
江南油杉花粉离体萌发的适宜条件与萌发特征
2019-05-08 03:14:14
广西林业科学 2019年4期
(广西壮族自治区林业科学研究院 广西优良用材林资源培育重点实验室国家林业和草原局中南速生材繁育重点实验室,南宁 530002)
江南油杉(Keteleeria fortuneivar.cyclolepis)属松科(Pinaceae)油杉属,为我国特有常绿乔木,是山地造林、用材林和园林绿化的理想树种。现块状分布或散生于江西、湖南、广东、广西、贵州、云南、福建、浙江等地,常在海拔1 km 以下的丘陵和低山的阳坡地带生长良好[1]。每年1—2月江南油杉在多年生枝条上形成花芽,3月初花粉发育成熟并开始散粉,花粉具2个气囊,风媒传粉[2]。
花粉活力对植物果实的授粉和受精、种子产量和质量有直接影响,主要受植物自身遗传特性与外界因素的影响,尤其易受温度影响[3-5]。研究花粉的离体萌发特性,确定最佳培养基组分,可以揭示花粉的生殖细胞和营养细胞发育及花粉管的生长过程,对于植物杂交育种也具有重要的现实意义[6-8]。近年来的野外观察发现,江南油杉在传粉时极易受雨季影响,致使大量球果授粉不足,进而导致种子败育率高、质量较低。本研究通过花粉离体萌发对江南油杉的花粉萌发特性进行研究,对其花粉活力进行快速检测,并了解其花粉管生长发育过程,旨在找出适合江南油杉花粉萌发的最佳培养基配方,为解决球果授粉不足问题及开展种质资源保存与人工授粉提供基础数据,也为裸子植物孢粉学研究积累有用的资料。
1 材料与方法
1.1 试验材料
试验所需的江南油杉花粉采自广西南丹县里湖乡麻风寨(107°28'E,25°07'N)保存良好的天然林。于2018年3月初,采集发育正常尚未散粉的江南油杉雄球花,带回实验室后用镊子剥下小孢子叶球,放入铺有硫酸纸的培养皿中,置于25℃培养箱内散粉24 h,后收集花粉,置于-20℃冰箱保存备用。
1.2 试验方法
1.2.1 花粉的离体萌发
采用液体培养法对江南油杉花粉进行离体萌发试验,通过单因素试验设计分别对培养基蔗糖浓度、硼酸(H3BO3)浓度、氯化钙(CaCl2)浓度、pH 值以及培养温度进行研究。其中,蔗糖浓度试验的培养基为150 mg/L H3BO3+ 150 mg/L CaCl2+ pH 6.5,并分别添加0、50、100、150、200、250、300、350和400 g/L 蔗糖,共9个处理。H3BO3浓度试验的培养基为最适蔗糖浓度+150 mg/L CaCl2+ pH 6.5,并分别 添 加0、50、100、150、200、250和300 mg/L H3BO3,共7个处理。CaCl2浓度试验的培养基为最适蔗糖浓度+最适H3BO3浓度+ pH 6.5,并分别添加0、50、100、150、200、250和300 mg/L CaCl2,共7个处理。pH 值试验的培养基为最适蔗糖浓度+最适H3BO3浓度+最适CaCl2浓度,pH 值分别设置为5.0、5.5、6.0、6.5和7.0,共5个处理。培养基试验均在25℃条件下100 r/min 振荡光照培养。以筛选获得的最佳组分配制培养基,分别设置15、20、25、30和35℃培养温度,共5个处理,100 r/min 振荡光照培养。所有试验总计33个处理,每处理称取5 mg 江南油杉花粉置于1 mL 培养基中,重复3次。
1.2.2 花粉萌发率与花粉管长度观测
江南油杉花粉在吸水24 h 后才能逐渐萌发,因此,各处理分别培养48、72和96 h后从培养基取样,在OlympusCX-41 型光学显微镜下观测记录,每处理的每个重复随机观察5个视野,计算花粉萌发率,花粉管长度大于花粉粒体宽视为萌发。花粉萌发率=(萌发花粉数/花粉粒总数)× 100%。随机选取萌发的花粉测量花粉管长度,每处理的每个重复随机测量30个花粉管长度。
1.3 数据处理
采用Excel 2016对花粉萌发率和花粉管长度进行常规统计和制图。采用SPSS 19.0对各试验处理间花粉萌发率和花粉管长度进行方差分析和多重比较(邓肯氏新复极差法)。
2 结果与分析
2.1 蔗糖浓度对花粉离体萌发的影响
不同蔗糖浓度处理,江南油杉花粉萌发率和花粉管长度存在极显著差异(P<0.01)。在无蔗糖的培养基(0 g/L)中,虽然花粉也可萌发,但萌发率显著低于100~300 g/L 浓度下花粉的萌发率(P<0.05),花粉管长度显著低于50~300 g/L 浓度下花粉管长度(P<0.05)(表1)。3个时段,花粉萌发率和花粉管长度均随蔗糖浓度的增加呈先增加后减小的趋势;培养48 h,花粉萌发率在250 g/L 浓度下达到最大(17.5%),花粉管长度在200 g/L 浓度下最长(89.4 μm);培养72和96 h,花粉萌发率和花粉管长度均在250 g/L 浓度下达到最大,分别为24.4%、141.3 μm和32.2%、210.8 μm。250 g/L 蔗糖较适合江南油杉花粉萌发。
2.2 H3BO3浓度对花粉离体萌发的影响
不同H3BO3浓度处理,江南油杉花粉萌发率和花粉管长度存在极显著差异(P<0.01)。3个时段,花粉萌发率和花粉管长度均随H3BO3浓度的增加呈先增加后减小的趋势;培养48 h,花粉萌发率和花粉管长度均在200 mg/L 浓度下达到高峰,分别为14.8%和82.9 μm;培养72和96 h,花粉萌发率均在200 mg/L 浓度下达到高峰,分别为26.8%和36.1%,花粉管长度均在250 mg/L 浓度下达到高峰,分别为150.4和218.4 μm,但与200 mg/L 浓度下花粉管长度差异不显著(表2)。200 mg/L H3BO3较适合江南油杉花粉萌发。
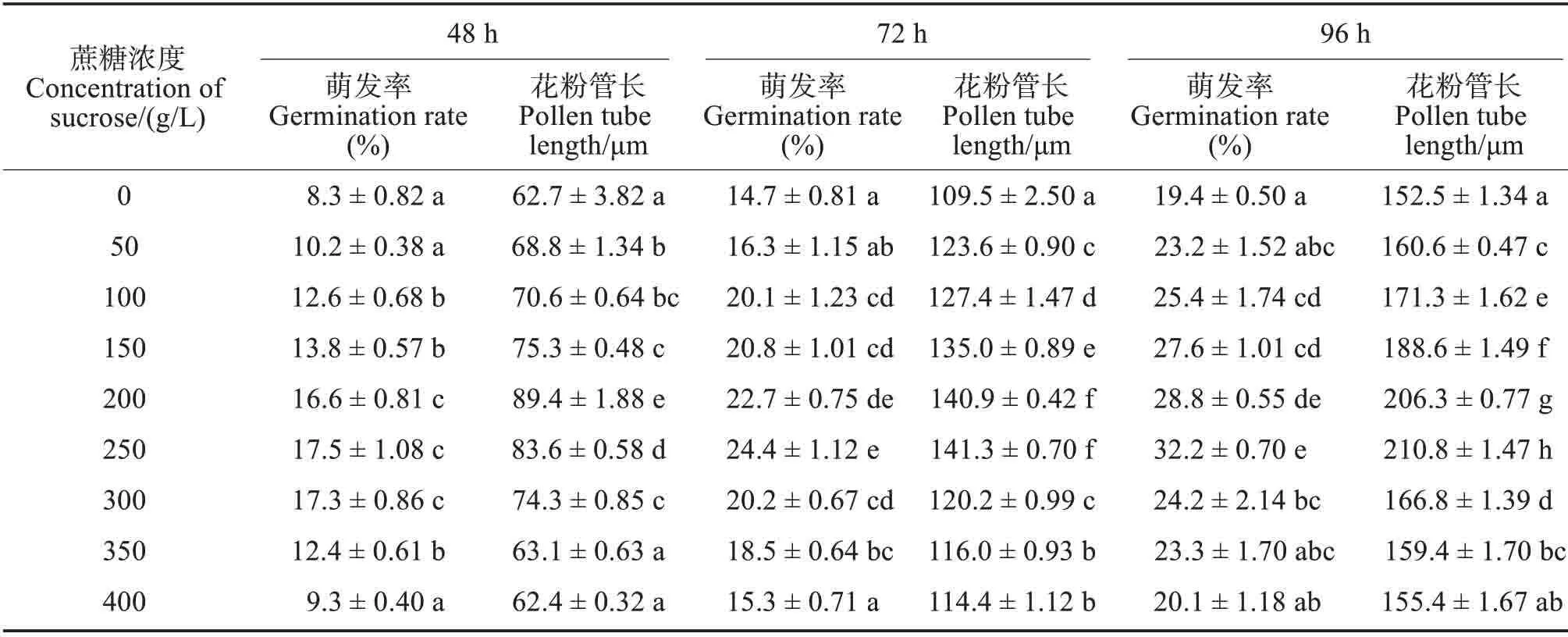
表1 蔗糖浓度对江南油杉花粉萌发率和花粉管长度的影响Tab.1 Effects of sucrose concentrations on pollen germination rate and pollen tube length of Keteleeria fortunei var.cyclolepis
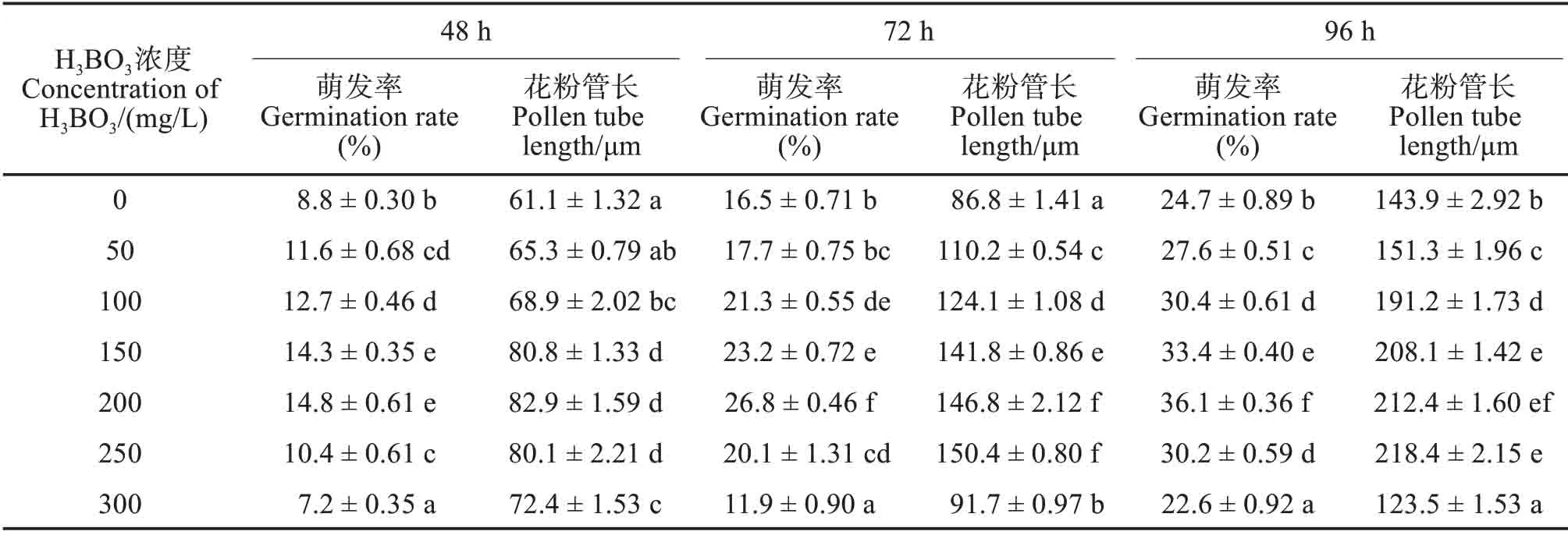
表2 H3BO3浓度对江南油杉花粉萌发率和花粉管长度的影响Tab.2 Effects of H3BO3 concentrations on pollen germination rate and pollen tube length of Keteleeria fortunei var.cyclolepis
2.3 CaCl2浓度对花粉离体萌发的影响
3个时段,江南油杉花粉萌发率和花粉管长度均随CaCl2浓度的增加呈先增加后减小的趋势;培养48 h,花粉萌发率和花粉管长度均在100 mg/L浓度下达到高峰,分别为18.7%和81.9 μm;培养72和96 h,花粉萌发率均在100 mg/L 浓度下达到高峰,分别为32.3%和41.3%,花粉管长度均在150 mg/L浓度达到高峰,分别为151.4和215.3 μm,但与100 mg/L 浓度下花粉管长度差异不显著(表3)。100 mg/L CaCl2较适合江南油杉花粉萌发。

表3 CaCl2浓度对江南油杉花粉萌发率和花粉管长度的影响Tab.3 Effects of CaCl2 concentrations on pollen germination rate and pollen tube length of Keteleeria fortunei var.cyclolepis
2.4 pH值对花粉离体萌发的影响
当培养基pH 值为5.0和5.5 时,江南油杉花粉萌发率较低,两者差异不显著,花粉管生长缓慢,随着培养基pH 值升高,花粉萌发率和花粉管长度也随之增加;当pH 值为6.5 时,3个时段花粉萌发率和花粉管长度均达到最大,之后降低(表4)。培养基pH 6.5 较适合江南油杉花粉萌发。

表4 pH值对江南油杉花粉萌发率和花粉管长度的影响Tab.4 Effects of pH values on pollen germination rate and pollen tube length of Keteleeria fortunei var.cyclolepis
2.5 温度对花粉离体萌发的影响
不同温度处理下江南油杉花粉萌发率和花粉管长度差异显著(P<0.05)。在15℃处理下,花粉萌发率较低,花粉管生长缓慢;随着温度升高,花粉萌发率迅速升高,3个时段均在25℃最高,分别为18.5%(48 h)、33.2%(72 h)和42.1%(96 h)(图1a)。3个时段花粉管长度随温度的变化规律不一致;培养48 h,花粉管长度随温度升高逐渐增加;培养72 h,花粉管长度随温度升高先增加后减小,在30℃达到最大(156.24 μm),但与25℃花粉管长度差异不显著;培养96 h,花粉管长度随温度升高先增加后减小,在25℃达到最大(217.44 μm)(图1b)。
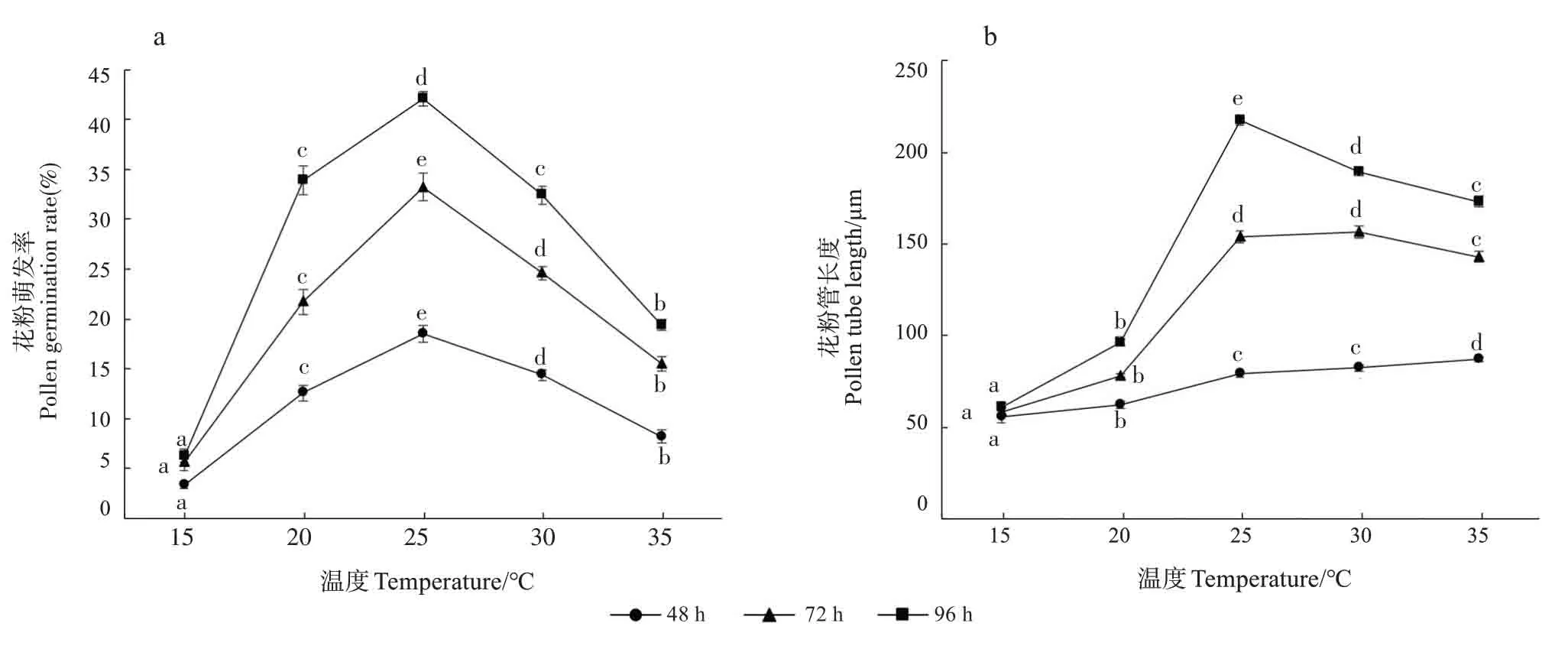
图1 温度对江南油杉花粉萌发率和花粉管长度的影响Fig.1 Effects of temperatures on pollen germination rate and pollen tube length of Keteleeria fortunei var.cyclolepis
3 结论与讨论
大量研究表明,在花粉离体萌发培养基中蔗糖、H3BO3和Ca2+为最常用的添加物质,其中,蔗糖既是花粉萌发和花粉管壁合成的营养物质来源,又是花粉萌发时内部代谢与跨膜运输的碳源,维持着外界环境的渗透压和花粉萌发形态[6,9]。Dane 等[10]研究发现花粉在离体萌发时,其自身储存的营养物质调控着外界的蔗糖浓度。本研究结果表明,在无蔗糖培养基中江南油杉花粉仍可萌发,但萌发率和花粉管长度显著低于添加蔗糖的培养基,可能是花粉自身储存的营养物质为其萌发提供一定养分,但随着营养物质逐渐消耗,花粉管生长缓慢;添加不同浓度蔗糖的培养基之间,花粉萌发率和花粉管长度差异显著,高浓度蔗糖会抑制花粉萌发和花粉管生长,这与田菊等[11]对油松(Pinus tabuliformis)花粉离体萌发和李国平等[12]对黑松(Pinus thunbergii)花粉离体萌发的研究结果一致。
花粉离体萌发培养基中的H3BO3可引导培养基内的Ca2+进入花粉细胞内,并增加花粉细胞对糖的吸收与代谢,促进花粉萌发和花粉管生长[13-14]。Ca2+对花粉管极性生长与生长方向具有调控作用[15]。相关研究表明,H3BO3浓度和Ca2+浓度对油松[11]、黑松[12]、五针松(Pinus parviflora)、罗汉松(Podocarpus macro⁃phyllus)[16]和银杏(Ginkgo biloba)[17]等裸子植物离体花粉的萌发率和花粉管长度均表现出先促进后抑制的作用,高浓度硼酸和Ca2+会抑制花粉的萌发,与本文研究结果一致。但本研究中,培养48 h,花粉管长度在200 mg/L H3BO3和100 mg/L CaCl2处理下达到高峰,而培养72和96 h,花粉管长度均在250 mg/L H3BO3和150 mg/L CaCl2处理下达到高峰,这可能是因为在培养前期,200 mg/L H3BO3和100 mg/L CaCl2能满足花粉管生长的需求,随着培养时间增加需要更高的H3BO3和CaCl2浓度才能促进花粉管生长。
不同植物花粉离体培养基最佳pH 值和最适培养温度差异较大。赵丽娟等[6]研究表明小黑杨(Pop⁃ulus simonii×P.nigra)花粉离体萌发的最佳pH 值为6.0,最适培养温度为21℃;黄家兴等[18]研究表明凯特杏(Prunus armeniaca‘Katy’)花粉离体萌发的最佳pH值为6.0,最适培养温度为20℃;Kakani等[19]研究表明陆地棉(Gossypium hirsutum)花粉离体萌发的最适温度为32℃。试验表明,pH 值和培养温度均对江南油杉花粉萌发影响显著。江南油杉花粉离体培养基最适pH值为6.5,当pH值为7.0时,花粉萌发率下降,当pH 值为5.0 时,花粉萌发率极低,且花粉管生长缓慢,说明江南油杉花粉萌发需要弱酸环境,强酸可能会破坏花粉细胞内部代谢平衡[20]。江南油杉花粉萌发的最适温度为25℃,过高或过低均会抑制其花粉萌发,25℃与江南油杉散粉期外界温度相近,说明温度并不是自然条件下江南油杉球果授粉不足的限制因素。
综上所述,江南油杉花粉离体萌发最佳培养基组合和培养条件为250 g/L 蔗糖+ 200 mg/L H3BO3+100 mg/L CaCl2,pH值为6.5,置于25℃光照培养96 h时萌发率为42.4%,为进一步开展江南油杉种质资源保存和人工授粉的研究提供基础数据。
猜你喜欢
西部林业科学(2023年4期)2023-09-02 07:23:28
中国果业信息(2022年1期)2022-11-28 07:43:38
三农资讯半月报(2020年10期)2020-06-08 15:32:59
山东林业科技(2019年2期)2019-06-03 10:10:54
生物学杂志(2018年4期)2018-08-15 11:21:32
现代园艺(2017年11期)2017-06-28 11:32:46
中国果业信息(2017年8期)2017-01-13 20:06:36
奥秘(2016年10期)2016-12-17 13:13:11
广西林业科学(2016年3期)2016-03-16 05:43:42
广西林业科学(2016年3期)2016-03-16 05:43:40
