
大兴安岭北部天然针叶林土壤氮矿化特征
2019-05-31 01:06:08肖瑞晗满秀玲丁令智
生态学报 2019年8期
肖瑞晗,满秀玲,丁令智
东北林业大学, 哈尔滨 150040
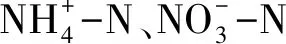
近年来,国际上对土壤氮矿化开展了广泛的研究[5- 7],其中包括森林土壤氮矿化速率变化特征[8],水、热因素对土壤氮矿化影响[9- 11]和植物对土壤氮矿化影响等[12]方面。虽然已取得了许多研究成果,但对土壤氮的动态认识依旧存在许多不确定性,在全球范围内没有一个统一的矿化模式。国内开展了许多模拟自然条件的室内控温、控湿实验,提出了相应的因子控制机理及理论矿化模型[13- 15]。在原位培养方面以温带草原、农田土壤、热带森林及亚热带温带森林研究[16-21]较多,而对于森林土壤氮矿化的研究则以人工林为主,较少研究天然林的土壤氮矿化特征,且很少考虑林地土壤氮矿化的垂直空间异质性,因而难以全面的分析和评价自然条件下土壤氮矿化过程。
大兴安岭北部是我国唯一的高纬度寒温带地区,冬季漫长寒冷,夏季短暂湿热,昼夜温差较大,月平均气温在0℃以下的月份长达8个月之久。本区地带性土壤是棕色针叶林土,土壤层较薄,多在20—30 cm,石块较多,且有多年冻土存在。由于气候寒冷,生长季较短,土壤微生物活动和有机质分解受到限制,因此土壤养分供给水平是本区植物生长重要的影响因子。本研究以大兴安岭北部地区天然针叶林为研究对象,探讨森林土壤无机氮含量、氮矿化特征及其影响因素,旨在揭示该地区土壤氮素转化特征和供给水平,为大兴安岭地区森林生态系统土壤养分管理及森林经营提供帮助。
1 研究地概况与研究方法
1.1 研究地概况
研究地位于大兴安岭地区漠河县境内黑龙江漠河森林生态系统国家定位观测研究站,地理坐标为122°07′—122°27′E,53°22′—53°30′N,该地区属寒温带大陆性气候,具有明显的山地气候特点,冬季由于受极地大陆气团控制,寒冷、干燥而漫长,夏季在副热带海洋气团的影响下,短暂湿热,年均温-4.9℃,多年平均降水量350—500 mm,多集中在7月份,全年无霜期80—90 d。研究区地带性植被为兴安落叶松林,另外还有樟子松林、樟子松-落叶松混交林、白桦(Betulaplatyphylla) 林、山杨(Populusdavidiana)林等森林类型,林下植物以兴安杜鹃(Rhododendrondauricum)和笃斯越桔(Vacciniumuliginosum)为主。地带性土壤为棕色针叶林土,局部地段分布有沼泽土和草甸土,并有永冻层存在。
1.2 样地设置和研究方法
在前期踏查的基础上选择本区典型针叶林为研究对象,即樟子松林、兴安落叶松林和樟子松-兴安落叶松混交林,在每种林型中选择典型地段布设3块20 m×20 m的样地,对样地进行基本情况调查,详见表1。每个样地每月选取3个培养点,采用顶盖埋管法[4]估测氮素的矿化速率。在实验开始前准备好长20 cm,内径5 cm的PVC管、塑料布、沙布和橡皮圈。每次取样时把3个PVC管垂直打入土中,然后小心取出装满土样的PVC管,再把PVC管顶部用塑料布封住,底部用纱布封住并用橡皮筋固定后放回原处;同时用另外的3个PVC管取原状土,将管内土壤分为0—10 cm和10—20 cm两层,分别装入袋中,带回实验室做室内分析。野外培养时间为30天,每次取样时先取出上次所埋管中的培养土,再用同样的方法埋管并取对应的原状土带回实验室。在每一样地的对角线上选3个50 cm×50 cm的样方,用钢板尺对枯落物的厚度测量并记录,同时用塑料袋收集样方内的枯落物及每个采样点(0—10 cm、10—20 cm)土壤,密封带回实验室分析。实验从2017年5月5日开始至2017年10月5日结束,共进行5个时段的培养。

表1 3种林型样地基本概况
1. 兴安刺玫; 2. 兴安杜鹃(Rhododendrondauricum); 3. 羽节蕨(GymnocarpiumJessoense); 4. 北国红豆(Vacciniumvitis-idaea); 5. 矮生悬钩子(Rubusclivicola); 6. 笃斯越桔; 7. 凤毛菊(Saussureaamurensis); 8. 红花鹿蹄草(Pyrolaincarnata); 9. 东方草莓(Fragariaorientalis); 10. 杜香(Ledumpalustre); 11. 越桔(V.vitis-idaea)

表2 3种林型样地土壤性质
氮矿化速率计算公式为:
M=[(I0′+I1′)-(I0+I1)]/D
(1)
M1=[(I0′-I0]/D
(2)
M2=[I1′-I1]/D
(3)
式中,M为净氮矿化速率、M1为净氨化速率、M2为净硝化速率,mg kg-1d-1;I0和I0′分别为培养前后铵态氮浓度,mgN/kg;I1和I1′为相对应的培养前后硝态氮浓度,mgN/kg;D为培养天数,d。
1.3 数据分析
2 结果与分析
2.1 土壤的变化特征
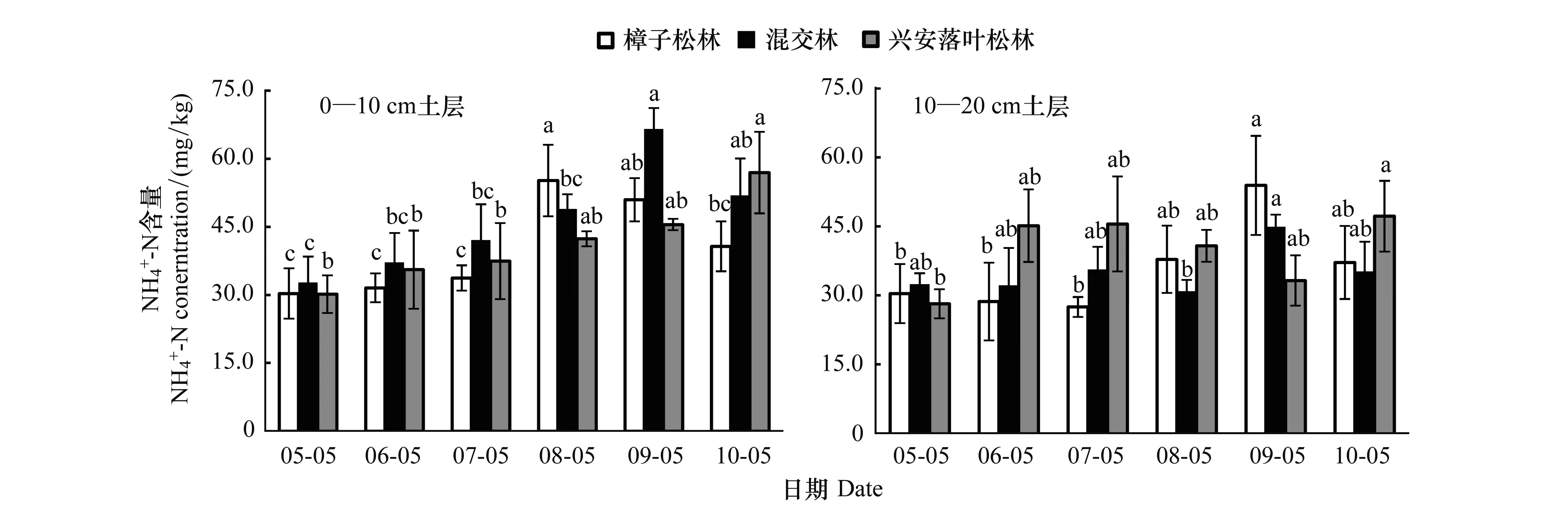
图1 3种林型月变化动态Fig.1 Monthly changes ofconcentration in the three forestsa、b、c和d为林型内不同月份土壤含量多重比较结果(P < 0.05)


图2 3种林型变化动态Fig.2 Monthly changes ofconcentration in the three forestsa、b、c和d为林型内不同月份土壤多重比较结果(P<0.05)
2.3 土壤净氮矿化速率变化特征
2.3.1净氨化速率
由图3可知,3种林型净氨化速率均呈V型变化趋势,且月变化显著(P<0.05),7、8月净氨化速率表现为负值,而其他月份则为正值。5—10月樟子松林、混交林及兴安落叶松林的净氨化速率变化范围分别为-0.34—0.89 mg kg-1d-1、-0.15—0.32 mg kg-1d-1和-0.51—0.60 mg kg-1d-1。3种林型不同土层净氨化速率存在显著差异(P<0.05),净氨化速率主要发生在0—10 cm土层,观测期内3种林型0—10 cm土层氨化作用占0—20 cm土层总氨化作用的65.3%—85.8%、57.5%—67.8%和59.3%—69.3%,而10—20 cm土层占比分别为14.2%—35.7%、33.2%—42.5%和30.7%—40.7%,其中樟子松林在6、7、9月0—10 cm土层净氨化率明显高于10—20 cm土层;混交林在6月0—10 cm土层净氨化率明显高于10—20 cm土层,而8月10—20 cm土层净氨化率明显高于0—10 cm土层(P<0.05)。
由此可见,整个观测期内3种林型0—10 cm土层净氨化速率明显高于10—20 cm土层(P<0.05),土壤的净氨化作用主要发生在0—10 cm土层,占整个0—20 cm土层的60%以上。
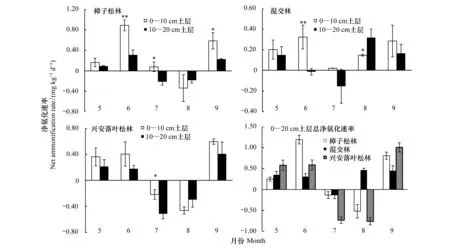
图3 3种林型净氨化速率月变化动态Fig.3 Monthly changes of net ammonium rates in the three forests图中*和**分别表示0—10 cm、10—20 cm土层净氨化速率差异显著(P<0.05)和极显著(P<0.01)
2.3.2净硝化速率
樟子松林10—20 cm土层净硝化速率月变化差异显著(P<0.05),其他均差异极显著(P<0.01)(图4)。3种林型净硝化速率最大值出现在6月,但混交林在9月又出现一次明显上升。3种林型净硝化速率在5、6、9和10月为正值,且0—10 cm土层净硝化速率大于10—20 cm土层(P<0.05); 7、8月净硝化速率产生波动出现负值,且出现10—20 cm土层净硝化速率大于0—10 cm土层。3种林型间土壤平均净硝化速率存在明显的差异(P<0.05),0—10 cm土层平均净硝化速率依次为:混交林((0.026±0.010)mg kg-1d-1)>兴安落叶松林((0.017±0.0028)mg kg-1d-1)>樟子松林((0.0014±0.011)mg kg-1d-1);10—20 cm土层平均净硝化速率大小依次为:樟子松林((0.023±0.010)mg kg-1d-1)>混交林((0.013±0.0064)mg kg-1d-1)>兴安落叶松林((0.0090±0.011)mg kg-1d-1)。3种林型0—20 cm土层净硝化速率变化范围分别为:-0.022—0.084 mg kg-1d-1(樟子松林)、-0.040—0.099 mg kg-1d-1(混交林)、-0.064—0.044 mg kg-1d-1(兴安落叶松林)。
综上所述,3种林型土壤净硝化速率除7、8月外其余月份为正值且0—10 cm土层明显高于10—20 cm土层(P<0.05),7、8月为负值且8月份10—20 cm土层高于0—10 cm土层(P<0.05)。3种林型间土壤平均净硝化速率存在明显的差异(P<0.05)。
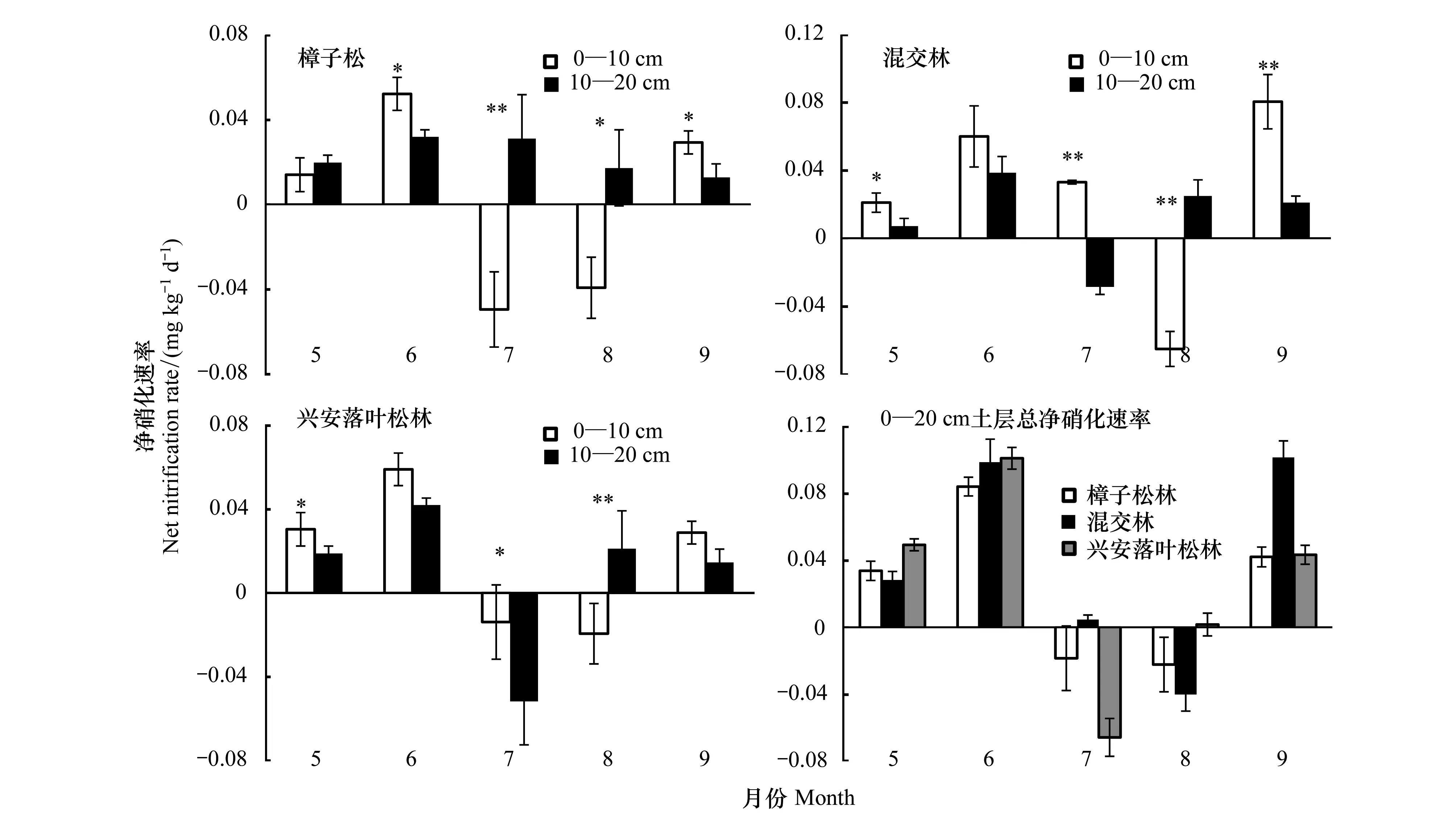
图4 3种林型净硝化速率月变化动态Fig.4 Monthly changes of net nitrification rates in the three forests图中*和**分别表示0—10 cm、10—20 cm土层净硝化速率差异显著(P<0.05)和极显著(P<0.01)
2.3.3净氮矿化速率
由图5可知,5—10月3种林型的净氮矿化速率变化趋势与净氨化速率趋势类似,即5、6月呈上升趋势,7、8月呈下降趋势且净氮矿化速率出现负值,9、10月又有所上升。樟子松林0—20 cm土层在6月净氮矿化速率出现最大值为(1.28±0.11)mg kg-1d-1,8月出现最小值为(-0.54±0.18)mg kg-1d-1;混交林和兴安落叶松林则在9月出现最大值,分别为(0.55±0.13)mg kg-1d-1和(1.05±0.12)mg kg-1d-1,在7月出现最小值分别为(-0.13±0.086)mg kg-1d-1和(-0.80±0.088)mg kg-1d-1。3种林型净氮矿化速率表现出显著的垂直差异性(P<0.05),但与土壤净氨化速率和净硝化速率相比其垂直差异性相对减弱。3种林型间平均净氮矿化速率差异不显著(P>0.05)。
由此可见,在观测期内3种林型土壤净氮矿化速率呈V型变化趋势且均表相出显著的垂直差异性(P<0.05)。但3种林型间平均净氮矿化速率无显著性差异(P>0.05)。

图5 3种林型净氮矿化速率月变化动态Fig.5 Monthly changes of net mineralization rates in the three forests图中*和**分别表示0—10 cm、10—20 cm土层净硝化速率差异显著(P<0.05)和极显著(P<0.01)
2.4 土壤和枯落物碳氮含量对土壤氮矿化速率的影响
由表3可以看出,不同土层净氮矿化速率影响因子有所不同,但总体而言土壤净氮矿化速率与土壤含水量呈二次相关关系,说明土壤水分含量的增加在一定程度上促进氮矿化的进程,但土壤水分含量超过一定阈值时会抑制土壤氮矿化的发生;土壤氮矿化速率与土壤有机碳含量呈显著正相关(P<0.05),土壤有机碳含量增高氮矿化速率也逐渐增加;净氮矿化速率与枯落物全氮和C/N呈显著负相关(P<0.05),说明枯落物中C/N高,氮素转化较慢,释放量少,进而影响氮的矿化速率。土壤净氮矿化速率与土壤pH值、土壤全氮和枯落物储量相关性不显著(P>0.05)。另外,0—10 cm土层净氮矿化速率还与土壤C/N呈显著正相关(P<0.05),与枯落物有机碳含量呈显著负相关(P<0.05)。
3 讨论
3.1 土壤无机氮变化特征
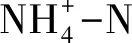

表3 净氮矿化速率与土壤基质和枯落物相关性分析
表中*和**分别表示相关性水平显著(P<0.05)和极显著(P<0.01)
无机氮含量标志着土壤中可供植物吸收利用的氮素水平,是评价土壤供氮能力的一项重要指标[26,38],是土壤直接供给植物吸收的养分,其含量显著影响着森林土壤的生产力。研究区土壤无机氮平均含量高于其他地区平均水平,说明该地区土壤具有较强的供氮能力。不同林型间无机氮含量及组成存在一定差异,这表明不同森林类型和生态环境氮素有效性有所差异[23]。土壤氮素供应取决于土壤有机质的积累与分解状况,与地上植被类型及其根系、凋落物输入与分解相关;与此同时,不同森林类型的土壤容重和土壤孔隙度等物理性质也影响土壤无机氮含量及其组成特征。大兴安岭地区由于其特殊的地理位置及气候特点使得该地区的生长季较短,较高的无机氮水平保证了土壤对植物有效的氮素供应,提高了这一地区森林土壤的生产力,能够为森林生态系统的持续经营提供养分保障。
3.2 土壤净氮矿化速率
3种林型土壤氮矿化过程存在明显的月际差异(P<0.05),且0—10 cm土层的氮矿化作用明显强于10—20 cm土层(P<0.05)。一般来说,土壤净氮矿化速率与土壤微生物活性有关,土壤微生物活性高有助于提高土壤净氮矿化速率,而微生物活性与土壤有机质和全氮含量有关[9- 41], 3种林型0—10 cm土层具有较高的有机质和全氮含量,为微生物的活动提供了保障,从而为土壤净氮矿化创造了良好的条件[24]。3种林型观测期内土壤净氮矿化速率均值变化范围樟子松林为-0.54—1.28 mg kg-1d-1、樟子松-兴安落叶松混交林为-0.13—0.55 mg kg-1d-1、兴安落叶松林为-0.80—1.05 mg kg-1d-1。可以看出,混交林矿化速率变化幅度较小,这与混交林土壤无机氮含量较高有关,土壤矿质氮含量升高能抑制土壤氮素矿化[39]。3种林型5、6月土壤矿化速率较高,为矿化的活跃期[24]。而7、8月份土壤氮矿化速率为负值,其原因可能与土壤中养分含量变化及土壤水分含量有关。当土壤中养分充足时,土壤氮素矿化作用弱于微生物对氮的固持作用,土壤中无机氮含量转为消耗性为主。反之,在其他月份土壤中养分缺乏,土壤氮素矿化作用明显高于微生物对氮的固持作用,土壤表现为无机氮素积累过程,土壤氮矿化速率增大[42]。
3种林型土壤净氮矿化速率在不同月份变化较大,且7、8月份矿化速率为负值,与其他地区土壤氮矿化速率变化趋势有所不同,而7、8月份为该地区降水比较集中的时期,说明在不同地区气候因素和环境因素大不相同,对氮矿化的影响也不同。森林土壤存在很大的垂直空间异质性,表层土壤氮矿化作用更强,这也说明土壤氮矿化速率是气候因子、林分组成、枯落物质量及土壤基质等多种影响因子综合作用的结果。
3.3 土壤净氮矿化的控制因子
影响土壤净氮矿化速率的因素很多也很复杂,周才平等对长白山两种林型下土壤氮矿化进行研究,发现与温度和湿度关系密切[13]。在本研究中,土壤净氮矿化速率与土壤水分间存在二次相关关系,即在较低土壤水分范围内,随水分增加,土壤净氮矿化能力增强,但当水分超过一定阈值后反而会抑制土壤净氮矿化过程[13-14,43]。土壤净氮矿化速率与土壤温度相关性不显著(P>0.05),这可能与大兴安岭地区寒冷的气候条件有关。土壤有机质含量和土壤C/N的提高会增强土壤净氨化、净硝化及净氮矿化速率[41,44]。土壤净氮矿化速率与枯落物全氮含量及C/N呈极显著负相关,C/N越大越难被矿化,说明枯落物通过影响土壤中的碳氮含量间接的影响土壤氮矿化速率[45]。而土壤氮矿化速率与枯落物储量却无显著相关性,这说明枯落物对土壤矿化的影响主要表现为质量控制,而非数量上的控制[41,46-47]。通过对0—10 cm及10—20 cm土层土壤氮矿化速率影响因子相关分析发现,0—10 cm土层氮矿化速率受土壤及枯落物碳氮含量及C/N影响较大,其原因可能是由于上层土壤养分较高,且其土壤容重较小,透气及透水能力较强有助于土壤氮矿化的发生。
4 结论
(2)3种林型生长季土壤净氮矿化、净氨化、净硝化速率均呈V型变化趋势,最小值在7、8月,且为负值。土壤净氨化过程是土壤氮矿化过程的主要形式,占比在60%以上。
(3)土壤氮矿化过程受多种因素的共同影响。土壤水分含量、土壤有机质含量、枯落物全氮含量及碳氮比在土壤氮矿化过程中起主要作用;而土壤温度、土壤pH、土壤全氮和枯落物数量及有机碳含量对氮矿化速率影响较小。森林生态系统土壤及枯落物的质量影响土壤氮矿化特征及植物对氮素的吸收利用。
致谢:东北林业大学黑龙江漠河森林生态系统国家定位观测研究站提供野外基础支持。
猜你喜欢
北京大学学报(自然科学版)(2022年4期)2022-08-18 06:57:34
草业学报(2021年9期)2021-09-22 08:55:16
河南畜牧兽医(2020年11期)2020-01-11 05:31:49
生态学报(2018年21期)2018-12-19 10:47:28
水土保持通报(2018年3期)2018-07-26 00:41:46
农业环境科学学报(2017年2期)2017-03-20 14:57:30
河北林业科技(2016年5期)2016-11-08 03:13:01
现代农村科技(2016年13期)2016-08-09 03:33:38
兽医导刊(2016年6期)2016-05-17 03:50:53
农村农业农民·B版(2015年9期)2015-10-16 16:11:41
