
3′-碘甲状腺原氨酸:保护心脏和调节代谢的新武器
2019-05-29 10:05:34郑辉常宝成葛焕琦
国际内分泌代谢杂志 2019年2期
郑辉 常宝成 葛焕琦
1泰达国际心血管病医院内分泌科,天津300457;2国家卫生健康委员会激素与发育重点实验室(天津医科大学),天津市代谢性疾病重点实验室,天津医科大学代谢病医院内分泌研究所300070
T4是甲状腺激素的主要形式。在靶组织内,T4被脱碘酶脱碘转化为T3。T3在组织内通过其受体调节下游蛋白的转录是一个缓慢的过程,需要数小时甚至数天。2004年Scanlan在人和啮齿类动物体内发现一种新的天然甲状腺激素衍生物,能够在数分钟内降低体温、减慢心率,减少心脏输出。这一衍生物被称为3′-碘甲状腺原氨酸(3′-Iodothyronamine,T1AM),它是G蛋白耦联的微量胺受体(TAAR)1的激动剂,刺激产生快速的生理和行为调节,有些作用是与T3相对抗的。近年来的研究发现,T1AM与心血管系统和代谢调节密切相关。目前进行的T1AM临床研究有限,更多的是进行动物实验和体外研究。本文将就T1AM对心血管保护和代谢调节的最新研究进展做一介绍。
1 T1AM的结构与代谢过程
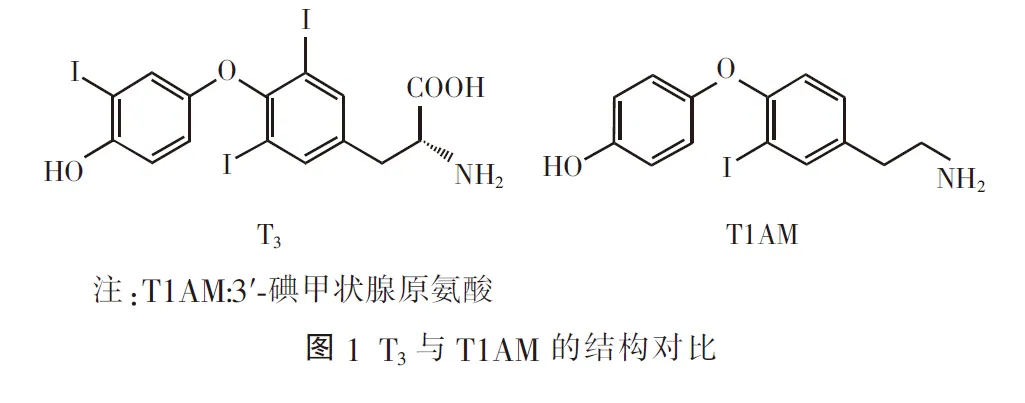
图1是T1AM的结构图。T1AM合成的生物学过程还不清楚。由于T1AM与T3结构相似,它可能是由T3脱碘、脱羧生成。Hackenmueller等[1]应用13C9-15N-T4[重-T4(H-T4)]技术测定肝脏中T1AM,发现甲状腺过氧化物酶抑制剂能够抑制T1AM的合成。对于T1AM合成过程有3种推测:一是苯酚与单碘酪氨酸的耦联,是在T4分泌过程中产生甲状腺球蛋白水解的副产物。二是由甲状腺直接合成并分泌。最后,T1AM可能是甲状腺产生的其他物质的代谢产物或者是特异性产生在甲状腺内的T4代谢产物。总之,T1AM合成过程还需要进一步的研究。
组织中T1AM的代谢过程是经单胺氧化酶氧化脱氨生成3碘乙酸,再经过脱碘酶脱碘生成乙酸,最后被硫酸盐和葡糖醛酸盐进行N乙酰化和酯化。T1AM及其代谢产物由胆道和肾脏排出。T1AM在体内、外迅速被单胺氧化酶代谢产生乙酸。
2 人血清T1AM的测定
人血清中T1AM的检测一度陷入困境。Galli等[2]应用高效液相色谱-串联质谱法定量分析人血清和组织中的T1AM浓度平均为0.219 nmol/L。但该方法不能评估与蛋白相结合的T1AM,不能区分游离和结合的T1AM。Hoefig等[3]应用化学发光免疫分析法,测定人血清中的T1AM浓度为66 nmol/L,但是该方法测定可能高估T1AM水平,因为在血清中T1AM与载脂蛋白B-100结合;另外也对T1AM衍生物产生交叉反应,比如硫化的T1AM和其他被修饰的T1AM。
3 T1AM的组织分布
在啮齿类动物体内包括大脑、心脏、血液以及人类的血液中均能监测到T1AM。应用高效液相色谱法测定,在大鼠组织中的浓度为1~90 pmol/g,在肝脏、大脑和肌肉中的含量最高,血清中的浓度在0.2~0.3 nmol/L,可见组织中的浓度显著高于血清中的浓度,表明组织特异性的摄取T1AM。在血清中,T1AM有90%与载脂蛋白 B-100结合[4]。
4 T1AM作用的受体
T1AM结合的受体是多源性的。大致分为G蛋白耦联受体和非G蛋白耦联受体。G蛋白耦联受体主要包括TAAR、α2A肾上腺素能受体和β2肾上腺素能受体。而非G蛋白耦联受体主要包括瞬时感受电位通道(TRPs)。
TAAR是2001年首先报道的,共分为9种不同的亚型,在多种器官中广泛表达,包括大脑、心脏、肝脏、肾脏、胰腺和脾脏。其中,TAAR1与T1AM的亲和力最强。TAAR1与GαS蛋白耦联激活腺苷酸环化酶,增加细胞内cAMP的生成,促进内向钾离子通道开放。除了TAAR1,T1AM还能作用于TAAR5[5],抑制三磷酸肌醇形成和丝裂原活化蛋白激酶(MAPK)信号通路。
T1AM与α2A肾上腺素能受体结合能调节胰岛素的分泌,激活GI/o,减少去甲肾上腺素诱导的MAPK的活性[6]。
T1AM能够直接作用于HEK293细胞的β2肾上腺素能受体,浓度依赖性的激活Gs信号通路[5]。β2肾上腺素能受体阻滞剂timolol能够阻断T1AM诱导的Ca2+内流。
T1AM还能与非G蛋白耦联受体的TRPs作用。TRPs是超家族,包括28个成员,T1AM主要与TRPM8相结合作用于Ca2+,进而影响体温[7]。
5 T1AM的生理功能
5.1 T1AM对心脏的作用 T1AM能够通过减少氧化应激起到保护心脏的作用。Ghanian等[8]发现,高剂量的T1AM(25 mg/kg)能降低小鼠心脏氧化应激水平,而在肾脏却增加氧化应激水平。表明了T1AM作用的器官特异性,可能在不同器官中作用于不同的信号通路。
T1AM能够改善心肌供血。T1AM作用于蛋白激酶C/KATP信号通路,可减少分离的大鼠心肌缺血-再灌注模型的梗死面积,起到保护心肌的作用。
T1AM可调节心肌细胞离子稳态。它能减少内质网钙离子释放,抑制钾离子电流。尤其是细胞外流It0和Ik1背景流。20~25 μmol/L的T1AM能够降低心肌收缩力,但氧气消耗和葡萄糖摄取没有变化。
T1AM呈现剂量依赖性的负性变力、负性变时效应。Chiellini等[9]发现,离体工作的大鼠心脏随着给与T1AM浓度的增加,心脏输出量和心率也逐渐下降。la Cour等[10]观察心力衰竭患者T1AM水平显著升高,与心脏射血分数呈负相关。
最新的研究表明,T1AM 能通过作用于中枢系统间接引起血管舒张,进而影响心血管系统[11]。该研究阐明,虽然在雄性鼠的尾动脉和主动脉中可能存在T1AM的作用靶点,但肌电图研究未显示T1AM对血管舒张有任何直接作用,提示其作用可能是间接的。然而,脑室内应用T1AM出现了尾动脉的扩张和体温下降,并导致下丘脑神经元激活,表明T1AM或其代谢产物通过下丘脑信号途径引起尾部血管扩张。
5.2 T1AM对体重的作用 肥胖是心血管疾病和代谢性疾病的危险因素。在大鼠研究中,给予T1AM 10 mg/(kg·d),持续7 d,净重下降6%,给予25 mg/(kg·d),净重下降14%,但不影响食欲及生活,能使碳水化合物转化为脂肪代谢[12]。
T1AM能直接作用于中枢系统,控制摄食行为。T1AM受体在中枢神经系统中有广泛的分布。在大脑主要是TAAR,其在单胺能系统的主要神经核中含量丰富。脑内注射T1AM能显著改善摄食行为,促进食欲,增加摄食量, Manni等[13]发现T1AM改变食欲呈双向,低剂量(3.3 nmol/kg)导致厌食,高剂量(51 nmol/kg)导致食欲旺盛,而大剂量(79 nmol/kg)长时间(14 d)治疗却减少食物的摄取。Haviland等[12]观察持续8 d给予T1AM产生的慢性代谢影响,发现注射T1AM后脂肪利用增加,治疗8 d,体重下降8.2%,而且减重效果持久,在停止治疗2周后,体重反弹仅有下降体重的1.8%。而这些慢性的作用可能与基因表达的调节有关。T1AM改变摄食行为的机制还不清楚,首先可能与T1AM在体外诱导下丘脑释放神经肽Y有关。其次,组胺能神经元形成一个网络,平衡神经内分泌和下丘脑食物摄取,T1AM可能通过这种双重作用对食欲及摄食行为产生影响。
5.3 T1AM对代谢的作用
5.3.1 T1AM对脂代谢的影响 Braulke等[14]在啮齿类动物实验中发现,腹腔内注射T1AM后的急性代谢反应是降低呼吸商,表明代谢途径由碳水化合物为主转变为脂质氧化为主,引起酮尿和体内脂肪含量的显著下降。同时,T1AM通过降低脂代谢关键调节基因PTP1B和PLIN2的表达,减少肝脏胆固醇和甘油三酯的生成,显著增加代谢产物(糖异生、氨基酸、肌酐)[15]。T1AM能够使肝脏PTP1B编码的胰岛素信号通路蛋白表达下降,使肌肉组织中胰岛素受体底物2表达上调,使脂肪组织中葡萄糖转运蛋白2表达增加,并使GYS1(编码糖原合成酶)的基因下调,表明代谢从碳水化合物转向脂代谢。
在动物模型中,T1AM可迅速改变能量来源,从碳水化合物转到脂肪氧化,延缓蛋白质的降解。T1AM是F(0)/F(1)-ATP酶的非竞争抑制剂,体内低剂量T1AM作用于F(0)/F(1)-ATP酶的高亲和力的结合位点,占据F1的天然抑制蛋白IF1结合位点,促进IF1解离,增加ADP刺激的线粒体呼吸,抑制线粒体能量产生[16]。
Assadi-Porter等[17]发现,T1AM能引起肝脏中Sirt6基因(一个葡萄糖调节基因)的表达和葡萄糖激酶的表达,抑制Sirt4基因(该基因抑制脂肪氧化)表达。T1AM能够调节脂肪组织中350个基因和肝脏的100余个基因,这些基因表达的变化刺激脂肪分解和β氧化,抑制脂肪合成[18]。
5.3.2 T1AM对糖代谢的影响 一项小样本的临床研究显示,糖尿病患者血清T1AM浓度显著增加,且与糖化血红蛋白和空腹血糖呈正相关。T1AM水平升高的患者血糖水平更高,胰岛素分泌受损[2]。Klieverik等[19]给予大鼠静脉(50 mg/kg)和脑内注射(0.5 mg/kg)T1AM,均能迅速(10 min开始)增加内源性葡萄糖产物和血浆葡萄糖水平。
T1AM通过影响糖代谢调节激素,引起血浆葡萄糖升高。研究发现,无论静脉和颅内注射T1AM均能够引起胰高血糖素和皮质酮水平的升高,作用持久,而对血浆胰岛素的影响仅在注射后10 min产生抑制作用,注射120 min后胰岛素水平恢复到注射前水平。
近年来更多的研究证实,T1AM能够抑制胰岛素的释放。T1AM能够直接作用于α和β细胞的G蛋白耦联受体,其对胰岛素分泌是双向调节的,其作用于TAAR1可刺激胰岛素分泌,作用于α2A肾上腺素能受体则抑制胰岛素分泌。在生理状态下,T1AM主要通过α2A肾上腺素能受体抑制胰岛素分泌。Lehmphul等[20]体外研究结果显示,T1AM通过与胰岛细胞细胞膜的甲状腺激素跨膜转运子结合,进入到细胞内,被细胞内的单胺氧化酶降解为3-碘甲酰乙酸,T1AM和3-碘甲酰乙酸均能抑制线粒体F0/F1-ATPase,减少ATP的生成和葡萄糖依赖的胰岛素分泌。
T1AM还能调节糖异生途径。Ghelardoni等[21]发现,0.5~1 μmol/L T1AM 能够增加肝脏酮体生成,刺激HepG2细胞的糖异生。
5.3.3 T1AM对蛋白质代谢的影响 T1AM长期作用的结果能够促进蛋白质的分解代谢。T1AM治疗 5~7 d的小鼠,出现缬氨酸、甘氨酸等蛋白质代谢产物的增加。机体由碳水化合物的氧化转向蛋白质的氧化。因此,高剂量T1AM能够减少肌肉张力和肌肉收缩的强度。迄今尚未报道短期T1AM治疗对蛋白质分解代谢的影响。通过激活FoxO-蛋白酶体信号及下调蛋白激酶B1-S6K信号,T1AM抑制代谢,诱导肌细胞肥大[22]。T1AM抑制代谢的作用主要通过激活蛋白质分解代谢信号、抑制合成代谢信号,进而诱导肌细胞萎缩,抑制多种代谢相关基因的磷酸化。
5.3.4 T1AM对内分泌激素的影响 T1AM能作用于大鼠下丘脑-垂体-肾上腺轴,调节包括胰岛素、糖皮质激素和甲状腺激素。观察T1AM在大鼠体内调节下丘脑-胰腺-甲状腺轴,发现较低剂量(1.3 μg/kg)的T1AM能产生低血容量效应,减少下丘脑c-fos的活性,增加血浆血糖,减少胰岛素敏感性,降低血浆FT3水平。
T1AM抑制心脏的作用与甲状腺激素作用相反,而它的代谢效应却与甲状腺激素起到协同作用,均能促进脂肪分解。
6 总结
大自然中的生命现象都是互相制衡,共同存在的。新近成为研究热点的甲状腺激素衍生物T1AM正是体内拮抗甲状腺激素生理作用的物质。深入研究其作用信号通路和更多的生理学作用,可为包括治疗心力衰竭、肥胖、糖尿病等提供新的治疗方向。
猜你喜欢
青少年科技博览(中学版)(2022年11期)2023-01-07 06:21:30
保健医苑(2022年4期)2022-05-05 06:11:08
猪业科学(2022年2期)2022-04-21 02:18:26
汽车维修与保养(2021年8期)2021-02-16 00:28:20
中国生殖健康(2020年2期)2021-01-18 02:51:14
中国生殖健康(2019年8期)2019-01-07 01:18:00
工业设计(2016年4期)2016-05-04 04:00:15
物理化学学报(2015年7期)2015-12-30 12:13:04
河南科技(2015年2期)2015-02-27 14:20:35
振动、测试与诊断(2014年4期)2014-03-01 01:14:07
