从天然产物中寻找具有新颖结构和显著活性的化合物,仍将是更快捷、更有效的方式。
基因组学研究表明,目前已鉴定的微生物天然产物大约只占微生物生产能力的10%,大量的天然产物由于在实验室条件下处于“沉默”状态或微量表达而有待发掘[5-6]。随着对天然产物生物合成机制了解的日渐深入,发现天然产物的“沉默”状态或微量表达主要是由于在其生物合成基因的表观遗传、转录、翻译以及蛋白后修饰环节的调控问题导致的[7]。基于这些环节,近年来国际上发展了一系列有效的方法,在激活“沉默”天然产物生物合成基因簇("silent" or "cryptic" gene cluster)中取得了巨大成功[8]。其中,本研究发现了微生物体内磷酸泛酰巯基乙胺转移酶(phosphopan-tetheinyl transferases,PPTase)过强或过弱的修饰次级代谢和初级代谢载体蛋白(carrier protein, CP)是导致天然产物沉默的重要因素[9]。通过向33株放线菌中过量表达来自枯草芽胞杆菌(Bacillus subtilis)和黄萎病链霉菌(Streptomyces verticillus)的PPtase基因sfp和svp,共有23株菌株(70%)产生了一系列原先“沉默”的天然产物。对其中的一些菌株的激活产物的分离鉴定,我们发掘到了包括II型聚酮(type II polyketide) oviedomycin、I型聚酮(type I polyketide)化合物halichomycin和defumarylhygrolidin以及核苷类化合物puromycin等一系列沉默的天然产物[9-10]。为了进一步发掘和鉴定未知激活的天然产物,本研究中,我们对前期获得的一株链霉菌重组菌株Streptomyces ghanaensisCGMCC 4.1967-PPtase中的一个未知激活组分的分离,通过核磁鉴定发现了其为一种高效的细胞因子抑制剂cytoxazone。
1 实验部分
1.1 材料与仪器
仪器:LC-20AT型岛津高效液相色谱(HPLC,Shimadzu公司),C18分析柱(Agilent Eclipse XDB-C18,5μm,4.6mm×250mm,美国Agilent公司);核磁仪(Bruker BioSpin GmbH,400MHz德国Bruker公司);ZWY-2102C型摇床(上海智城分析仪器制造有限公司),SPX-70BIII生化培养箱(天津市泰斯特仪器有限公司)。试剂:均为分析纯试剂,购于国药集团化学试剂有限公司。
1.2 培养基和化学试剂
TSB(100mL):3.0g 胰蛋白胨大豆肉(Tryptone Soya Broth, TSB),去离子水定容至100mL。MS(100mL):2.0g 甘露醇(Mannitol),2.0g 黄豆粉(Soy Flour),自来水定容至100mL。阿泊拉霉素(Apramycin,购自Sigma公司)。
1.3 实验方法
1.3.1 发酵及HPLC检测条件
从-30℃低温保存冰箱中取出保种用的甘油管,在无菌操作台中,取出20μLS.ghanaensisCGMCC 4.1967-PPTase菌液[9]接种于含有相应浓度的阿泊拉霉素抗生素的4mL TSB培养基中,于30℃、220r/min摇床培养至一定浓度,然后吸取200μL菌液转接入250mL MS液体培养基中(共9L),置于28℃,220r/min摇床中,发酵培养7d后取出。将等体积的乙酸乙酯加入发酵培养基中,超声萃取15min,充分静置分层后,取出乙酸乙酯层,反复萃取3次,旋转浓缩得发酵浸膏9.5g。称取少量浸膏,用甲醇溶解,溶液用0.22μm的有机相微孔滤膜过滤,用于HPLC检测,检测条件:分析条件为T=0min,5%B;T=50min,100%B;T=50.01min,5%B;T=60min,5%B。其中A相:超纯水;B相:乙腈;流速为1mL/min;检测波长为254nm;柱型号:Agilent Eclipse XDB-C18。
2 结果与讨论
2.1 分析S.ghanaensis CGMCC 4.1967-PPTase发酵液组分
前期的研究过程中我们获得了一株sfp和svp过表达的菌株S.ghanaensisCGMCC 4.1967-PPTase,该菌株相对于野生型菌株S.ghanaensisCGMCC 4.1967能够激活产生聚酮化合物halichomycin和defumarylhygrolidin[9]。进一步对比其与野生菌株的发酵液组分,我们发现在检测波长为254nm的条件下,保留时间为16.8min (HPLC)处还产生了一个显著增强的峰(图1)。尽管在野生型菌株发酵液相应的保留时间处也存在着一个峰,但是两者之间的紫外吸收明显不同,属于不同化合物。

图1 HPLC分析S.ghanaensis CGMCC 4.1967野生菌株和PPTase重组菌株的发酵液组分Fig.1 HPLC analysis of metabolites production in S.ghanaensis CGMCC 4.1967 wild-type and PPTase strains
2.2 分离鉴定激活产物
为了鉴定该激活产物,我们对发酵并萃取得到的9.5g浸膏与甲醇混溶后硅胶拌样,以石油醚(PE):乙酸乙酯(EA),乙酸乙酯:丙酮作为流动相,以硅胶作为固定相,TLC为检测手段,按0:100~100:0的梯度进行洗脱,分离得9个组分(编号为S1~S9);其中,目标化合物主要集中于S9(石油醚:乙酸乙酯=90:10),分离流程见图2。目标化合物在S9组分静置过程中以白色无定型白色粉末形态析出,过滤分离以及反复溶解析出后可以得到纯净的化合物30mg。
2.3 化合物的结构鉴定
高分辨质谱分析确定该化合物正离子分子量为[M+H]+m/z224.0920,可以推出分子式为C11H13NO4。由1H (400MHz, CD3OD)(图3A),δH7.22(d,J=8.4Hz, 2H),δH6.96 (d,J=8.4Hz, 2H)。可知,该化合物含有一个1,4-双取代的苯环结构,这一点通过13C (101MHz, CD3OD)(图3B)δC127.8,δC113.6的高度是其他碳谱的2倍得以证实。由δH3.82 (s, 3H),δC54.4,δC159.9说明对位取代的基团是OMe。δC160.7说明含有一个羰基。由δC57.2,61.5,81.3可知这3个碳原子与杂原子相连,由δH5.00(d,J=8.4Hz, 1H)可知与该氢相连的碳原子与苯环相连(受苯环电子屏蔽效应,化学位移向低场位移),且邻位碳上含有一个氢原子,根据1H谱,该氢应是δH4.88(m, 1H),所连碳原子为手性碳原子,则另外两个氢原子δH3.20(m, 2H)所在的碳原子与其毗邻,通过与文献对比[11],确定该化合物为cytoxazone。
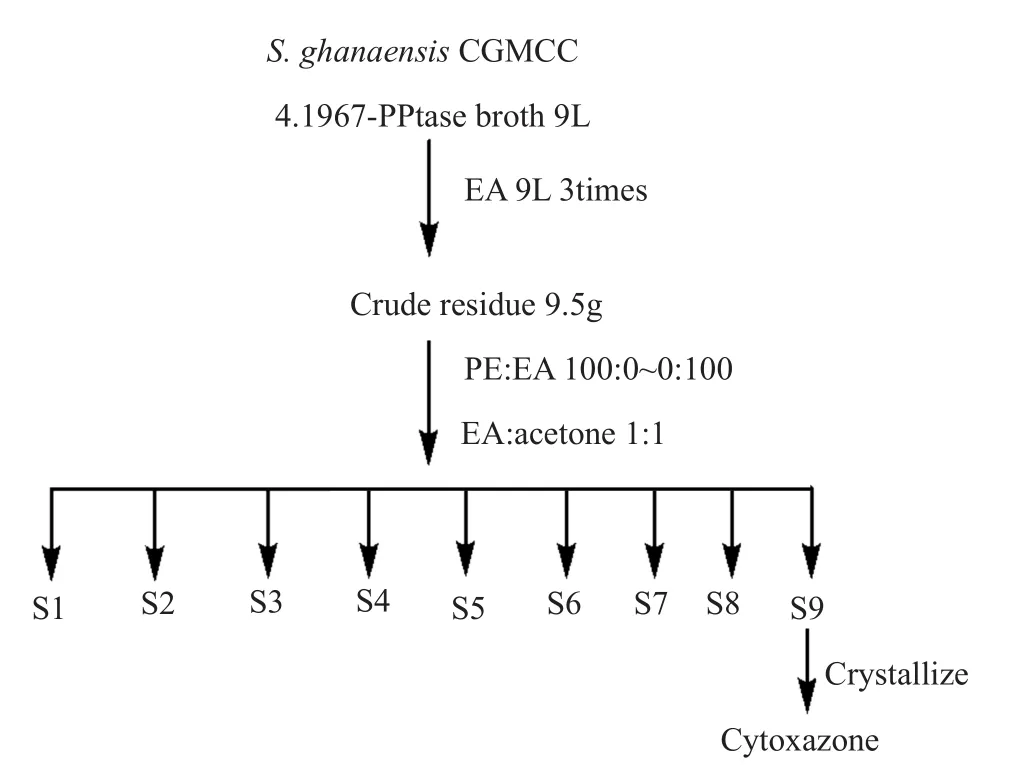
图2 Cytoxazone的分离纯化流程Fig.2 Purification process of cytoxazone
Cytoxazone能够选择性抑制Th2细胞(对Th1细胞无效)产生Ⅱ型细胞因子,因此具有很好的应用前景,可以用于治疗一系列细胞因子失调的疾病,如风湿免疫系统疾病,恶性淋巴瘤等[12]。此前,该化合物仅报道由一株分离自广岛的链霉菌(Streptomycessp.)产生[11],S.ghanaensisCGMCC 4.1967中为首次分离得到。

图3 Cytoxazone核磁谱图Fig.3 NMR spectra of cytoxazone
3 结论
本研究通过对一株过表达了PPtase基因的重组菌株S.ghanaensisCGMCC 4.1967的发酵产物进行分析,发现了一个新的激活产物。通过扩大培养,分离纯化和核磁鉴定,确定了该化合物为cytoxazone。该化合物是从嘎纳链霉菌S.ghanaensisCGMCC 4.1967首次分离得到,为后续研究其生物合成机制,合成生物学改造,以及开发新结构活性细胞因子抑制剂提供了坚实的基础。
猜你喜欢
中学生数理化(高中版.高考理化)(2024年2期)2024-03-20 01:30:20
现代仪器与医疗(2022年4期)2022-10-08 05:54:40
大学化学(2021年2期)2021-04-09 11:15:32
中小学班主任(2019年12期)2019-09-10 07:22:44
天然产物研究与开发(2018年4期)2018-05-07 06:47:53
中成药(2018年1期)2018-02-02 07:20:03
中学生数理化·高二版(2017年2期)2017-04-19 16:29:54
百姓生活(2016年6期)2016-06-22 14:39:00
科技创新导报(2016年1期)2016-05-30 06:38:19
实验室研究与探索(2015年3期)2015-02-21 06:26:55
