
甘蓝型油菜茎高QTL定位及株高相关位点整合
2019-05-31 06:45魏丽娟刘瑞影陈志友强李加纳
作物学报 2019年6期
魏丽娟 刘瑞影 张 莉 陈志友 杨 鸿 霍 强李加纳,*
1西南大学农学与生物科技学院 / 油菜工程研究中心, 重庆 400715; 2西南大学现代农业科学研究院, 重庆400715
甘蓝型油菜是重要的油料作物之一, 也是重要的饲料来源和工业原料。株型是指作物的形态特征和空间分布, 在作物育种过程中具有最明显的变化。理想株型可以提高植物叶面积指数, 进而影响植物的光合作用, 对植物生长发育和籽粒产量的形成具有重要的作用[1]。
株高是油菜重要的株型性状, 它与植株的倒伏和产量密切相关。主茎高度(茎高)和主花序长度是油菜植株高度的重要组成部分, 茎高包括分枝区段长度和一次分枝高度。杨光等[2]发现薹肥施用时期和施肥量对株高的构成因素产生了不同的影响, 推迟薹肥施用时期, 主花序长度增加, 而主茎高度降低。对甘蓝型油菜产量及株型相关性状的通径分析表明, 千粒重和主茎高度对经济产量的效应最大[3]。因此, 在甘蓝型油菜中, 研究主茎高度具有重要的意义。近年来, 研究者们通过QTL定位和全基因组关联分析, 将甘蓝型油菜株高、主花序长度、分枝区段长和一次分枝高度定位在19条染色体上[1,4-16], 并找到一些与株高相关的候选基因。Zheng等[12]对株高和一次分枝高度进行全基因组关联分析, 株高定位在 A02、A03、A06、A07、C02和 C03染色体上, 一次分枝高度定位在 A02、A07、A08和A09染色体上, 并在A02染色体上找到株高候选基因BnRGA和BnFT。Li等[13]利用60 K SNP芯片对甘蓝型油菜的株高进行了全基因组关联分析, 共检测出8个与株高相关的QTL, 它们分别分布在染色体A03、A05、A07和C07上, 进一步研究确定了3个与株高相关的候选基因, 其功能主要与赤霉素合成相关。然而未见甘蓝型油菜茎高定位的相关研究报道。
尽管株高相关性状定位方面的研究报道较多,但是由于作图群体和标记类型不同, 很难对不同QTL进行比较。Raboanatahiry等[17]和张江江等[18]对株高进行 QTL整合, 但未包含最新的研究报道,而且未对株高其他相关性状进行分析。本研究利用重组自交系和自然群体定位甘蓝型油菜茎高, 并整合株高相关性状, 找到茎高候选基因。
1 材料与方法
1.1 试验材料
重组自交系群体(甘蓝型油菜 GH06为母本, 黑籽油菜ZY821为父本, F2代连续自交10代以上, 形成含 186个株系的重组自交系群体)和自然群体(国内及国外的 455份甘蓝型油菜, 包括春性、冬性和半冬性甘蓝型油菜)均来自重庆市油菜工程技术研究中心。2015年9月至2016年5月(性状统计记录为2016)与2016年9月至2017年5月(性状统计记录为 2017)将亲本及重组自交系群体和自然群体连续 2年种植于重庆市北碚区歇马镇油菜基地, 随机区组设计 2个重复, 每小区3行, 每行10株, 行距40 cm, 株距 20 cm, 田间管理按照常规方式进行,确保所有材料的外部生长环境一致。
1.2 性状考察
成熟期取5株生长一致油菜, 测定茎高(子叶节到主茎最上部一个有效分枝着生点之间的距离)、主花序长度(主茎最上部一个有效分枝着生点到顶端之间的距离)、株高(子叶节到整株油菜最高部分的长度)、结荚高度(近地第一个有效角果的着生高度)、结荚厚度(最顶部第一个有效角果到近地第一个有效角果之间的高度差)、一次有效分枝起点(子叶节到主茎上最下部一个有效分枝之间着生点之间的距离)、经济产量(5株植株所有自然风干种子总重的平均值)。
1.3 表型数据分析
利用Microsoft Excel 2013对重组自交系和自然群体表型数据进行t测验和绘制正态分布图, 利用DPS 7.05统计分析软件对重组自交系和自然群体进行相关性分析和方差分析。
变异系数CV =σ/μ, 其中σ为标准差,μ为平均值。
广义遗传力h2=σ2G/(σ2G+σ2GE/e+σ2e/re), 其中σ2G表示基因型方差,σ2GE表示基因型与环境互作方差,σ2e为误差,e代表环境数目,r代表每个环境的重复数。
1.4 遗传图谱构建和QTL定位
取每个株系的5个幼嫩叶片混合, 提取DNA用于SNP标记分析。按照Illumina公司的Infinium HD Assay Ultra的说明进行DNA样品的预处理、与芯片杂交、洗脱、单碱基延伸、染色及包埋, 芯片准备好后利用 Illumina HiSCAN 扫描, 并利用GenomeStudio genotyping software v2011对扫描结果进行分析, 获得每个株系的基因型。利用MSTmap软件[19]构建 SNP遗传图谱, 取最小阈值LOD 5.0对所有标记进行分组, 通过标记与标记两两之间最小重组频率计算每个连锁群上的标记顺序。SNP遗传图谱包括8575个SNP标记, 1201个bin, 覆盖甘蓝型油菜基因组 6140.2 cM。利用WinQTLCart2.5[20]软件和复合区间作图(composite interval mapping, CIM)法对茎高进行QTL定位分析,LOD阈值设置为2.5, QTL的命名是斜体的小写字母“q”加上年份, 再依次加上性状名字、染色体和QTL序号。如q2016SH-C01-1表示 2016年茎高在 C01染色体上的第1个QTL。
1.5 全基因组关联分析
提取甘蓝型油菜 DNA, 构建插入片段大小为350 bp的DNA文库, 利用Illumina HiSeq 4000对甘蓝型油菜进行重测序分析, 双末端测序, 测序深度为5×, reads读长为150 bp, 测序数据提交到NCBI数据库中(BioProject: PRJNA358784)。使用 FastQC和 Trimmomatic[21]进行对原始数据处理, 利用Burrows-Wheeler软件[22]将clean reads比对到甘蓝型油菜参考基因组后, 使用 SAMtools’s mpileup (version 0.1.19-44428cd)软件[23]对每个样品进行SNP分析, 获得个 385,691个 SNP标记(MAF>0.05)。利用这些SNP标记, 对甘蓝型油菜自然群体进行群体结构和亲缘关系分析。利用structure软件分析甘蓝型油菜群体结构[24], 组群数目(K)设定为1~10,K值运行 3次(number of iterations)。MCMC (Markov Chain Monte Carlo)不作数迭代(length of burn-in period)和不作数迭代后的MCMC全部设为100,000次, 并选择混合模型和等位变异发生频率相关模型进行分析。
利用TASSEL软件[25]中的一般线性模型和混合线性模型, 以Structure运算后的Q值和亲缘关系K值作为协变量, 结合基因型数据, 进行全基因组关联分析。以每个SNP位点的-lg (P)观察值和期望值,绘制Quantile-Quantile散点图(QQ plot)。与性状显著关联的SNP位点的阈值为P<2.59×10-6, 计算方法为1/关联分析所用的所有 SNP数目(本研究中为385,691)。
1.6 株高相关性状位点整合
收集株高、主花序长度、分枝区段长度和一次分枝高度定位结果[1,4-16], 对于 QTL定位结果, 利用QTL区间侧翼引物与甘蓝型油菜基因组[26]比对, 得到其物理位置; 对于GWAS结果, 定位区间为显著关联SNP位点左右延伸500 kb。通过比较株高相关性状的物理位置, 整合株高相关性状和本研究定位结果到甘蓝型油菜物理图谱上, 并用Circos图[27]展示。
2 结果与分析
2.1 重组自交系和自然群体茎高表型变异分析
重组自交系群体中, 亲本ZY821和GH06的茎高分别为134.95 cm和152.60 cm,t测验结果表明,两亲本茎高差异未达到显著水平(P>0.05)。但是群体茎高在不同年份呈现广泛的变异, 变异范围为 114~175 cm, 变异系数为 7.48%, 广义遗传力为 83.3%;自然群体茎高表型变异范围为 62~202 cm, 变异系数为 12.87%, 其变异范围略高于重组自交系群体,广义遗传力为 86.5%(表 1)。茎高在2个群体中呈连续变异, 近似正态分布, 表明其受多基因控制(图1)。
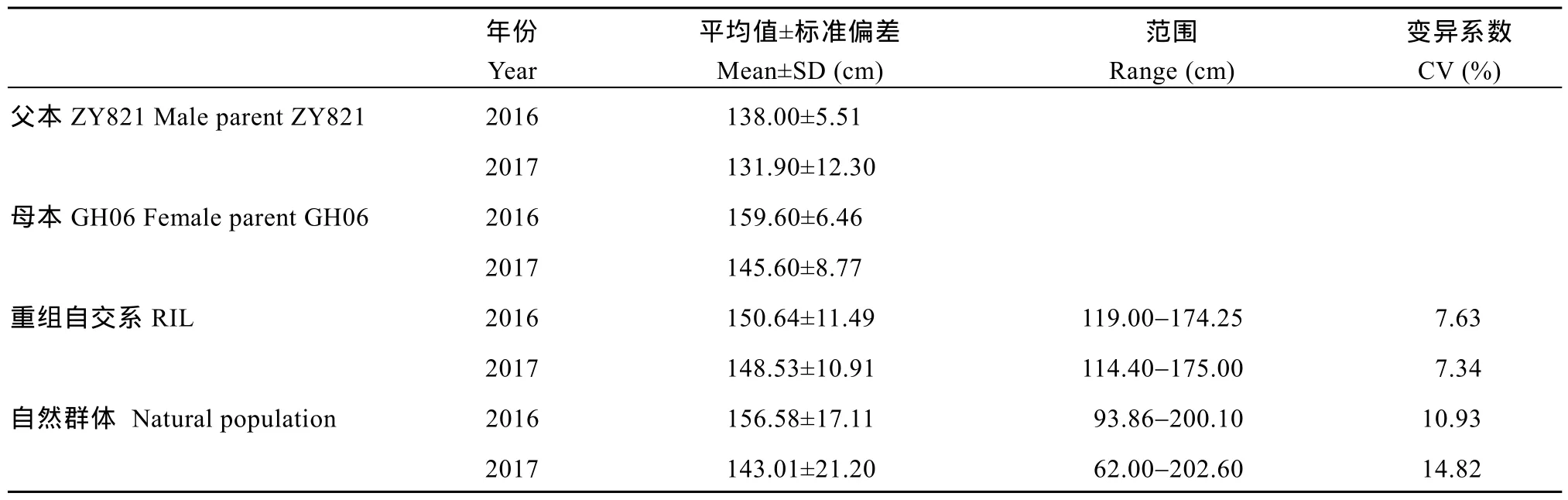
表1 重组自交系和自然群体甘蓝型油菜茎高的表型变异Table 1 Phenotypic variation of stem height in RIL population and natural population
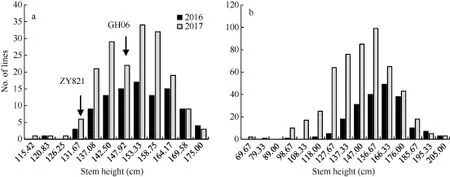
图1 重组自交系(a)和自然群体(b)茎高频率分布图Fig. 1 Frequency distribution of stem height in RIL (a) and natural population (b)
2.2 茎高与其他株型相关性状及产量的相关性分析
表 2表明, 重组自交系和自然群体结果几乎一致, 茎高与经济产量呈极显著正相关, 茎高对经济产量起着重要的作用; 茎高与株高、结荚高度、一次有效分枝起点呈显著正相关, 与结荚厚度不相关,表明茎高可以使株高、结荚高度和分枝起点提高, 却不能提高结荚厚度, 在生产实践中应适度控制茎高。
2.3 茎高QTL定位
采用复合区间作图法对茎高进行QTL定位分析,当LOD值≥2.5时, 2年共检测到11个茎高QTL, 分布在A04、A06、C04、A08、C01染色体上(表3和图2), 位点的表型贡献率为7.25%~19.61%, 在2年中没有QTL被重复检测到,q2016SH-A06-2位于A06染色体1.8~2.1 Mb处, 解释的表型最高, 为19.61%,加性效应值为负, 说明增效基因来自父本ZY821。
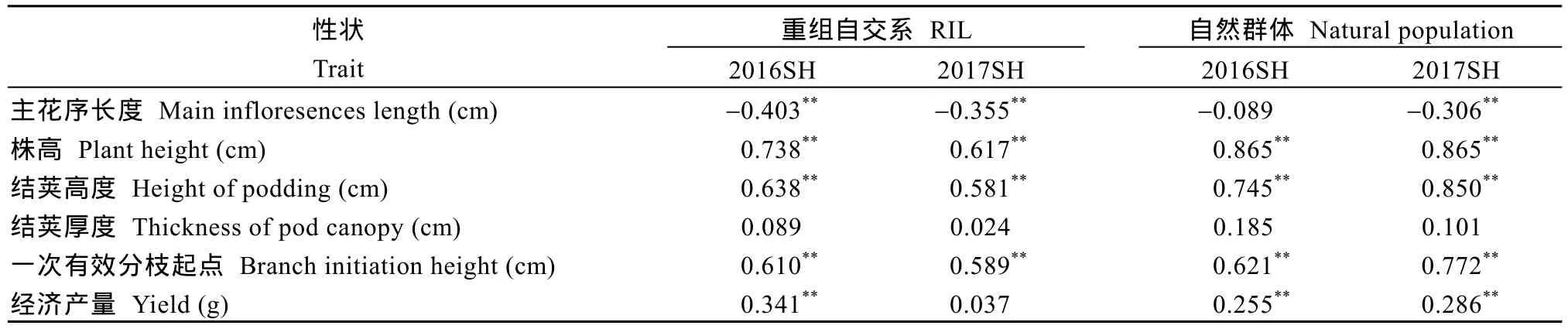
表2 甘蓝型油菜茎高与株型和经济产量性状相关系数Table 2 Correlation coefficients of stem height with plant architecture and yield traits in B. napus
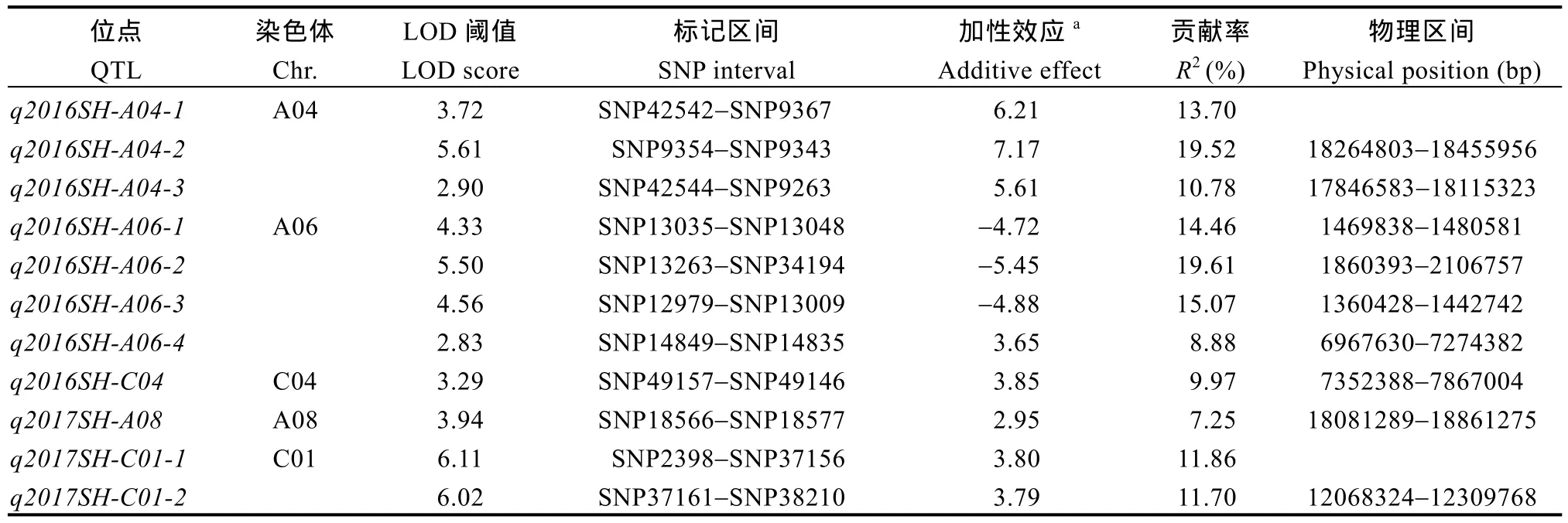
表3 甘蓝型油菜茎高QTL结果Table 3 QTL related to stem height in B. napus
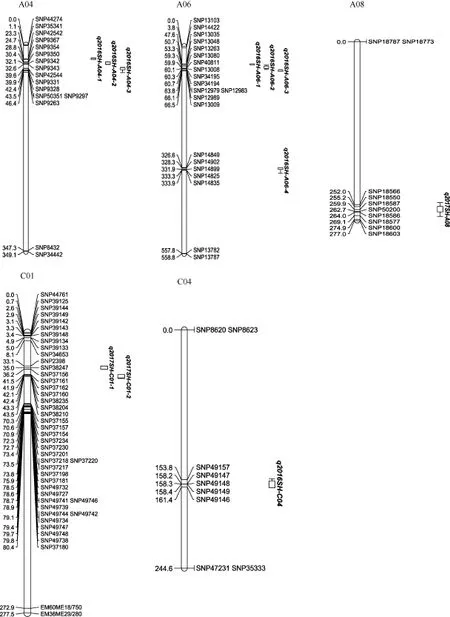
图2 甘蓝型油菜茎高QTL在连锁图上的位置Fig. 2 Chromosomal locations of QTL for stem height in RIL population
2.4 茎高全基因组关联分析
将 445份甘蓝型油菜种质资源群体划分为 P1和P2两个亚群(图3), 其中P1亚群包括398份材料,主要是我国半冬性油菜栽培种, P2亚群大部分是引进的国外种质资源材料以及少量春性油菜和半冬性油菜。
2016年茎高的QQ图(图4)表明, Naive模型检测到的P值接近期望值, 具有较好的控制假阳性能

图3 甘蓝型油菜群体结构图Fig. 3 Population structure of 455 B. napus
力; 2017年茎高的最优模型为K+PCA。全基因组关联分析2年共检测到5个SNP与茎高显著关联(表4), 分布在A08、A10、C02和C06染色体上。其中2016SH-1和2016SH-2解释的表型变异较高, 标记S8_12459044位于染色体A08上12.4 Mb处, 阈值为5.90, 单个位点解释的表型变异为 15.90%;S10_16428321位于染色体 A10上 16.4 Mb处, 阈值为5.62, 单个位点解释的表型变异为 15.64%。RIL群体中检测到的 QTL区间与关联分析检测到的显著关联SNP位点没有重叠。
2.5 株高、主花序长度、分枝区段长、一次分枝高度定位比较
对已经报道的株高相关性状定位结果进行整理,共找到160个株高位点, 36个主花序长度位点, 14个分枝区段长位点和 43个一次分枝高度位点。除了A04染色体外, 株高位点在其他染色体上都有分布,其中A03 (20个)染色体分布最多, A亚基因组每条染色体平均有11个, C亚基因组每条染色体平均有7个。对不同群体株高定位结果进行整合, 发现4个以上群体在 A01 (6.3~8.0 Mb)、A03 (6.3~7.9 Mb)、A07(16.6~19.4 Mb)、C03 (4.6~6.8 Mb)和 C06 (19.4~24.1 Mb)染色体上重复检测到株高位点(图 5)。主花序长度位点在A亚基因组上有17个, C亚基因组上有7个, 2个群体在A10染色体11.5~13.8 Mb处找到共同的主花序长度位点。分枝区段长位点位于A02、A04、A06、A07、A09、A10、C03、C04、C06、C07、C08和C09染色体上, 并未找到分枝区段长重复区间。一次分枝高度位点, 除了A05染色体外, 在其余染色体上都有分布, 2个群体在A02 (6.0~8.1 Mb)和C03染色体(4.8 Mb)上找到一次分枝高度的重叠区间。
对株高相关性状比较分析, 分别在 A01、A03、A05、A06、A07、A09、A10和C06染色体上检测到株高与主花序长度的共同位点, 在A01、A07、C03、C05和 C08染色体上检测到株高与一次分枝高度的共同位点, 在A02、A07和C06染色体上检测到株高与分枝区段长的共同位点, 在A07、A08染色体上检测到主花序长度和一次分枝高度的共同位点。另外,本研究在A07染色体17.9~22.0 Mb处找到株高、主花序长度、分枝区段长和一次分枝高度共同的位点。
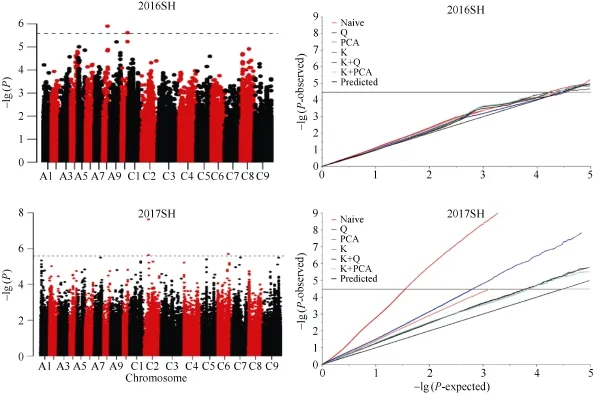
图4 甘蓝型油菜茎高全基因组关联分析Manhattan图和QQ图Fig. 4 Manhattan and quantile-quantile plot for stem height in 2016 and 2017 SH: 茎高。SH: stem height.
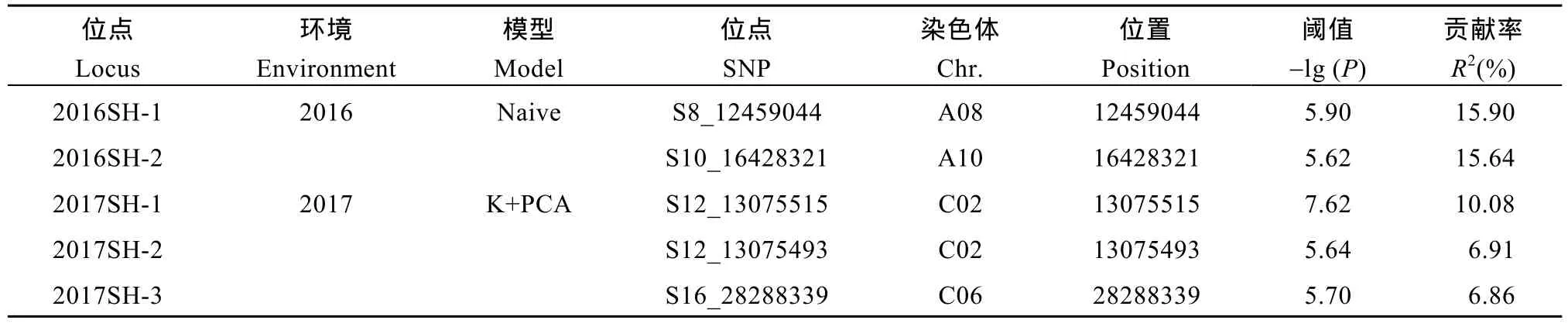
表4 甘蓝型油菜茎高显著关联位点Table 4 Markers with significant association for stem height
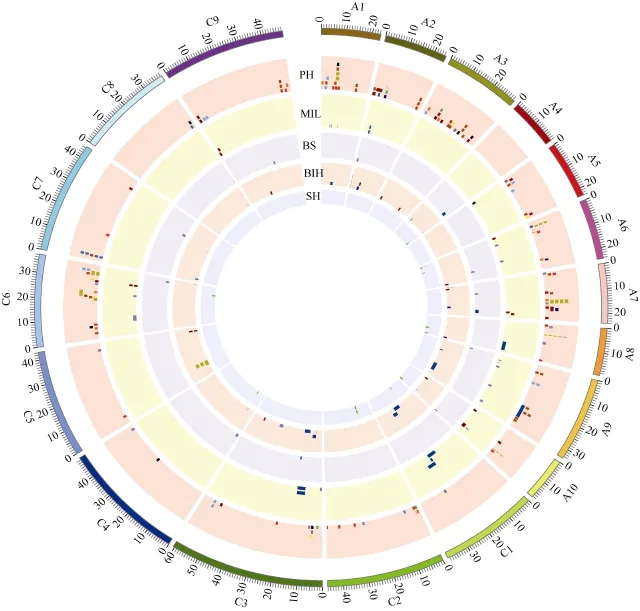
图5 不同群体株高相关性状位点整合。Fig. 5 Integration of the loci for plant height-related traits in B. napus
2.6 茎高候选基因鉴定
RIL群体和自然群体茎高定位区间与上述整合区间比较, 发现RIL群体定位位点q2016SH-A04-2、q2016SH-A04-3、q2016SH-C04、q2016SH-A06-1和q2017SH-A08, 自然群体定位位点 2016SH-1和2017SH-3与已报道的株高相关性状定位区间有重叠(图5)。根据茎高定位结果, 本研究在其附近寻找候选基因, 找到一些与激素途径(生长素、赤霉素和油菜素内酯)、光形态建成及植物生长发育相关的基因(表 5)。
3 讨论
株高是油菜重要的农艺性状之一, 其主要通过每株有效分枝数和角果数影响产量。株高与倒伏系数呈显著相关, 是影响倒伏的的一个重要因素, 倒伏不仅可以造成产量下降, 还可以使甘蓝型油菜含油量下降[28-29]。因此, 寻找控制株高的位点, 解析株高遗传机制, 筛选理想的株高品种是高产育种的关键[30]。主花序长度、分枝区段长和一次分枝高度是株高的重要组成部分, 现在已经有很多株高相关性状定位研究报道, 但由于使用的标记和群体不同,很难对其分析研究。本研究将株高相关性状定位位点整合到甘蓝型油菜物理图谱上, 可以准确和直观地比较不同群体的定位研究结果, 魏大勇等[31]通过整合甘蓝型油菜含油量位点, 在 A亚基因组染色体上(A01、A02、A03、A6、A08、A09和A10)找到7个整合区间。本研究中, 在4个以上群体A01、A03、C03和 C06染色体上都检测到控制株高的位点, 其可能是控制株高的稳定位点。另外, 在 A10染色体上找到一个主花序长度重叠区间, 在A02和C03染色上找到一次分枝高度重叠区间, 但未找到分枝区段长重叠区间, 可能是由于目前分枝区段长相关研究报道较少。本研究还找到株高与主花序长度、株高与一次分枝高度、株高与分枝区度长、株高与茎高定位的重叠区间, 表明株高组成性状对株高具有重要作用。在A07染色体17.9~22.0 Mb处, 检测到株高、主花序长度、分枝区段长和一次分枝高度共定位, 表明该位点具有一因多效。
茎高是株高性状的重要组成部分, 本研究结果表明茎高与株高呈显著相关, 对经济产量起着重要的作用, Sabaghnia等[3]也证明茎高对经济产量的效应最大, 因此研究控制茎高的位点是重要的。本研究通过 QTL定位和全基因组关联分析, 在 A04、A06、A08、A10、C01、C02、C04和 C06染色体上找到与茎高相关的位点。本文构建的遗传图谱大小为 6140.2 cM, 与其他油菜连锁群差异较大, 其可能原因是偏分离标记数目较多, N’Diaye等[32]研究表明, 当利用 10%的共分离标记时, 小麦遗传图谱长度增加 4.5%; 当利用 80%的共分离标记时, 遗传图谱长度增加 16%。虽然共分离标记可以增加图谱长度, 但是对标记顺序没有影响。除了共分离标记外,定位群体大小和类型、标记数目、定位软件、重组事件都会对遗传图谱大小有影响[33-36]。本研究中RIL群体定位到的 QTL解释表型变异大于 100%, 可能是由于一些QTL位点存在连锁, 当QTL独立遗传时,多个QTL的贡献率等于单个QTL的贡献率之和; 当QTL连锁时, 贡献率不能简单相加[37]。另外, RIL群体茎高主效QTL 2年未被重复检测到, 可能是由于2年环境差异较大, 2016年重庆气温偏高, 而 2017年气温偏低, 暴雨偏多。QTL与环境互作是影响数量性状表达的重要因素, 大多数 QTL受环境影响,张江江等[18]研究表明环境对株高 QTL表达有很大影响。
本研究在RIL群体中检测到的QTL区间与关联分析检测到的显著关联SNP位点也没有重叠, 可能是由于 2种方法利用的定位群体不一样, 检测到的茎高位点不相同。RIL作图群体为两亲本杂交重组后代, QTL定位受亲本影响; 而关联分析利用了自然群体中长期积累的重组和变异。虽然本研究中未找到共同定位的区间, 但是与上述整合区间比较发现, 与前人报道的株高相关性状定位区间有重叠。另外, 本研究在其附近找到一些茎高候选基因, 为甘蓝型油菜株型育种提供了理论基础。
4 结论
茎高可以使株高、结荚高度和分枝起点提高,却不能提高结荚厚度, 在生产实践中应适度控制茎高。在 A04、A06、A08、A10、C01、C02、C04 和C06染色体上找到与茎高相关的位点及一些候选基因。整合株高相关性状位点, 找到遗传稳定的株高、主花序长度和一次分枝高度区间。
猜你喜欢
果树实用技术与信息(2022年6期)2022-11-21
作物学报(2022年12期)2022-10-14
作物学报(2022年4期)2022-02-10
作物学报(2022年3期)2022-01-22
语文周报·教研版(2021年28期)2021-08-19
农村科学实验(2021年2期)2021-04-03
中国农业科学(2020年16期)2020-09-11
农村百事通(2019年17期)2019-10-08
陕西农业科学(2019年4期)2019-05-13
现代农业科技(2017年1期)2017-03-06
