
铁皮石斛分子遗传学和多组学研究进展
2019-05-17 09:36乔宇琛周思静宋梅芳王平刘桂君
生物技术通报 2019年3期
乔宇琛 周思静 宋梅芳 王平 刘桂君
(1. 北京市辐射中心,北京 100015;2. 北京师范大学射线束技术教育部重点实验室,北京 100875)
铁皮石斛(Dendrobium officinaleKimura et Migo,D. officinale)是一种兰科(Orchidaceae)石斛属(Dendrobium)的多年生草本植物,是一种古老、极具药用价值的中药材,药用部分主要是新鲜的茎或干燥的茎,又称“石斛枫斗(Dendrobii officinalisCaulis)”,性甘,微寒,有益胃生津、滋阴清热的功效[1-2],享有“九大仙草”之首的美誉[3],且兼具较强的观赏性。铁皮石斛野生资源在自然条件下需要与某些真菌共生才可萌发、加上过度采挖和生境破坏而濒临灭绝。1988年,国务院发布的《野生药材资源保护管理条例》,将铁皮石斛列为三级保护品种[4];1991年,在《中国植物红皮书》中被收录为稀有濒危植物[5];2018年5月正式进入药食同源植物名录。由于其药用价值较高,市场需求量大,且现在人工栽培技术也较为成熟[6-7],我国铁皮石斛产业近年来快速发展,年产值近100亿元[8]。
鉴于铁皮石斛重要的食药用价值和巨大的应用前景,已成为近年来人们研究的热点。但大部分研究还是集中在成分提取、多糖的功能、产品开发及人工栽培技术开发与优化等方面[9-12]。然而,随着不断迭代的测序技术的出现[13],以及新的分子生物学方法的开发[14],对铁皮石斛在分子生物学方面的研究也越来越多。本文综述了近期国内外有关铁皮石斛在分子遗传学、基因组、转录组、蛋白质组与代谢组方面的研究,旨为今后更好的从分子生物学角度开发应用铁皮石斛这一重要资源提供科学依据。
1 铁皮石斛分子遗传学
1.1 分子鉴定与遗传多样性分析
建立DNA指纹图谱、筛选合适的分子标记技术,有利于鉴定和筛选铁皮石斛优良品种、规范市场秩序与规避掺假。目前用于铁皮石斛鉴定的分子标记技术有随机扩增多态性DNA(Random amplified polymorphic DNA,RAPD)及相关技术、序列相关扩增多态性(Sequence-related amplified polymorphism,SRAP)及相关技术、简单序列重复(Simple sequence repeats,SSR)及相关技术、扩增片段长度多态性(Amplified fragment length polymorphism,AFLP)及相关技术、目标起始密码子多态性(Start codon rargeted polymorphism,SCoT)和 基因芯片(Genechip)等。
同时,上述标记技术还可用于铁皮石斛的遗传结构和多样性分析。铁皮石斛作为珍稀濒危的重要食药两用植物,确定其遗传结构和多样性水平可有利于铁皮石斛种质资源的保护和可持续利用。多项研究发现不同地区的铁皮石斛在物种水平上具有丰富的遗传多样性,而居群间遗传分化程度较低[15-17]。可能的原因是其多生长在悬崖峭壁上,自然繁殖能能力较低,居群间的基因交流少[18]。此外,自交不亲和性也是保持铁皮石斛在物种水平上具有较高遗传多样性的重要原因[19]。一个物种的遗传多样性水平还会受到地理分布的影响,一般而言,广泛分布的物种的遗传变异水平可能比狭窄分布的物种的遗传变异水平要高[20]。在我国铁皮石斛野生种质资源较为广泛地分布于安徽、福建、广东、广西、贵州、湖南、江西、四川、台湾、云南和浙江等省。
1.1.1 RAPD标记及相关技术 RAPD标记技术是较早,同时也是应用最广的一种分子鉴定手段。张铭等[21]对石斛属内26 个种的基因组DNA进行RAPD分析,可从其中方便快捷地鉴别出铁皮石斛。丁鸽等[15]采用RAPD标记技术,找到引物S412可以有效鉴别铁皮石斛的8 个野生居群,且居群间具有较丰富的遗传多样性。王慧中等[22]采用RAPD标记技术筛选出10 对随机引物可以很好地区分13 种石斛属植物,且发现供试石斛属植物遗传多样性高。金波等[23]以扩增得到的铁皮石斛特异RAPD序列为基础,设计出一对新的特异性引物,扩增得到了300 bp序列特异性扩增区(Sequence characterized amplified regions,SCAR)标记片段,能特异性的快速鉴定铁皮石斛样品。SCAR标记是由RAPD标记转化而来,所用引物较长,引物序列与模板DNA完全互补,具有良好的扩增严谨性,且扩增结果稳定性好、重复性强。
1.1.2 SRAP标记及相关技术 SRAP标记技术是一种操作简单、产量中等、成本低、可信度高和易于测序的分子鉴定方法。Ding等[16]采用SRAP标记技术分析了来自9 个野生居群的84 个铁皮石斛个体,检测到物种水平上较高的遗传多样性,但与具有相似生活史特征的其他物种相比,居群水平上的遗传多样性较低。包英华等[17]采用SRAP标记技术分析8个铁皮石斛居群的DNA条带多态性,最终筛选出7对有效的SRAP引物组合,经过遗传分析发现铁皮石斛种质资源的遗传多样性水平较高,且居群内的遗传分化偏高于居群间。李怀志等[24]使用25对SRAP引物对供试的铁皮石斛进行遗传多样性分析,共获得86 条多态性条带;20 份铁皮石斛品系间Jaccard相似系数为0.002-0.821,显示出供试铁皮石斛有较为丰富的遗传变异;同时,构建出20份铁皮石斛品系的SRAP特征指纹图谱。
靶位区域扩增多态性(Target region amplification polymorphism,TRAP)标记技术是从SRAP标记技术改进而来,基于已知的cDNA或表达序列标签(Expressed sequence tag,EST)序列信息,较易将性状与标记相关联,具有操作简单、效率高、共显性、重复性好和易测序等优点[25]。刘玲等[26]利用铁皮石斛多糖生物合成途径中的尿苷二磷酸葡萄糖焦磷酸化酶基因、磷酸烯醇式丙酮酸羧化酶基因以及石斛碱合成过程中苯丙氨酸解氨酶基因,设计构建TRAP分子标记,对铁皮石斛野生居群及其杂交后代进行鉴别。最终从54 对TRAP引物中共筛选出7 对引物,可用于鉴别铁皮石斛野生居群,并且在杂交后代中可同时检测到亲本特征条带和新产生条带,可有效地鉴别出亲本及其杂交后代。
1.1.3 SSR标记及相关技术 SSR标记技术又名微卫星技术[27],由于具有高度可变性、可重复性、共显性、特异性及技术难度低,且在全基因组中随机分布等优点[28],在分子鉴定中应用较多。谢明璐等[29]利用该技术成功开发出4 对引物,能够对铁皮石斛种质纯度进行鉴定。徐蕾等[30]对36 份来自云南、浙江、广东、广西等地的铁皮石斛进行SSR分析,共获得249 个SSR标记位点,遗传分析发现供试的中国铁皮石斛品种具有较高的遗传多样性。Hou等[31]设计三核苷酸SSR标记对铁皮石斛遗传多样性进行分析,最后成功获得15 个新的SSR标记。Xu等[32]对来自4 个自然居群的55 个铁皮石斛植株的多态性水平进行测定。从设计的15 对叶绿体(Chloroplast,cp)SSR引物中找到9 对具有多态性。在另一组51个石斛属个体(属于17 种不同物种)上验证9 对cp SSR引物的可转移性,发现3 种标记物可以转移到所有测试的物种,而其余的6 种标记物在大多数测试物种中可成功交叉扩增。Xu等[33]对2个铁皮石斛栽培品种的2 年生茎进行转录组测序。共获得51 683 个的全长转录本,平均长度为505 bp,获得40 405 个非重复序列基因(Unigenes)。从7 332个(18.15%)Unigene序列中共鉴定出8 527 个潜在SSRs,其中1 023 个(2.53%)Unigenes含有多于一个的SSR标记。此外,还设计了68 个基因座特异性引物对,其中17 对可产生多态性产物。由于它们具有高度的多态性和可转移性,这些基因型SSR标记不仅可用于石斛种质资源保存,而且还可用于石斛的系统发育研究。
基于SSR标记技术,简单重复序列间(Intersimple sequence repeat,ISSR)的DNA标记技术得以开发,它具有耗资少、DNA用量少和多态性高的优点,且无需知道任何靶标记的序列背景。Shen等[34]利用该技术鉴定8 个铁皮石斛居群,从76 对设计的ISSR引物中筛选到10 对引物,扩增出127 个DNA片段,多态性条带为90.5%,最终找到16 个特定的鉴别标记。为了提高鉴别效率,使用6 个多态性条带来构建ISSR指纹识别码,用于验证铁皮石斛居群。结果发现铁皮石斛8 个野生居群均可采用ISSR准确鉴定。Ding等[35]利用RAPD和ISSR标记技术对9个野生的铁皮石斛居群进行遗传多样性分析,得到89%的多态性条带,结合分子方差分析法(Analysis of molecular variance,AMOVA)发现居群内存在78.88%(RAPD)和78.84%(ISSR)的种间变异,居群间变异与地域无显著相关性。9 个铁皮石斛野生居群具有丰富的遗传多样性,并且ISSR的多样性检测优于RAPD。
随机扩增微卫星多态性(Random amplified microsatellite polymorphism,RAMP)标记技术是一种结合SSR与RAPD优点而发展起来的新标记技术,具有共显性特点,不需要开发特异性引物,多态性丰富,重复性强,分析较为简便[36]。能更好的揭示物种间的亲缘关系,且更适合于遗传背景尚不太清楚物种的遗传多样性研究。沈洁等[37]设计出16 对引物,利用RAMP标记技术对9 个野生居群的112株铁皮石斛进行检测发现,不同居群间存在较丰富的遗传多样性。对RAMP扩增条带数据进行聚类分析,发现铁皮石斛居群地理分布的远近与居群间遗传关系的亲疏呈现良好的相关性。
1.1.4 AFLP标记及相关技术 AFLP标记技术1992年由Zabeau等[38]创立,较之前的标记技术,其可信度、灵敏度和经济实用等方面都有了较大的提升。王慧中等[39]采用AFLP标记技术,选择3 种扩增引物组合分别对13 种石斛属植物进行扩增,获得丰富的条带。着重分析了100-300 bp之间的条带,共获得346 条谱带,多态条带比率为98.8%,说明供试石斛属植物有着丰富的遗传多样性。聚类分析表明,在相似系数0.54处,可将13 种材料分为Ⅰ、Ⅱ、Ⅲ类,其中铁皮石斛分属于Ⅰ类。AFLP分析结果与传统分类学的结果基本一致,表明该技术可用于对石斛属植物的遗传多样性和分类研究。Li等[40]采用该技术对12 个铁皮石斛居群进行AFLP分析。结合POPGENE软件和AMOVA分析检测到高水平的遗传多样性(He = 0.269),发现ΦST值在0.047-0.578范围内的居群间存在中等变异,平均遗传变异为26.97%。非加权平均距离法(Unweighted pair-group method with arithmetic means,UPGMA)树状图显示3 个主要聚类,这与使用NTSYS软件进行主坐标分析的结果一致。提示保持一个稳定的环境对于这种珍稀濒危植物的原生境保存和管理至关重要,对于非原生境保存来说,设计一个完整的种质资源库非常有意义。
DNA甲基化是表观遗传修饰的基本方式之一,DNA甲基化与基因沉默有关,去甲基化可导致基因表达。随着对DNA甲基化研究的深入,在AFLP标记技术的基础上发展出甲基化敏感扩增多态性(Methylation sensitive amplification polymorphism,MSAP)标记技术,它操作相对简单,多态性高,引物设计也相对容易,可检测出样品DNA中大量的甲基化位点。李雅婷等[41]采用AFLP和MSAP标记技术,探讨铁皮石斛二倍体经染色体加倍后得到四倍体的过程中,遗传差异和基因组DNA甲基化水平及模式变化情况,分析铁皮石斛二倍体与同源四倍体的基因组变异及甲基化差异。AFLP结果显示,在25 对引物组合扩增的1 006 条带中,多态条带比率为47.12%;甲基化模式分析显示,铁皮石斛同源四倍体有28.63%的位点发生了去甲基化,30.63%的位点发生了过甲基化。铁皮石斛同源四倍体DNA碱基序列同其二倍体相比,总甲基率低,且甲基化模式也发生了较大调整。这为进一步探索四倍体铁皮石斛的表观遗传调控机制与基因组变异机理奠定了基础。关蕾等[42]利用MSAP标记技术研究了强光胁迫下铁皮石斛抗氧化系统与基因组DNA甲基化水平受外源硫化氢的影响发现,200 μmol/L的硫氢化钠(硫化氢的供体)可显著提高超氧化物歧化酶、过氧化氢酶和过氧化物酶的活力,同时降低丙二醛的含量,以此来缓解铁皮石斛在强光胁迫下造成的氧化损伤。高浓度硫氢化钠(600 μmol/L)则会加重铁皮石斛的氧化损伤。MSAP分析表明,同对照铁皮石斛35.4%的基因组DNA甲基化比率相比,强光胁迫下0、200和600 μmol/L硫氢化钠处理(T1、T3和T5)的甲基化比率均低。T1与对照相比、T3和T5与T1相比,铁皮石斛基因组DNA的去甲基化和甲基化变化位点的比例分别为6.4%、7.7%、6.7%和9.0%、8.1%和9.2%。通过对差异条带进行回收测序和Blast比对发现,其中有与光合作用有关的合成酶、与抗逆相关的转录因子等同源的片段。
1.1.5 SCoT标记技术 SCoT标记是Collard和Mackill[43]以单引物扩增反应(Start codon targeted polymorphism,SPAR)为基础开发的新型分子标记技术,它根据植物基因中的ATG翻译起始位点侧翼序列的保守性,设计单引物并对其基因组进行扩增,多态性丰富,且操作简单[44]。赵瑞强等[45]利用正交试验设计与单因素实验的方法,对铁皮石斛SCoT-PCR反应体系进行了构建和优化,以32 份铁皮石斛叶片为实验材料,验证发现该体系在遗传多样性分析中具有良好的稳定性与重复性。徐旭栋等[46]采用单一变量控制的方法获得铁皮石斛SCoTPCR 20 μL的优化反应体系。这些研究为铁皮石斛种质遗传多样性分析、分子标记辅助育种等研究提供了反应体系。随后,徐旭栋等[47]对20 份不同来源的人工栽培铁皮石斛样品进行SCoT分析。从设计好的56 对SCoT引物中最终筛选出20 对,共获得226 个扩增条带,总多态条带比率为90.3%。表明不同来源的人工栽培种铁皮石斛存在较大的遗传变异,提示目前市场中的人工栽培铁皮石斛具有丰富的遗传多样性,SCoT标记技术可用于铁皮石斛的遗传多样性研究。
1.1.6 基因芯片技术 基因芯片(也叫DNA微阵列)[48]可以利用特异性探针高通量地鉴别铁皮石斛的真伪。Zhang等[49]利用已有的16 种石斛的ITS1-5.8S-ITS2序列,设计出带有荧光标记的ITS2序列探针,利用基因芯片技术鉴定《中国药典》中收录的5 种药用石斛,可以对不同石斛种类实现有效区分。Sze等[50]利用5S rDNA基因间隔区序列,设计特异性荧光探针,以此开发出商业化基因芯片,可以有效区分铁皮石斛与其他种类的石斛。然而,基因芯片可以从遗传水平鉴别中药品种,但中药药效成分大多为次生代谢产物,基因芯片无法对中药的优劣进行鉴别,因此需与中药的化学成分指纹图谱等鉴定相结合,来评价铁皮石斛药用价值的优劣,发挥最佳作用[51-52]。
除了上述提到的标记技术,利用铁皮石斛的其他分子标记基因也可以实现鉴定的目的,如核基因组的rRNA、ITS,叶绿体基因组的rbcL、matK、rpoC,以及线粒体的nad1基因内含子等。丁小余等[53]通过扩增、测序21 种枫斗类石斛的rDNA ITS,初步建立出一个小型枫斗石斛的rDNA ITS数据库。Ding等[54]基于38 种石斛的rDNA ITS序列,设计出一对特异性PCR引物TP-JB01S和TP-JB01X,66℃退火时,只有铁皮石斛得以特异性扩增,可以快速有效地鉴定铁皮石斛。滕艳芬等[55]利用matK基因序列比较了《中国药典》收录的石斛和市场中常见的混淆品种发现,matK基因序列在非石斛属混淆品与正品石斛之间的差异要远大于正品石斛间的差异。Ding等[56]利用叶绿体基因matK和rbcL、核rDNA ITS区和线粒体基因nad1内含子序列对8 个铁皮石斛居群进行研究。除了在rDNA ITS区域发现9 个单核苷酸多态性(SNPs)位点之外,居群间matK、rbcL和nad1内含子序列中没有发现任何差异。设计两对基于SNP等位基因特异性的PCR引物来验证两个真实群体——GSG和FSC。GSG和FSC的等位基因特异性PCR(allele specific PCR,AS-PCR)片段大约为600和560 bp,退火温度分别在55-57℃和58-60℃。这两对引物同样也可以对来自真实群体的干燥的“枫斗”样品进行鉴定。
1.2 遗传转化
遗传转化可以实现外源基因定向导入受体细胞,是研究基因功能、分子改良的重要基因工程技术。按照作用方式不同,铁皮石斛的遗传转化可分为间接转化的农杆菌(Agrobacterium tumefaciens)介导的遗传转化法和直接转化的基因枪法。
农杆菌介导的铁皮石斛遗传转化体系已建立并进行了不断地优化。武荣花等[57]用农杆菌介导ACO反义基因对铁皮石斛原球茎(Protocormlike bodies,PLBs)进行遗传转化,找到优化的转化条件:经刀切法处理的PLBs,在乙酰丁香酮(Acetosyringone,AS)浓度为100 μmol/L、农杆菌菌液浓度OD600值为0.8的条件下侵染30 min,共培养60 h,接种到含有30 mg/L美罗培南(Meropenem,Mer)和3.0 mg/L草甘膦(Glyphosate,PPT)的筛选培养基中,遗传转化效率最高。筛选获得的抗性植株经β-葡萄糖醛酸糖苷酶(β-glucuronidase,GUS)组织化学染色和PCR鉴定,初步证明ACO反义基因已整合到铁皮石斛基因组中。崔波等[58]以铁皮石斛PLBs为受体,对农杆菌介导的铁皮石斛遗传转化体系进行优化发现,经摁压处理的PLBs,在AS浓度为200 μmol/L、农杆菌菌液浓度OD600值为1.0的条件下侵染30 min,共培养48 h,转入到含有50 mg/L Mer和2.0 mg/L PPT的筛选培养基中,遗传转化效率最高。经GUS组织化学染色和PCR分析鉴定,初步证明ACC合成酶反义基因已成功整合到铁皮石斛的基因组中。
除了农杆菌介导的遗传转化,基因枪法转化技术也在石斛中有应用。Kuehnle和Sugii[59]以石斛兰(Dendrobium× Jaquelyn Thomas杂交种)的 PLBs为受体,用含有植物可表达的Nos-NPT II基因和番木瓜环斑病毒(PRV)外壳蛋白(CP)基因的质粒pGA482GG/cpPRV4微粒进行轰击,得到来自4 个杂交系大约280 个原球茎,经过卡那霉素筛选培养和PCR鉴定发现,来自两个杂交系的13 株抗性植株带有Nos-NPT II基因,其中1 株同时带有PRVCP基因。Chia等[60]利用荧光素酶基因(Luc)作为组合标记/报告基因,成功对石斛兰进行了基因枪法转化。用1.3 μm由钨包覆、携带有35SLuc嵌合基因的质粒颗粒对石斛兰类PLBs进行轰击转化,经过荧光显微成像技术与Southern印迹分析证实Luc基因整合到石斛兰基因组中。杨雪飞等[61]以铁皮石斛PLBs为受体,利用基因枪法将来源于大麦的抗旱耐盐基因lea3进行遗传转化,经过PPT筛选检测和PCR鉴定,证明lea3基因已整合到6个株系7株铁皮石斛转化株的基因组中。这两种遗传转化体系的建立,有助于铁皮石斛基因功能的鉴定,可依照实验实际需求选择。
2 铁皮石斛基因组
2.1 基因组
基因是生物遗传的基本单位,基因组测序技术的问世,让人们可以从基因层面全面深入地揭示生物体的秘密。兰科植物占所有种子植物种类的10%左右,具有很大的经济价值,因其极具观赏性和良好的生态适应性而具有特殊的科学价值。Yan等[62]以云南省普洱地区可追溯来源的F3代人工自交型铁皮石斛为材料,整合二代Illumina和三代PacBio两大测序平台的优势,2015年首次从头组装出1.35 Gb的铁皮石斛基因组序列,杂合性0.48%。共预测到35 567 个蛋白质编码基因,所有的测序量合计覆盖超过了铁皮石斛基因组94%的序列,功能注释到铁皮石斛大约97.56%的蛋白质编码基因(34 699 个)。分析发现兰花有着完整的花序基因集,相比于其他单子叶植物,铁皮石斛具有特异的花序基因。与真菌共生和抗旱性有关的一些基因家族发生了明显扩张。另外,对铁皮石斛药用成分生物合成信号通路进行分析发现,与多糖生成相关的基因SPS和SuSy发生大规模复制,铁皮石斛石斛碱的合成信号通路可以从已有的研究结果基础上延伸到16-epivellosimine 的合成[63]。
但Yan等[62]报道的铁皮石斛的基因组序列,有着高度分散的装配,K-mer分析中存在多个峰,表明其序列可能源自人造杂种,这复杂化了铁皮石斛基因组的正确解读。Zhang等[64]于2016年对石生兰花——铁皮石斛(Dendrobium catenatum)基因组序列进行了报道。这两篇报道的研究对象均为铁皮石斛(表1),只是在学名叫法上有争议,后者认为铁皮石斛的学名应为Dendrobium catenatumLindl.[65],对于铁皮石斛的拉丁学名问题,有必要进一步商榷,本文暂不做过多探讨。Zhang等[64]研究发现铁皮石斛与蝴蝶兰(Phalaenopsis)拥有几乎相同的基因数,共享全基因组重复,为二倍体,基因组大小1.01 Gb,含有38 条染色体,共预测到28 910 个蛋白编码基因。分析发现铁皮石斛基因组中有许多抗性相关基因发生了扩张,表明其具有一个用以适应广泛生态位的强大免疫系统。涉及葡甘聚糖合酶活性基因的大量重复,可能与药用多糖的合成相关。参与调控发育和生长的MADS-box基因cladesANR1、StMADS11和MIKC*均有不同程度扩张,表明这些扩张可能与石斛属植物形态具有高度的多样性有关。相反,I型MADS-box基因家族的基因缺失,可能解释了种子胚乳的丧失。与其他植物基因组相比,铁皮石斛拥有更多的长基因和许多独特的重复。长末端重复序列(Long rerminal repeats,LTRs)占大约基因组的46%,通过计算它们的插入次数,发现铁皮石斛在过去的500万年中出现了一系列LTR活性,因此推断这些LTR是从蝴蝶兰属的物种中被插入到铁皮石斛基因组中的。
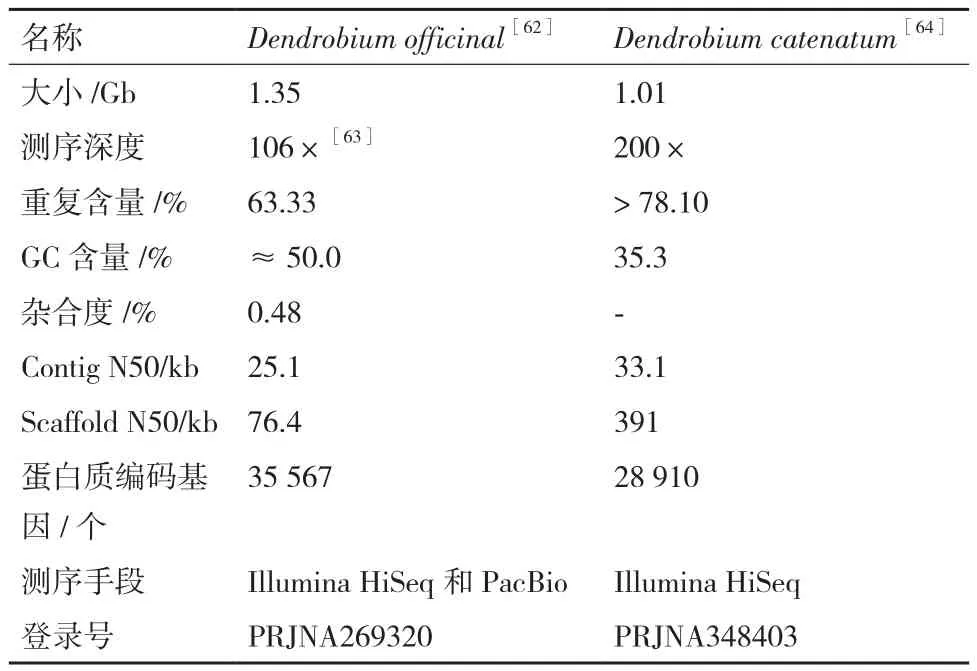
表1 铁皮石斛基因组特征
铁皮石斛基因组图谱的成功绘制,使铁皮石斛研究迈入了基因时代,不仅改变了铁皮石斛产业光凭经验发展的现状,而且为人工合成药用铁皮石斛活性成分奠定了分子基础,为品种鉴定、基因育种、成分和功能研究,以及药用成分的开发利用提供了重要资源,同时对珍稀食药两用植物的保护和优质种质资源品种优化也有重要意义。
2.2 叶绿体基因组
和全基因组不同,cp基因组相对较短(75-250 kb之间),主要为母本遗传[66],且大多数陆地植物的cp基因组含有高水平的插入和缺失,碱基置换的高发主要集中在特定基因和基因间隔上[67]。此外,完整的cp基因组具有大大超过常用DNA条形码长度的保守序列,可提供更多的差异来区分密切相关的植物,从而使其成为超级条形码[66]。cp基因组已成为研究植物物种鉴定、分类学和系统发育分析进化的常用方法。
Luo等[68]和 Yang等[69]均对铁皮石斛的 cp 基因组进行了报道,测序拼接后得到的结构一致,只是在片段大小与基因数目等方面略有不同(表2)。铁皮石斛cp DNA是一个长的环状分子,由一个大单拷贝区(Large single copy,LSC)、一个小单拷贝区(Small single copy,SSC)和一对反向重复序列(Inverted repeat sequence,IR)组成,其中两个IR将LSC和SSC隔离开。Luo等[68]还运用比较基因组的方法,对7 种光合兰花的cp基因组(大花杓兰,Cypripedium Macranthos;硬叶兰,Cymbidium mannii;D. officinale; 扇 叶 文 心 兰,Erycina pusilla;文心兰,Oncidium Gower Ramsey;蝴蝶兰,Phalaenopsis aphrodite;小兰屿蝴蝶兰,Phalaenopsis equestris)进行对比分析发现,它们在结构、基因顺序和含量方面显示出较高的相似性,只在反向重复/小单拷贝接合点和ndh基因上存在些许差异。
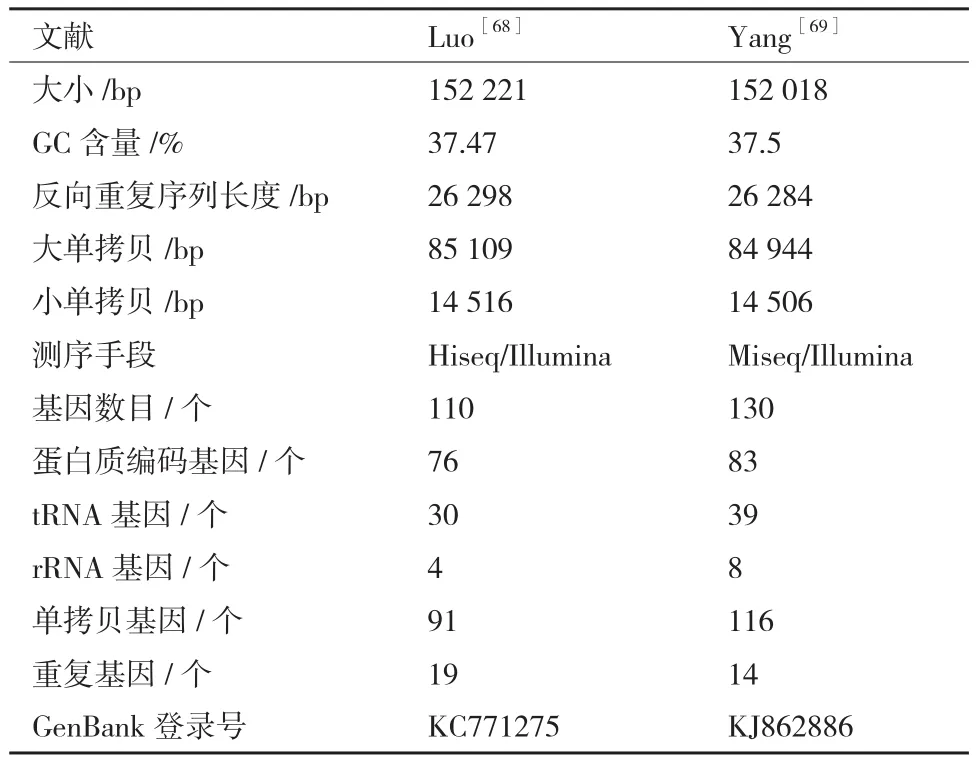
表2 铁皮石斛叶绿体基因组特征
3 铁皮石斛转录组
转录组可以高效全面地快速挖掘基因的表达、结构、功能及其调控机理,已经成为是食药用植物分子生物学领域较为成熟的研究手段。吴超等[70]应用高通量测序技术对铁皮石斛两年生叶片进行转录组测序,共获得11.15 Gb数据,GC含量47.77%。组装后获得121 596 个Unigenes,平均序列长度660 bp,功能注释到52 345 个Unigenes。基因本体(Gene ontology,GO)分析将Unigene功能分类分为生物过程、细胞组分和分子功能的3 大类57 个分支。蛋白质直系同源(Protein orthologous group,COG)数据库分析将40 842 个Unigene功能注释后分到25 类直系同源蛋白分类中,主要涉及如复制、重组和修复、转录、翻译、核糖体结构和生物合成等过程。京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)分析发现,Unigene可定位到脂类代谢、氨基酸代谢、碳水化合物代谢等128 个代谢途径分支上。Meng等[71]通过结合RNA、小RNA(Small RNA,sRNA)和降解组测序技术对铁皮石斛进行了转录组范围内器官特异性的研究。对铁皮石斛4 个器官(花、叶、茎和根)进行RNA测序,共获得536 558 个组装好的转录本,其中鉴定出2 645、256、42和54 个转录本分别在4 个器官中高度表达。基于sRNA测序,鉴定出2 038、2、21和24 个sRNA分别特异性累积在4 个器官中。共检测到1 047 个成熟的候选微RNA(MicroRNA,miRNA)。基于二级结构预测和测序数据,从装配的转录本中鉴定出几十种潜在的miRNA前体。针对候选miRNA进行目标鉴定,发现1 257 个miRNA靶基因。该研究初步建立起涉及铁皮石斛生长发育、激素信号传导、次生代谢和Argonaute1(AGO1)蛋白调控相关的子网络,为之后更深层次的分子研究夯实了数据基础。
铁皮石斛作为我国重要的传统中药材,药用有效成分主要为一些植物次生代谢产物,如石斛碱(生物碱)、石斛多糖、石斛酚、氨基酸、芪类及其衍生物及挥发性成分等[72-73]。其中石斛碱是石斛类植物特有的,具有抗肿瘤、止疼、解毒、降低心率、血压和减慢呼吸、对心血管、胃肠道有抑制作用及止痛退热等作用,可产生中度的高血糖,且可解巴比妥中毒[74]。Guo等[75]对铁皮石斛茎利用 Roche 454平台进行测序,得到553 084 个ESTs。组装后获得36 407 个独特的推测转录本,共注释出69.97%的独特序列,获得了生物碱生物合成的详图。基于KEGG功能分析找到69 个独特序列,代表了涉及石斛碱骨架合成的25 个基因。实时荧光定量PCR(Real time quantitative PCR,RT-qPCR)验证发现,铁皮石斛叶片中5 个石斛碱合成关键酶基因的表达水平高于茎。筛选出细胞色素P450s、氨基转移酶、甲基转移酶、多药耐药(Multidrugresistance,MDR)转运蛋白和转录因子可能参与石斛碱的合成。研究中的39 个ABC转运蛋白转录本包括11 个MDR转运蛋白,这是首次在铁皮石斛中发现MDRs。另外,从36 407 个Unigenes中检测到1 061 个SSR,二核苷酸重复是最丰富的重复类型,其中,179 个基因与KEGG中的代谢途径有关,这为建立新的SSR标记提供丰富的DNA序列来源。
除石斛碱外,石斛多糖也是主要活性成分之一,具有抗氧化、抗炎、抗肿瘤、增强免疫功能等活性[76-77],一直是药食同源植物领域中的研究热点。Zhang等[78]为了鉴定可能与多糖合成有关的基因,从铁皮石斛幼苗期和成苗期构建了2 个cDNA文库,分别命名为石斛-1和石斛-2。2 个转录组数据库整合后获得共计145 791 个Unigenes,17 281个Unigenes注释到126 个KEGG通路上,其中135个Unigenes参与果糖和甘露糖代谢;GO分析显示大多数基因与代谢和细胞过程有关。找到430 个糖基转移酶和89 个纤维素合成酶相关基因。分析比较石斛-2与石斛-1,共发现32 794 个差异表达基因(Differential expression gene,DEGs),基因上调 22 051 个、下调10 743 个。在石斛-1和石斛-2中分别有1 142 个和7 918 个Unigenes特异性表达,这些DEGs主要与代谢途径和次生代谢产物的生物合成有关,其中170 个DEGs属于糖基转移酶基因,37个DEGs属于纤维素合酶基因,627 个DEGs编码转录因子。He等[79]利用4 个数字基因表达谱分析,对温室栽培下铁皮石斛茎中的甘露聚糖生物合成相关基因进行研究。基于甘露聚糖积累和基因表达水平,从DEGs数据库中鉴定出8 个与甘露聚糖生物合成相关的类纤维素合成酶基因(Cellulose synthaselike protein A,CSLA)。为了进一步分析这些CSLA基因,通过cDNA末端的快速扩增(Rapid-amplification of cDNA ends,RACE)获得每个基因的全长cDNA,发现这8 个基因属于CesA超家族的CSLA家族,含有CesA超家族的保守结构域,并且绝大多数基因在铁皮石斛茎中高表达,与非生物胁迫有关。表明铁皮石斛的CSLA家族基因参与生物活性甘露聚糖的生物合成。
Shen等[80]综合测定并比较了铁皮石斛不同器官中石斛碱和石斛多糖的含量。对根、茎、叶和花进行了转录组测序,基于GO和KEGG分析,确定出同参与果糖和甘露糖代谢酶有关的Unigenes以及与石斛碱生物合成途径中推测的上游元件相关的Unigenes。鉴定到大量的候选基因,包括35 个全长糖基转移酶基因和49 个全长P450基因,并确定了这些基因的器官特异性表达模式。
He等[81]对铁皮石斛中的选择性剪接异构体和有关铁皮石斛从叶片到茎的糖易位信息进行了研究。分析铁皮石斛叶和茎中的多糖含量,利用二代测序和单分子实时(Single molecule real time,SMRT)测序技术完成了这2 种组织的转录组测序,得到基因和mRNA同种型表达数据。分析比较2 个转录组共发现1 414个DEGs,其中上调844个,下调570个。在这些基因中,一种糖最终输出转运蛋白(Sugar will eventually be exported transporter,SWEET)[82]和一种糖转运蛋白(Sucrose transporters,SUT)[83]在铁皮石斛茎中表达程度比叶中高。2 种糖基转移酶和4 种纤维素合酶基因经历不同程度的选择性剪接。在茎中,多糖含量是叶中的两倍。找到另外两个基因DoSWEET4和DoSUT1在茎中显著表达,它们可能参与糖在韧皮部的装载。Wu等[84]利用转录组学和代谢组学技术,基于生理特征分析揭示在铁皮石斛幼苗生长过程中对低温驯化(Cold acclimation)的响应。这些研究极大扩充了铁皮石斛转录组信息库,为进一步筛选鉴定参与主要活性物质生物合成的候选基因,以及揭示其代谢通路与合成机制提供了可能。
4 铁皮石斛蛋白质组与代谢组
4.1 蛋白质组
赖氨酸琥珀酰化是各种真核细胞和原核细胞中普遍存在且重要的蛋白质翻译后修饰。然而,其在高多糖含量的铁皮石斛中的功能在很大程度上是未知的。Feng等[85]将液相色谱-二级质谱联用(Liquid chromatography-mass spectrometry/mass spectrometry,LC-MS/ MS)技术用于鉴定通过用高效琥珀酰赖氨酸抗体免疫纯化富集的肽。共鉴定207 种蛋白质中的314 个赖氨酸琥珀酰化位点。GO分析表明,这些蛋白质与广泛的细胞功能相关,从代谢过程到应急反应过程均有涉及。此外,还鉴定了2 种保守的琥珀酰化基序“***Ksuc******K**”和“****EKsuc***”(*表示随机氨基酸残基;Ksuc表示琥珀酰化大-K)。数据显示赖氨酸琥珀酰化发生在糖酵解途径中的5 种关键酶上,这5 种酶在植物上的平均琥珀酰化位点数目低于细菌和哺乳动物。2个活性位点氨基酸残基K103和K225可以在果糖二磷酸醛缩酶中被琥珀酰化,表明赖氨酸琥珀酰化在调节糖酵解酶活性中具有潜在功能。琥珀酰化蛋白质的相互作用网络表明,它们可能参与包含有糖酵解、三羧酸循环、氧化磷酸化和核糖体等几个功能项。这首次证明琥珀酰化参与糖酵解过程,并且可能加速琥珀酰化合成作为主要活性成分的多糖合成的未来生物学研究。
4.2 代谢组
由于铁皮石斛起作用的大多是代谢产物,因此,研究其代谢组在中药材的鉴定、质量控制上有应用意义。Jin等[86]在甲醇/水相中分离了不同生长年限的栽培铁皮石斛和霍山石斛茎中的代谢物,使用气相色谱-质谱联用(Gas chromatography-mass spectrometry,GC-MS)技术对其进行鉴定。石斛甲醇/水相中的代谢产物主要为糖类和糖苷类、氨基酸类、有机酸类和醇类。通过聚类分析,结合多因素统计分析手段,对铁皮石斛、霍山石斛以及它们不同的生长年限进行了区分。其中11 种代谢物(包括蔗糖、葡萄糖、半乳糖、琥珀酸盐、果糖,十六烷酸盐、油酰腈、肌醇和甘油等)对这种分化有显著贡献(P< 0.05)。石斛化学成分的代谢谱分析表明,霍山石斛的多糖含量高于铁皮石斛。此外,基于石斛兰代谢物的积累,石斛的最佳收获时间为第3年。
代谢组学除了可用于中药材鉴定和质量控制外,还能够探究复杂的生命过程。植物低温驯化是一种涉及基因调控和表达的基因复杂现象。铁皮石斛是一种重要的食药用植物且兼具观赏价值,对低温敏感,是研究低温驯化的理想材料。Wu等[84]利用大规模代谢组学和转录技术,基于生理特征分析揭示了铁皮石斛幼苗对低温驯化的响应。当温度由4℃降至-2℃时,24 h内抗氧化酶活性和电解质渗漏均显著升高(P< 0.01)。根据生理学分析,首次获得了0℃和20℃下20 h内的最适低温驯化点。发现铁皮石斛在低温驯化期发生大量的转录组和代谢组重编程。从基因到代谢物网络分析均表明铁皮石斛的低温驯化是一种通过激活糖酵解、氨基酸分解代谢和三羧酸循环的高能量需要过程。铁皮石斛中有2 767 个基因的表达水平受低温驯化显著影响,其中CBF转录因子上调153 倍,胞质级联蛋白激酶MAPKKK 16上调56 倍。运用低温和对照每个样品10 个平行样本的原则,采用GC-MS技术对代谢产物谱进行分析,在铁皮石斛叶片中共识别出68 个代谢物,其中33 个显示出差异性积累(27 个上升,6 个下降)。基因相互作用和调控网络分析表明,低温驯化作为一个活跃的过程,受转录、转录后、翻译和翻译后水平的严格调控。该研究提供了一个全面的调控机制,包括低温信号转导、转录调控和基因表达,这有助于深入了解铁皮石斛低温驯化期间所发生的高度复杂的调控过程。此外,铁皮石斛对于环境温度的要求较高,最适温度在25-28℃间[87],主要生长地在我国南方地区,近些年,北方地区开始在温室中栽培铁皮石斛,但成本一直较高,如果可以低温驯化铁皮石斛,提高它的抗低温性,将有利于将铁皮石斛这一资源更好的引入北方市场。
5 总结
2015年,铁皮石斛基因组得以破译,标志着铁皮石斛分子生物学时代的开始,一系列关于铁皮石斛转录组、蛋白质组和代谢组等组学方面的研究也取得了可喜的成绩。然而,还有很多问题亟待解决。例如,铁皮石斛种子萌发生长需要借助某些真菌共生[88],但现在的研究多是对于其相互作用的表型层面研究,其完整的互作机理、营养吸收和代谢机制等问题尚未明确。铁皮石斛作为珍稀濒危物种,借助全基因组数据与分子标记技术,保护、鉴定和优化种质资源,建立完善的种质资源分子遗传图谱和可追溯的原始档案方面的工作亟待加强。此外,对于利用多组学技术结合新型的分子生物学手段,发掘铁皮石斛特异和功能性基因资源的工作也需加速开展,这将帮助我们推进铁皮石斛有益成分生物合成机制及合成生物学的研究进程。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
作物学报(2022年8期)2022-05-29
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
数学大王·中高年级(2020年12期)2020-12-31
趣味(语文)(2020年4期)2020-07-27
现代园艺(2018年2期)2018-03-15
湖北农业科学(2017年7期)2017-05-13
浙江柑橘(2016年3期)2016-03-11
安徽农业科学(2015年10期)2015-02-27
