
高原鼠兔洞穴密度与高寒草甸植物群落结构以及土壤因子的关系
2019-05-28 06:35程云湘周国利马青青常生华
草业科学 2019年4期
田 富,程云湘,2,周国利,马青青,申 波,曲 久,常生华
(1.兰州大学草地农业生态系统国家重点实验室 / 兰州大学农业农村部草牧业创新重点实验室 / 兰州大学草地农业教育部工程研究中心 / 兰州大学草地农业科技学院,甘肃 兰州 730020;2.内蒙古大学生态与环境学院,内蒙古 呼和浩特 010021;3.西藏那曲市畜牧兽医技术推广总站,西藏 那曲 852000)
高寒草甸是在高原和高山寒冷环境条件下形成的一种草地类型,相对于其他草地类型,高寒草甸在我国青藏高原分布面积最大,可利用面积达5.88万hm2,高寒草甸具有多重功能与效益,除为农牧民提供基本的生产资料外,还为青藏高原提供重要的生态保障[1-2]。由于所处的特殊地理位置,其生态环境较为脆弱,目前我国近90%的高寒草甸都有不同程度的退化,局部地区退化严重,导致高寒草甸退化的原因包括气候变迁、过度放牧以及不合理的草地管理制度等,其中人为干扰以及对高寒草甸生态功能重要性的忽视是导致高寒草甸退化的最主要原因[3],高原鼠兔(Ochotona curzoniae)的频繁活动也加速了高寒草甸的退化[4],高山嵩草草甸因植株低矮常成为高原鼠兔的理想栖息地,研究高原鼠兔对高寒草甸土壤及植被的影响对于保护高寒草甸生境、揭示其群落演替具有重要意义。
高原鼠兔是一种小型非冬眠的植食性哺乳动物,鼠兔挖掘洞穴及排泄行为可为植被生长提供养分,其洞穴不仅可为自身栖息及抵御外界的不良环境提供庇护,同时也是其他鸟类以及小型穴居动物的栖息场所,高原鼠兔作为青藏高原生境重要的物种之一有着不可替代的作用[5-6],但近年来鼠兔种群密度急剧增多[7-9],对植被的采食破坏了当地的生态环境,对当地植被多样性及畜牧业经济发展产生了严重影响。高原鼠兔在高寒草甸生态系统中的作用随种群密度的变化而发生改变,适宜的高原鼠兔干扰可以改善土壤养分结构[10-11],提高土壤涵养水源能力[12],鼠兔的采食及挖掘活动有利于传播和扩散植物种子[13],提高高寒草甸植被多样性,增加可食牧草比例[14],但当高原鼠兔干扰过高时则会降低土壤养分[10-11],涵养水源能力下降[15],植被群落结构不均匀性增加,不可食牧草比例增多[14],可见高寒草甸生境状况会随高原鼠兔种群密度变化而发生适应性改变。
为保护青藏高原生态环境,缓解鼠害问题,有必要对高原鼠兔种群密度产生影响的土壤及植被做相关的研究。目前对于高原鼠兔与高寒草甸植被群落与土壤的关系研究集中在比较高原鼠兔干扰与非干扰区[16-17]、有无鼠丘区[18-19]以及不同鼠丘堆积年限之间的差异[20],但对于高原鼠兔在不同干扰水平下与植被群落结构与土壤因子的关系研究较少。本研究以青海省果洛州玛沁县为调查地,以青藏高原高寒草甸为研究对象,分析高原鼠兔有效鼠洞与斑块状裸地、土壤物理特征、高寒草甸植被特征间的关联性,以期对高原鼠兔影响青藏高原高寒草甸的演变提供一定的参考依据,为合理区划鼠害界限及促进三江源区草畜平衡提供数据支持。
1 研究地区概况与方法
1.1 研究地区概况
果洛藏族自治州位于青海省东南部(97°54′-101°50′ E,32°31′-35°40′ N),面积 76 312 km2。气候属典型高原大陆性气候,年均气温为-4 ℃,无绝对无霜期。年降水量为400~760 mm,80%~90%的降水集中在5-9月。果洛草地以高寒草甸类为主,藏嵩草(Kobresia tibetica)、矮嵩草(K.humilis)、高山嵩草(K.pygmaea)、紫花针茅(Stipa purpurea)为常见优势种,植被盖度在80%~100%。当地鼠害严重,暴发频繁,受当地宗教习俗的影响,鼠兔防治主要依靠外来人员,成效微弱。
1.2 研究方法
1.2.1 试验设计
受多种因素的影响,直接观测单位面积内高原鼠兔数量具有不确定性[21]。啮齿动物的有效洞穴数目与高原诸多环境因子都具有很好的相关性[22],本研究借鉴了先前的研究[21-22],采用有效鼠洞数目来指示高原鼠兔密度。
研究区设在果洛州玛沁县大武镇格姆滩,选取以自由放牧利用下的草地为研究对象,草地类型以高寒草甸为主。在2017年6月共设置27个样地,取10 m × 10 m 的鼠洞采样范围,在该范围内采用连续3 d堵洞法统计有效鼠洞数量,以第1天统计的鼠洞数记为总洞口数后填埋,后3 d连续每天在15:00-16:00调查记录被鼠兔盗开的新洞口并计数,然后重新填埋。每天被鼠兔盗开的新洞口即为当天的有效洞口,最后确定的有效洞口即为连续3 d的平均值[23]。同时调查各样地内的裸斑面积、植被盖度及优势种植物。根据有效洞口数、斑块状裸地总面积和植被总盖度等,从27个样方聚类中选择立地条件相似的样地,共设置4个鼠洞密度梯度的样地类群 (Ⅰ、Ⅱ、Ⅲ、Ⅳ),其有效鼠洞密度每100 m2依次为(4 ± 3)、(15 ± 2)、(24 ± 2)、(37 ± 5)个。
1.2.2 测定指标及方法
在每个梯度样地内按“N”形分布设置4个0.5 m ×0.5 m的样方,共16个。清查和统计出现在样方中的所有植物种,测定植株个体的种类、高度、盖度等,辨认可食与不可食植物类群,然后分种采集植物地上生物量[24]。利用土壤紧实度测定仪(SC900)测定样方内土壤紧实度,并按对角线打土钻3次,按照0-10 cm、10-20 cm分层收集,装入密封袋中带回实验室分析。用直径为5 cm的土钻沿样地的对角线等距离取样,3次重复,称湿重后在105 ℃下烘干48 h至恒重,称干重,计算表层(0-10 cm)土壤含水量。
1.3 数据处理与分析
1.3.1 土壤含水量
土壤含水量一般是指土壤的绝对含水量,即在单位重量内土壤所含有的绝对水分[25]。
计算公式:W= (W2-W3)/(W3-W1) × 100%。式中:W为含水率(%);W1为称皿重(g);W2为称皿 + 风干土重 (g);W3为称皿 + 烘干土重 (g)。
1.3.2 重要值
重要值是计算物种多样性指数的重要指标[26],本研究采用相对高度和相对频度计算物种相对重要值。
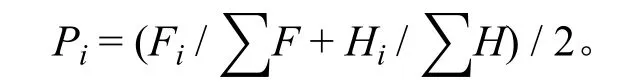
式中:Pi为0.25 m2样方中第i种的重要值;Fi表示i种在单个样方中出现的次数,Hi表示i种的平均高度;F、H分别表示样方中物种的出现的总次数、总平均高度高。
1.3.3 植物多样性:
采用Pielou指数、Shannon-Wiener指数和Simpson指数分别表征均匀度指数、多样性指数和优势度指数[12],计算公式: ∑
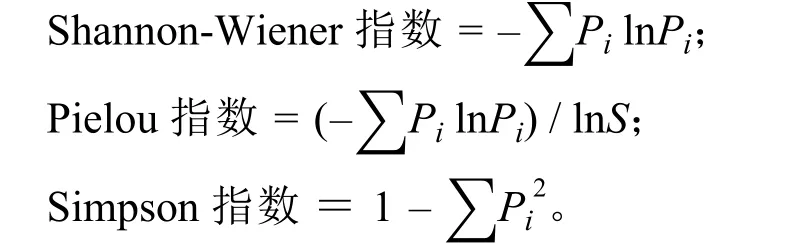
式中:S为 0.25 m2样方中的物种数,Pi为 0.25 m2样方中第i种的重要值。
1.3.4 软件统计分析
用SPSS 20.0统计分析软件单因素方差分析比较各处理间的差异显著性。
2 结果与分析
2.1 鼠洞密度对高寒草甸土壤物理性质的影响
鼠洞密度显著降低了高寒草甸表层土壤含水量(P< 0.05)(图1),梯度Ⅰ显著高于其他梯度。在Ⅱ、Ⅲ、Ⅳ样地随鼠兔密度的增加土壤含水量基本维持在50%以下,且梯度Ⅱ、Ⅲ、Ⅳ之间差异不显著(P> 0.05);鼠洞密度对高寒草甸土壤紧实度的影响微弱,除梯度Ⅰ显著低于梯度Ⅱ样地外,其他梯度两两之间的土壤紧实度差异不显著(P> 0.05)(图2)。
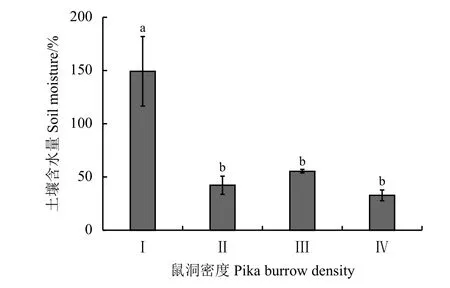
图1 不同鼠洞密度梯度下高寒草甸的土壤含水量Figure 1 Soil moisture at different pika burrow densities
2.2 鼠洞密度对样地裸地面积的影响
鼠洞密度显著影响样地内斑块状裸地面积(P<0.05)。梯度Ⅱ、Ⅲ、Ⅳ时斑块状裸地总面积依次上升,分别为 3、7、8.75 m2(图 3)。
2.3 鼠洞密度对植物群落结构的影响
2.3.1 鼠洞密度对群落盖度的影响
鼠洞密度对高寒草甸群落盖度有显著影响(P<0.05)(图4)。其中,梯度Ⅰ群落盖度显著高于其他梯度,随鼠洞密度的增加,群落盖度逐渐降低,但在Ⅱ、Ⅲ、Ⅳ之间植物群落盖度差异不显著(P> 0.05)。
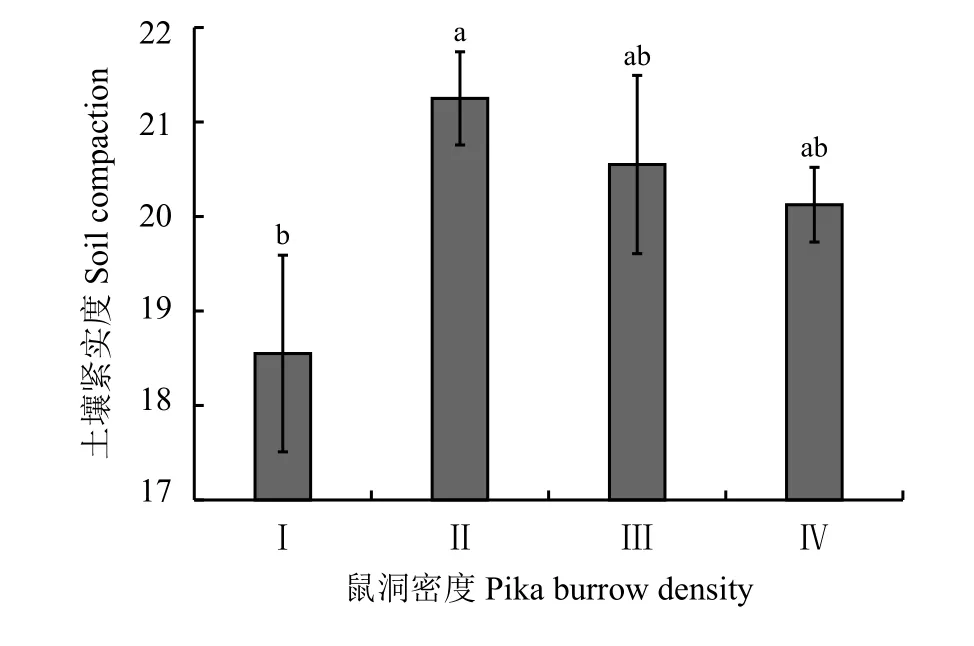
图2 不同鼠洞密度梯度下高寒草甸的土壤紧实度Figure 2 Soil compaction at different pika burrow densities
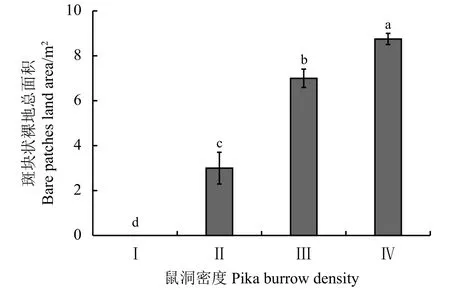
图3 不同鼠洞密度梯度下高寒草甸的斑块状裸地总面积Figure 3 Total area of bare patches at different pika burrow densities
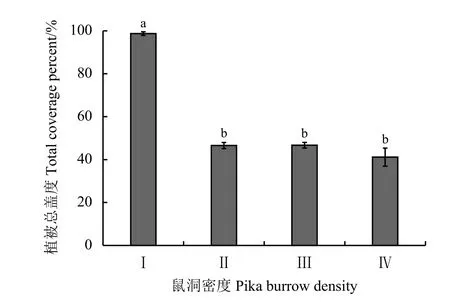
图4 不同鼠洞密度梯度下高寒草甸的植被总盖度Figure 4 Total coverage percent at different pika burrow densities
2.3.2 鼠洞密度对群落物种组成及其重要值的影响
伴随鼠洞梯度的逐步增加,植物群落物种组成总数逐步减少(表1)。其中,梯度Ⅰ时物种数量最多,梯度Ⅳ时物种数量最少。优势种高山嵩草、矮嵩草的重要值呈现先增加后降低的趋势,但其重要值仍为最大数值,说明其优势种地位并没有发生改变;伴生种秦艽、垂穗披碱草、毛果婆婆纳的重要值伴随鼠洞梯度的增加逐步增大,说明其在植物群落内的作用逐步增强;矮泽芹、西藏堇菜、矮金莲伴随鼠洞梯度的增加逐步退出高寒草甸植被群落,其在植被群落中的作用完全消失;梯度Ⅰ下高寒草甸植被群落物种组成丰富,少见种甘肃嵩草(Kobresia kansuensis)、花葶驴蹄草(Caltha scaposa)、南山龙胆(Gentiana grumii)的重要值只在梯度Ⅰ出现。高寒草甸植物群落物种组成数量伴随鼠洞数梯度的增高而呈现逐步较少的趋势,梯度Ⅰ种类最多,梯度Ⅳ种类最少,梯度Ⅱ、Ⅲ间植被群落物种组成数大致相同,仅个别物种类型有所误差,二者间植物群落物种组成结构差异不大。
2.3.3 鼠洞密度对高寒草甸群落多样性的影响
不同鼠洞密度下植物多样性指数及丰富度指数差异不显著(P> 0.05),随鼠洞密度增加,植物多样性指数和丰富度指数呈现出相同的变化趋势,梯度Ⅰ、Ⅱ下高于梯度Ⅲ、Ⅳ(表2);鼠洞密度对均匀度指数影响显著(P< 0.05),且数值随不同鼠洞梯度的逐渐变大呈逐步增加的趋势。在各项指标中,梯度Ⅱ内的植被数据相对于其他3个梯度数值较高,说明梯度Ⅱ内物种多样性指数丰富,鼠类活动干扰影响最为微弱。

表2 不同鼠洞梯度下高寒草甸植物群落的物种多样性指数分析Table 2 species diversity index analysis of alpine meadow flora under different pika burrow density gradients
3 讨论
3.1 鼠洞梯度与高寒草甸土壤物理性质的关系
高原鼠兔栖息地的选择受植被、土壤、地形、气候、天敌等诸多生物及非生物因素的影响,鼠兔洞穴的建造最直接相关的即为土壤因素[27]。本研究分析了土壤含水量与土壤紧实度和鼠兔洞穴的关系,结果发现,土壤含水量对鼠兔洞穴的建造影响占比较多,以土壤0-10 cm地下含水量低于50%为界限,鼠兔洞穴的密度较大,说明高原鼠兔对于其栖息地环境具有选择性行为,相对于其他地区,洞穴建造位置的选取可能与果洛地区降水量有关,高原鼠兔洞穴的建造倾向于坡度较缓的坡地[25, 28-29],且从河滩平地向坡地逐步进行了转移,鼠兔倾向于向坡度较缓的坡地聚集的选择行为可能与坡地利于排水、保持洞穴干燥有关。当鼠兔洞穴数目急剧下降甚至消失时,地下含水量通常较高且其生境高原鼠兔种群密度不具备过高的风险。土壤紧实度对鼠兔洞穴建造位置的选取并无明显的相关性,这可能与果洛地区高海拔地势以及气候有关,土壤间紧实度的变化不大从而影响效果微弱。
3.2 鼠洞密度与裸地面积的关系
斑块状裸地是生态系统的景观特征,斑块状裸地面积的集聚导致“黑土滩”现象的出现,形成退化的高寒草甸:土壤养分降低,草地植被退化严重,植被更新周期延长,生境恢复难度加大。根据以往的研究,对于斑块状裸地未有完全规范的定义,“黑土滩”是目前较为认可的一种称呼,它的形成是多种因素共同作用形成的结果[30],其中高原鼠兔挖掘洞穴加速了“黑土滩”的形成,在严重退化的黑土滩草地上,鼠洞密布且鼠类活动猖獗[31-32]。本研究主要探讨高原鼠兔干扰与斑块状裸地面积间的互作关系,当地鼠害严重,样地中“黑土滩”存在的形式多为斑块状裸地,呈秃斑块状存在,且单个面积有不断扩大的趋势。林慧龙等[33]在江河源区“黑土滩”退化草地秃斑与鼠洞的分形特征中指出,鼠洞总面积和鼠洞破碎指数呈反比关系,鼠洞形状和退化程度相关,秃斑总面积与秃斑破碎指数并不严格呈反比关系,秃斑形状与退化程度相关,随退化程度加重而逐渐增多,秃斑多样性指数越高。由于秃斑多样性与研究样地大小及当地“黑土滩”退化程度等相关,本研究结果指出,斑块状裸地总面积与鼠洞梯度大致呈正比关系,伴随鼠洞梯度增高,裸斑地总面积也急剧增加,说明退化严重的地区更易导致鼠类的迁入并且长期生存,同时鼠类的聚集栖息也加剧了该地区生态环境的退化,高寒草甸秃斑化和高原鼠兔生境迁入二者存在相互影响的关系,这与江小蕾[34]的研究结论相一致。与以往研究不同,这里需要指出在样地面积一定的前提下,斑块状裸地总面积增长到一定比例(8.75%)时达到饱和状态,此时鼠洞数目保持相对稳定且不在增长。
不同斑块面积形状的秃斑地对土壤侵蚀的意义还有待研究,但可以确定的是鼠类活动与土壤侵蚀间存在着某种必然的联系,斑块状裸地数目与鼠类栖息活动也相互关联,近年来高原鼠兔种群密度的增加是高寒草甸生态环境退化、斑块状裸地面积增多的伴生物。
3.3 鼠洞密度与高寒草甸植被群落的关系分析
3.3.1 鼠洞密度与群落盖度的关系
青藏高原植被主要以低矮草原群落为主,在开阔的生境中并没有高大的树木为原生动物提供庇护场所,众多动物的栖息场所主要以洞穴为主。高原鼠兔是青藏高原的一个关键物种,种群数量庞大,栖息地范围广泛[35],它的洞穴为诸多动物提供了栖息场所,褐翅雪雀、棕背雪雀、黑喉雪雀、棕颈雪雀等的巢穴都建立在高原鼠兔洞穴的基础上,这些鸟类在鼠兔居住的地区更为丰富[36]。此外,其他穴居动物也使用鼠兔洞穴作为繁殖和栖息场所。高原鼠兔的消失,以及由此产生的洞穴问题,对许多物种产生了负面影响,并会减少青藏高原上的原生生物多样性[37]。
由于高原生物群落结构单一,隐蔽条件差,又因高海拔气候的影响,洞穴栖息对其繁衍生息至关重要,它们的栖息环境除非为不可抗力突变,并不会随意迁移,而以长期定居为主。研究发现,鼠兔洞穴建筑位置的选择倾向于周边植株低矮,覆盖率低的环境,鼠兔会不定期对洞穴周边植被进行刈割以保持开阔的视野,降低被天敌捕食以及周边环境的威胁[38-39]。果洛地区降水充沛,高寒草甸群落覆盖率较高,当覆盖率低于50%时,鼠兔洞穴数目逐步增大,相反,当覆盖率过高时,则不适宜鼠兔筑巢栖息,过高的植被群落盖度会影响高原鼠兔的视野,增加采食的风险,缩小高原鼠兔的活动范围。高原鼠兔对于高寒草甸植被盖度的影响与斑块状裸地数量的增加也存在因果关系,鼠兔种群密度增多,其采食及刈割植被行为导致植被覆盖率下降,物种多样性指数减少,进而导致斑块状裸地面积的增加,植被盖度下降与裸地面积增多是鼠兔干扰增多的相关关联性表现。
高原鼠兔栖息地的选择受多种因素的影响,在诸多影响因素中,由于高寒草甸条件直接决定着鼠兔食物的数量和品质,同时也在很大程度上影响着其他环境因素的状况[40],因此,高寒草甸群落状况在鼠兔栖息地选择的影响因素中占有重要的地位。青藏高原上以鼠兔为代表的穴居动物引起的地表扰动会影响群落物种的多样性,鼠兔挖掘鼠洞的行为会对群落结构形成干扰,研究表明,高寒草甸群落高度和盖度显著影响鼠兔种群结构,但群落多样性与鼠兔群落结构表现为非线性相关,鼠兔采食喜食植被后,植被群落盖度与物种多样性指数数值变化不大,说明受鼠兔栖息环境的影响,当地的原优势植被仍占主导地位,伴随鼠洞密度梯度的增加,个别植被物种逐步退出了物种群落[41]。
3.3.2 鼠洞梯度与植被群落物种组成及结构的关系
对高寒草甸植被群落结构影响最直接的即为高原鼠兔对植被生物量的采食,鼠兔对各种植被的选择性采食主要受其口感喜好以及生境条件限制的影响。高原鼠兔属于白昼型活动的种类,活动范围一般仅限于洞穴周边地区,以各种牧草为食物,主要取食禾本科、莎草科以及豆科植物,而对嵩草属的植被采食率较低[42]。研究表明,伴随高原鼠兔干扰活动的逐步增加,植物总地上生物量呈现下降趋势,其中禾本科以及豆科植下降明显,高原鼠兔的选择性采食使得个别植被退出高寒草甸群落结构,与此同时另一种植被获得发展的机会,即一种植被被另一种植被所取代,植被的不均匀性增加,果洛地区植被群落结构的变化验证了这一结论[39]。
不同鼠洞梯度对植被多样性指数形成干扰,植被多样性指数与丰富度指数在低鼠兔洞穴梯度时数值较高,说明伴随鼠兔栖息数量的增加会导致植被群落结构的减少,丰富度指数下降;与无鼠洞数植被地相比,植被多样性和植被丰富度较高,但伴随鼠兔干扰的程度逐步加深,植被多样性与丰富度数值开始呈现逐步下降的趋势,鼠兔活动对于植被群落结构的影响显著。均匀度指数伴随不同鼠洞梯度的逐渐变大,指标数值呈现相同增加的趋势,说明鼠洞梯度与均匀度指数存在线性相关,高原鼠兔洞穴的选择会倾向于分布不均匀的植被群落结构,当植被不均匀性逐步增大时,鼠兔迁移居住的倾向性越明显,在植被分布不均匀,植被盖度低且斑块状裸地分布广泛的地区这种趋势更明显[43],可见植被的不均匀性也是诱使高原鼠兔迁入的因素之一[44-45]。
4 结论
高原鼠兔活动会对高寒草甸生态系统演替产生影响,当高原鼠兔干扰维持在较低水平时,土壤水分含量较低,植被群落结构分布不均匀且植被盖度低矮,裸地面积较少,此时不存在鼠兔种群密度过高的风险,鼠兔活动有利于促进高寒草甸正向演替,反之,当高原鼠兔干扰程度较高且超过环境的承载能力时则会对高寒草甸生态系统产生负面影响;同时,与鼠兔适宜生存环境相关联的植被土壤因子也可作为草地生境退化的指示指标,为进一步确定适宜鼠兔种群防控阈值,准确监测和合理区划鼠害界限提供数据支撑。
猜你喜欢
红领巾·探索(2022年11期)2023-01-06
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
数学物理学报(2022年1期)2022-03-16
数学物理学报(2021年6期)2021-12-21
应用数学(2020年2期)2020-06-24
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
活力(2019年21期)2019-04-01
小哥白尼(趣味科学)(2018年10期)2019-01-16
现代装饰(2017年9期)2017-05-25
