
物种保护的理论基础
——从岛屿生物地理学理论到集合种群理论
2019-05-24 08:44赵淑清方精云雷光春
中国学术期刊文摘 2019年9期
赵淑清 方精云 雷光春
许多自然环境都可看成是大小、形状和隔离程度不同的岛屿,而且岛屿有许多显著特征,如地理隔离,生物类群简单。这些特点为岛屿的生物学特征的重复性研究和统计学分析提供了方便。岛屿为发展和检验自然选择、物种形成及演化,以及生物地理学及生态学等领域的理论和假设提供了重要的天然实验室。所以Mac Archur 和Wilson的岛屿生物地理学理论一经提出就引起学术界广泛关注,并被迅速接受,而逐渐成为物种保护和自然保护区设计的重要理论依据。但是目前许多以前是连续分布的物种,由于生境的破碎化而以集合种群的方式存在,物种的灭绝也往往经历了集合种群的阶段。因此,近年来集合种群成为保护生物学所关注的研究热点。人类活动所造成的物种灭绝,事实上是从局部灭绝开始的。局部灭绝的后果可能导致物种的最后灭绝。集合种群理论关注的恰恰是具有不稳定局部种群的物种区域续存条件。因此,随着人类和其他生物赖以生存的环境破碎化程度的加剧,有关集合种群的概念和理论将会被广泛地应用于物种保护研究。
实际上,由于人类的干扰,栖息地的破碎化已成事实。大范围内建立完整的保护区已很困难。在这种情况下,集合种群理论在物种保护上更具现实意义。
1 岛屿生物地理学理论的兴衰
1.1 岛屿生物地理学理论
很久以前人们就意识到岛屿的面积与物种数量之间存在着一种对应关系,但是20 世纪60年代以前在岛屿生物地理学中基本上没有定量的理论。 Mac Archur 和Wilson的岛屿生物地理学理论定量阐述了岛屿上物种的丰富度与面积的关系,其关系式通常用(1)式表示:
式中,S代表物种丰富度,A代表岛屿面积,C为与生物地理区域有关的拟合参数,Z为与到达岛屿难易程度有关的拟合参数。
岛屿生物地理学理论首次从动态方面阐述了物种丰富与面积及隔离程度的关系,认为岛屿上物种的丰富度取决于新物种的迁入和原来占据岛屿的物种的灭绝。这两个过程的相互消长导致了岛屿上物种丰富度的动态变化。当迁入率与绝灭率相等时,岛屿物种数达到动态的平衡状态,即物种的数目相对稳定,但物种的组成却不断变化和更新。这种状态下物种的种类更新的速率在数值上等于当时的迁入率或绝灭率,通常称为种周转率(species turnover rate)。这就是岛屿生物地理学理论的核心内容。
岛屿生物地理学理论的提出和迅速发展是生物地理学领域的一次革命。大量资料表明,面积和隔离程度确实在许多情况下是决定物种丰富度的最主要因素,而且生物赖以生存的环境,大至海洋中的岛屿、高山、林地,小到森林中的林窗都可以视为大小和隔离程度不同的岛屿。岛屿生物地理学理论的简单性及其适用领域的普遍性使这一理论长期成为物种保护和自然保护区设计的理论基础。
1.2 岛屿生物地理学理论的局限性
岛屿生物地理学理论把岛屿的几乎所有的生物学特征都归结为一个变量,即物种的数量。它只关注岛屿上物种的数目与面积的关系,不考虑同一物种内部个体的大小和数量,而这恰恰是种群生物学最本质的特征之一,也是物种适应其所处异质环境的结果。物种数目和面积的关系在某种程度上是环境异质性的量度,而它作为岛屿生物地理学理论的核心内容,其本身反映了MacArchur 和Wilson 意识到环境异质性的重要性。然而,许多环境异质性量度指标不是简单地随面积增加而增加的,因此,物种数量与面积的关系也就很难准确地反映环境的异质性。这是岛屿生物地理学理论用来指导物种保护的一个最大缺憾,因为有效的物种保护应该包涵物种本身的保护及其栖息地异质生境的保存。然而,反过来,这也可能是该理论得以提出,进而为人们普遍接受的原因之一。因为环境异质性是一个非常复杂的问题,将其同物种发生和群落格局结合起来进行定量化描述就更难。
岛屿生物地理学理论用来指导物种保护的另一个局限性在于,该理论认为决定岛屿物种平衡的主要过程是随机的,而且对岛屿上所有物种都是均等的。另外,该理论也没有考虑其他决定岛屿群落结构的重要生态学因素,如竞争、捕食、互惠共生和进化等。在岛屿生物地理学理论中,存在一个永远都不会灭绝的大陆种群的假定,而实际上,这一假定是难以成立的,因为环境随机性和各种灾难可能会导致大陆种群的灭绝。Hanski 和Simberloff的研究表明,即使在特别大的保护区,如像美国最大国家公园的尺度,物种也可能遭受灭绝。
岛屿生物地理学理论中存在的局限性告诫人们,在物种保护过程中,要正确地领会岛屿生物地理学理论的精髓,不要死搬硬套。因为它的产生有其特定的范畴,它的应用也应该是有条件的。
1.3 岛屿生物地理学留给物种保护的启示
由于岛屿生物地理学理论起源于海洋岛屿中的物种研究,将其广泛应用于陆地生境岛屿研究中有其局限性是自然的。但它为物种保护提供了十分宝贵的理论财富,如将许多现实的生境喻为岛屿、物种丰富度与面积关系模式的建立、试图寻找小物种灭绝的原因、避难所设置的原则。不管这一理论存在着什么不足,它使物种保护的研究由仅仅通过定性比较来描述,转向通过野外模拟实验来验证物种形成的机制。尽管人们对岛屿生物地理学理论进行多方面的批评和修正,但这并不否认其应有的应用价值。Mac Archur-Wilson 学说的产生和发展丰富了生物地理学理论和生态学理论,促进了人们对生物种多样性地理分布和动态格局的认识和理解。
2 集合种群理论的兴起
集合种群是指在相对独立的地理区域内,由空间上相互隔离,但又有功能联系(一定程度的个体迁移)的两个或两个以上的局部种 群组成的种群镶嵌系统。集合种群生物学的概念最初可追溯到Wright的遗传漂变理论。早在1931年,Wright 就指出,存在局域灭绝和重新定殖的种群进化过程可能会快一些。但是真正具有生态学内涵的集合种群概念是体现在1954年的Andrewartha 和Birch《动物生态学》一书中。在基于大量昆虫种群生物学实验的基础上,他们发现,长期以来为人们所接受的种群控制理论中密度制约学说是站不住脚的,局域物种的灭绝是一种普遍现象。同时,他们也认识到在空白地域物种重建的可能性。但是,由于人们当时对于种群密度调节制约的无条件接受,使得这一刚刚萌芽的集合种群概念沉寂了10 多年之久。当然,在此期间Mac Arthur 和Wilson 的岛屿生物地理学动态理论在某些方面与集合种群理论概念具有共性。
真正使用集合种群(Metapopu-lation)一词是从Richard Levins 开始的。尽管在早期的研究中,学术界已注意到局部种群的频繁灭绝和新的局部种群不断建立的过程,但Levins 从一个全新的角度研究了这一问题,为集合种群的发展奠定了坚实的基础。可以说,Levins 的工作标志着近代集合种群生物学理论的开端。但是,由于岛屿生物地理学理论在保护生物学中几乎不可动摇的地位,在1991年Gilpin 和Hanski 出版《集合种群动态:经验和理论研究》第一本系统介绍集合种群理论的巨著之前,集合种群理论对于大多数生物学家还是陌生的。这本书问世之后,有关集合种群研究的文章以指数形式增长,目前集合种群概念在种群生物学及其他相关领域中已经牢固的建立 起来。
3 物种保护理论范式从岛屿生物地理学向集合种群理论的转变
经过20 世纪70年代的一阵研究热潮之后,岛屿生物地理学理论的准确性和可应用范畴充满争议,特别是20 世纪80年代未期以来,越来越多的生态学文章开始怀疑岛屿生物地理学理论的真实性,人们开始慢慢将注意力转向被遗忘多年的集合种群理论,而且现实景观破碎化的增加也确实使得许多物种符合集合种群研究的结构。特别是近几年来,有关集合种群研究的文章以指数形式增长,大有代替岛屿生物地理学在保护生物学中地位之势。事实上,仅仅简单的统计数字是不足以说明一种范式转变的原因的。物种保护理论范式由岛屿生物地理学理论向集合种群动态理论的转变主要是由下列原因造成的。
(1)近20年来,生态学思维的一个重要变革反映在研究重点的转移,即由强调平衡性、均质性、确定性和单尺度或小尺度现象到开始重视非平衡、不稳定性、异质性、不确定性、以及多尺度和等级特征。当然,岛屿生物地理学理论也是动态的,但却更强调平衡状态时物种的丰富度,甚至在平衡状态时将岛屿的迁入率与灭绝率视为恒定,因此,从本质上讲是属于平衡理论的范畴;而集合种群理论是研究局部种群灭绝和重新定殖过程的生态学理论,显然已不再是平衡理论。物种保护理论从岛屿生物地理学到集合种群理论的转变显然是顺应这种潮流。但事实上即使非平衡理论也不否认系统平衡状态的存在,只是强调生态学系统很少或根本不停留在平衡态而己。此外,平衡理论的倡导者们(如R.H.Mac Archur, R.Levins,R.M.May)通常假定平衡状态的存在是为了简化数学上的分析处理过程,但他们对过渡态行为同样感兴趣,并做了不少这方面的工作。
(2)随着自然生境的加速丧失和破碎化,空间在构成生态格局和塑造生态过程所起的作用越来越受到人们的重视。岛屿生物地理学理论基本上忽略了物种个体的变化而强调物种丰富度的平衡模式,所以这个理论的空间概念是模糊的;而集合种群理论则通过空间明确模型(Spatially Explicit Model)将生态过程和地理信息系统(GIS)分析工具相联合,系统运用有关空间信息和生态学知识,并将计算结果予以空间直观表达,从而加强了过程模型的预测能力以及运用GIS 的空间分析功能。
(3)保护生物学中一条最基本的原则就是考虑遗传学上的信息,尤其是小种群内部遗传漂变和近亲繁殖的研究。这样的话,种群层次要比群落层次的研究对于遗传学更为重要。岛屿生物地理学研究的核心是岛屿中的物种数量,显然是属于群落水平的研究;而集合种群强调的是同一种群内部不同局部种群之间物种个体的交流,种群水平为其研究对象。
(4)集合种群模型拯救了那些被岛屿生物地理学理论忽略的小生境,这样的一些特殊生境可能会拥有数量惊人的物种以及一系列的地方特有种,而且局域暂时存在的物种(如逃逸物种和演替早期物种)的保存对于区域物种长期续存是至关重要的。岛屿生物地理学将重要的生态学数据解释为简单的物种—面积关系,面积越大,物种绝灭率越低,因而物种的丰富度也就越严重,显然大面积的地域为岛屿生物地理学理论所期望。
(5)除了存在一个永远都不会灭绝的大陆种群,岛屿生物地理学理论基本上类似于集合种群理论的一种模式(在集合种群模型中,任何局部种群即使是一个非常大的局部种群也有灭绝风险),所以任何以一而概全的理论都是不科学的。
(6)当前对于集合种群动态理论具有如此浓厚兴趣的一个最重要的原因就是栖息地破碎化程度越来越高,以及由此带来的对许多物种长期续存的威胁,而集合种群理论关注的恰恰是具有不稳定局部种群物种的区域续存条件,避免物种的局部灭绝,乃至物种的最后灭绝。显然仅仅关注物种丰富度和面积关系的岛屿生物地理学理论对于此类问题的解决就显得力不从心。
4 集合种群动态理论
4.1 Levins 模型
Levins 模型是经典的集合种群概念框架模型。Levins 首先区别了单种种群动态与一个局部种群的集合动态之间的不同。Levins 模型假定有大量的离散生境斑块,并假定它们大小相同,而且相互之间通过个体的迁移,使各局部种群均等地连接在一起。在Levins 模型中,生境斑块被分为已定居和未被定居两种,而局部种群的真实大小则忽略不计。它假定所有种群有恒定的灭绝风险,种群建立的概率与斑块有种群定居的比例(P,迁移源)及当前未被定居斑块比例(1-P,迁移目标)成正比。按上述假定,P的变化速率为:

其中,m、e分别为侵占和灭绝参数。P的平衡值为:

从Levins 模型可以看出,在平衡状态时被定居的生境斑块比例 将随e/m比值的减少而上升。只要e/m<1,集合种群将能持续生存下 去(P>0)。Levins 模型虽然简单,但它抓住了典型集合种群动态的一个关键属性:集合种群若要持续生存,局部种群的重建速率必须高到足补偿灭绝速率。另外,e/m<1 或m/e>1 表明一个被空白斑块包围的局部种群(当P很小时)在其生存期(1/e)内必须至少建立一个新种群才能使集合种群持续生存下去。Levins 模型预言:物种在某一生境定居的比例P随生境斑块平均大小及密度的下降而下降。如果斑块太小或相距太远,集合种群都会灭绝(P=0)。
正如Hanski 等所指出的那样,无论是作为一个概念性的模型,还是作为一个数学的工具,Levins 模型在种群生态学中不仅是一个有价值的模型,而且也是为在这一领域中开展进一步的数量研究工作迈出的最重要的一步。
4.2 关联函数模型
Levins 模型只是研究种群的灭绝和形成之间的平衡,并不含有斑块质量和空间位置的信息,因而也就不能用于预测特定集合种群的动态。在这种情况下需用空间现实的集合种群模型(spatially realistic metapopulation models)。目前这类模型主要有3 种类型:种群模拟模型、状态转移模型(state transition models)和关联函数模型(incidence function models)。在所有这些模 型方法中,关联函数模型(IFM)最简单,并在许多情况下是最有用的。
关联函数模型是一个基于单个生境斑块的一阶线性马尔可夫链模型,其中的生境在定居与未定居状态间有恒定的转换概率。如果斑块i在当前是空白的,设其单位时间内它被重新定居的概率为(Ci);如果斑块i在当前时间是被定居的,设种群在单位时间内灭绝的概率为Ei。按以上假设,斑块i被占领的静态概率(stationary probablity)可称为该种在斑块i的关联Ji:
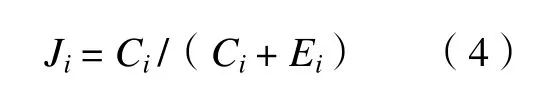
为了进一步了解模型的动态,首先需要更具体的有关斑块特性是如何影响斑块被重新定居的概率(Ci)及种群灭绝概率(Ei)的假设。这些假设是基于在特定斑块中种群大小的期望值随面积增加,而灭绝概率是随种群大小的下降而增加的这样一个基本原则:

通常来说,灭绝概率取决于斑块面积(灭绝概率依赖于种群大小,而种群大小依赖于斑块面积,并不特别依赖于斑块间的隔离程度)。其中,Ai是斑块A的面积,e,x是两个参数。在这个公式中存在一个最小斑块面积A0;当面积小于或等于A0时斑块内物种的灭绝概率为1。将灭绝概率与斑块面积联系起来是因 为斑块面积的数据比较容易获得。定居概率Ci是单位时间内到达斑块i的迁入者数量Mi的递增函数。为了方便起见,首先考虑简单的大陆—岛屿型集合种群:

其中,di是斑块(岛屿)i与大陆间的距离,T和U为两个参数。对常见种来说,它可以立即定居一个几乎没有隔离的斑块(di接近零),因此在方程(5)中可设U=1。在现实的集合种群模型中,Mi为来自现存种群迁移个体的总和。将这些种数的大小和距离考虑在内,可以有:

其中,Pj在被占领斑块取值为1,在空白斑块取值为0。dij为斑块i和j间的距离,T、U为两个参数(同方程5)。如果在新种群定居过程中迁入者之间无相互作用,Ci将随Mi指数增长。但通常情况下新种群成功定居的概率依赖于传播体(Propa)的大小。在这种情况下,Ci随Mi变化,呈“S”形增长:
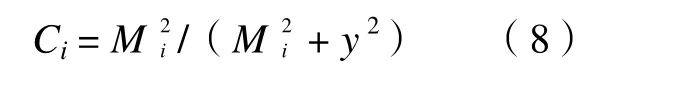
其中,y是一个新参数:当将方程(7)代入方程(8)只剩下参数组合y/U,以下用y表示。将方程(5)和方程(8)的Ei、Ci代入主程(4),并将拯救效应(rescue effect)考虑在内,也就是将因迁入造成的局部种群灭绝率下降考虑在内,于是得到:模型中的参数a、x、e和y可以通过非线性最大似然回归或其他方法估计(non-liner maximum likelihood regression)。

关联函数模型最大的优点在于它能够对特定的集合种群动态进行定量预测。能够了解在原始生境斑块、经过修饰的生境斑块及某些完全不同的生境斑块中,物种变化的瞬时和平衡动态,从而可以回答有关集合种群动态的许多实际问题。比如物种在某些特定的生境斑块集合中能否作为集合种群续存下去,生存于当前斑块网络中的种能否在某些斑块被除去或面积减小时仍然能够续存下去。
5 集合种群理论存在的不 足及其在其他领域的应用前景
虽然集合种群空间现实模型可以含有斑块质量和空间位置的信息,可用于预测特定集合种群的动态,但是集合种群理论经常是在尚未弄清是否满足其基本假设的前提下被使用的。如果保护策略基于的集合种群动态是不存在的,那么这样的保护行动就不可能达到预期的目标;如果集合种群内部局部种群之间的运动太过频繁,那事实上所研究的种群是同一个简单的种群,而不是集合种群;甚至可能是将一个由于内部交流不可能而只可能与其邻近种群相互作用的太大种群视为集合种群。
集合种群模型预测只有在局部种群栖息地的生境斑块不是太小而导致过高的物种绝灭率,也不可过于隔离而阻碍局部种群的重新建立的前提下,物种的长期保存才成为可能。当前栖息地破碎化程度如此之高,而且物种对生境的破碎化作出反应存在时滞,因而就不知道目前破碎景观中到底有多少“活着的死者”(living dead)。因此,当生境破碎化速度达到或超过集合种群绝灭—定殖的平衡过程时,集合种群将走向灭绝。这将向世人敲起警钟:随着生境斑块面积的降低及斑块隔离程度的增加,物种首先会从这些最小的生境斑块中消失,大的生境斑块中的物种存在的时间可以稍长一些,但是即使是在最大的生境斑块中的物种,其可存活时间也只能是在可预测的几十年或几百年的尺度上。这些物种一个接着一个消失,局部种群之间的隔离程度又过大,物种的重新定殖不再可能或可能性很小,集合种群理论对物种的保护也将不再起作用。集合种群产生于破碎化的景观中,但这并不是因为这样的景观结构对于物种的长期续存已经足够,而只是降低物种绝灭速率的一个补救的办法。
集合种群是从经验和理论上研究具有不同空间特征种群通过扩散、迁移和基因流来维持的生态及进化动态。这一理论对于许多应用问题,如日益加速的破碎化景观中物种的分化和种化(speciation)及遗传多样性的保存,乃至一些抽象的机制研究,如利他行为的群体选择,正像Hanski 等的观点,除非有这样一个模型,它能够描述局部种群的灭绝和新的局部种群的建立之间的平衡,否则要想了解这个过程的遗传学意义是不可能的。对于捕食与被捕食的平衡,寄主和寄 生物的动态都是一个非常好的概念工具。
6 结语
事实上,集合种群理论和岛屿生物地理学理论是既有联系又有区别的两个物种保护的理论基础。在这两个理论体系中有一个共同的基本过程,即个体迁入并建立新的局部种群以及局部种群的灭绝过程。十几年来,岛屿生物地理学关于SLOSS(single large or several small)的争论,即物种保护计划到底是应该建立一个大的生境,还是建立相互联系的小的生境的争论,从根本上讲是一个集合种群的问题。
当然这两个重要的理论体系之间仍然是有区别的,其中最重要的区别就是在岛屿生物地理学中总假定存在一个所谓的“大陆”,并且在这个大陆上的“大陆种群”不仅不会灭绝,而且所有迁移个体都只能来自于“大陆种群”;在集合种群中迁移个体可以来自于任何一个现存的局部种群,同时任何一个局部种群都是有可能灭绝的。显然集合种群模型更具现实意义。
岛屿生物地理学关注的是岛屿上物种的丰富度,是从群落水平上研究物种的变化规律,对物种多样性的保护可能更具理论意义;而集合种群考虑同一物种的不同个体之间的交流以及物种内部个体之间的基因交流,是从种群水平上研究物种的消亡规律,从生物多样性的层次考虑,应该是侧重遗传多样性,因而对于濒危物种的保护是更有效的方法论指导。物种数目的变化最终还要取决于单个独立物种的灭绝或再定居,因此从种群水平上探讨物种灭绝规律对于理解群落的变化是至关重要的。从这个意义上讲,岛屿生物地理学理论与集合种群理论是可以相互借鉴和补充的。
另外,值得一提的是,有时理论模式的转变也受到文化意识的影响。在北美的一次生物地理学研讨会上交流的11 篇论文中有9 篇论文引用了岛屿生物地理学理论的原著和相关论文,而时隔一年,在欧洲的一次类似领域的学术活动中交流的9 篇文章中没有一篇参考有关岛屿生物地理学理论的论文。北美生物地理学家及欧洲生物地理学家对于岛屿生物地理学理论衷爱程度的悬殊在一定程度上反映了两大陆地缘文化的差异,因为岛屿生物地理学理论产生于北美,而重新认识集合种群科学价值的是欧洲的生态学家。
总之,在物种保护过程中,应该将注重格局研究的岛屿生物地理学与强调过程研究的集合种群理论相互补充和结合,从而为物种保护提供更为全面合理的理论指导。〓
(摘自《生态学报》2001年第7期)
猜你喜欢
今日农业(2022年15期)2022-09-20
思维与智慧·上半月(2022年4期)2022-04-08
湖南电力(2021年1期)2021-04-13
落叶果树(2021年6期)2021-02-12
四川文学(2020年10期)2020-02-06
红土地(2018年7期)2018-09-26
现代园艺(2018年1期)2018-03-15
体育科技(2016年2期)2016-02-28
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
儿童故事画报(2015年5期)2015-07-22
