
西南红山茶花表型性状的变异
2019-05-05 01:46李芳,霍达,王进
贵州农业科学 2019年4期
李 芳, 霍 达, 王 进
(贵州省林业科学研究院, 贵州 贵阳 550005)
西南红山茶(Camelliapitardii)是山茶科山茶属植物,是高原山地优质木本油料树种。生于海拔1 000~2 400 m的林下、林缘或灌丛中。在贵州西部至中部、四川西南部至南部、云南南部至东南部、广西北部、湖南西部和湖北西部均有分布。西南红山茶树型美观,花朵美丽,不仅是油料树种,也是较好的观赏树种,相关研究主要涉及西南红山茶的种质资源开发与利用[1]、果实性状与含油量分析[2-5]、生长环境和植物生长调节物质对其生长发育的影响[6-13]等。西南红山茶野生群体中存在多样的花部形态和丰富的花色变异,具有很好的观赏性,是一种具有开发潜力的观赏植物。目前对该物种花表型多样性性状变异的研究鲜见报道。鉴于此,2018年3月对贵州省六盘水市、毕节市、贵阳市和黔南布依族苗族自治州等地现有分布的西南红山茶林分中的西南红山茶单株花性状进行调查,对9个野生居群西南红山茶单株花的表型多样性进行分析,以揭示其多样性,拟为西南红山茶野生种质资源的保护、引种及优良品种的选育提供科学参考依据。
1材料与方法
1.1样本来源
2018年3月对贵州六盘水市盘县老厂林场(P1)、大山镇(P2)、松河镇(P2)、柏果镇(P3)和淤泥乡八大山(P4),毕节市赫章县水塘堡乡(P5)和平山乡(P6)、七星关区阳山公园(P7),贵阳市清镇市九龙山(P8),及黔南布依族苗族自治州贵定县云雾镇(P9)等野生居群中西南红山茶分布区进行调查,每个居群内随机选取2~7株长势正常、无明显缺陷和病虫害的个体,株间距离大于30 m,并进行挂牌标记。各采样点的单株编号、地理位置等基本情况见表1。

表1 9个野生居群西南红山茶样本采集编号及地理位置
1.2方法
1.2.1表型性状的测定用中国传统色彩表示法标注花颜色,参照张海平等[14]的方法对所测得的花瓣颜色进行赋值。每株随机选择10朵花,用电子游标卡尺测量花直径(测量精度为 0.1 mm),肉眼观测花瓣数、萼片数、柱头开裂数、雌雄相对高度、萼片颜色(绿色1)、萼片绒毛(无绒毛0,有绒毛1)、子房绒毛(有绒毛1)、花朵香味(无香味0)、柱头裂位(高裂位1,浅裂位2)、雌雄蕊相对高度(雌高1,雄高2,雌雄等高3)和花瓣颜色(水红1,粉红2,浅粉色3,玫瑰红4,胡粉色5,粉白6,茶白7,鱼肚白8,藕合色9);每株随机选择10片叶子观测其叶长、叶宽和叶脉数。
1.2.2表型性状的计算采用方差分析、多重比较、表型分化系数等方法计算种群间表型性状变异特征[15];以平均值和标准差计算变异系数(CV=标准差/平均值×100%),用以衡量表型性状种群内变异水平,变异系数越大,说明该种群的性状离散程度越大,表型多样性越高,反之则性状离散程度越低[16];采用相关分析得出地理生态因子对表型性状的影响;计算各种群表型性状的平均值;采用Shannon-Weiner多样性指数评价花色丰富度和均匀度[17];采用欧氏平均距离计算各种群遗传距离。
1.2.3数据统计与分析采用SPSS 19.0及 Excel 2010等软件进行统计分析。
2结果与分析
2.1不同居群西南红山茶花表型变异特征
2.1.1花表型性状的表现贵州9个野生居群红山茶花的萼片色均为绿色,子房均有绒毛,花朵均无香味。从表2可知,柱头开裂数P4居群与其他8个居群间差异显著,柱头裂位P1居群与其他8个居群间差异显著,其余各性状在居群间均存在显著差异。P8居群红山茶的花瓣数、花径、萼片数、叶脉数、叶长和叶宽均最大。

2.1.2花表型性状的变异系数从表3看出,西南红山茶各性状的平均变异系数为13.73%,变异幅度为0~51.88%,其中萼片色、子房绒毛和花朵香味的变异系数均为0;花色的变异系数最大,为51.88%,萼片绒毛和雌雄蕊相对高度的变异系数较大,叶脉数、柱头裂位和柱头开裂数的变异系数较小。说明,西南红山茶的萼片色、子房绒毛和花朵香味未发生变异;叶脉数、柱头裂位和柱头开裂数的变异幅度较小,稳定性高;花色、萼片绒毛和雌雄蕊相对高度变异较大,稳定性差。9个野生居群15个性状的平均变异系数为5.89%~24.87%,P3居群的变异系数最小,为5.89%,其表型多样性程度较低;P1居群的变异系数最大,其表型多样性较丰富。

表39个野生居群西南红山茶花表型性状的变异系数
2.2西南红山茶花色性状的多样性
根据中国传统色彩表示法,将9个野生居群36个单株360朵西南红山茶花的花色分为水红、粉红、浅粉色、玫瑰红、胡粉色、粉白、茶白、鱼肚白和藕合色9类,西南红山茶的花色呈多样化,花色频率分布以水红为主,占50%,其次是茶白、浅粉色和粉红,分别占16.66%、11.11%和8.33%,玫瑰红、胡粉色、粉白、鱼肚白和藕合色各占2.78%。西南红山茶花色的多样性指数为1.594,说明西南红山茶花色多样,丰富度较大。
2.3西南红山茶花性状的相关性
从表4看出,西南红山茶花的15个性状中除花朵香味、子房绒毛和萼片色因无变异为常量不作相关性分析外,其余12个花性状间存在不同程度的相关性,相关系数为-0.581~0.742,其中,花瓣数与花色、萼片数相关性不显著,与叶宽呈显著负相关,与雌雄蕊相对高度极显著负相关,与其余各性状间均呈极显著正相关;花径与叶长、叶宽和叶脉数间呈极显著正相关,与雌雄蕊高度呈极显著负相关,与其余各性状间相关性不显著;花色除与叶长不相关外,与柱头裂位、雌雄蕊相对高度、萼片绒毛、叶长宽比间呈极显著正相关,与柱头开裂数、萼片数、叶脉数和叶宽间呈极显著负相关;柱头开裂数与叶长、叶宽和萼片数呈极显著负相关,与萼片绒毛间呈极显著正相关;柱头裂位与雌雄蕊相对高度、萼片绒毛、叶脉数和叶长宽比间均呈极显著正相关,与叶宽间呈极显著负相关;雌雄蕊相对高度与萼片数、叶长和叶宽间均呈极显著负相关;萼片绒毛与叶长和叶长宽比间均呈极显著正相关;叶脉数与叶长和叶宽间均呈极显著正相关;叶长与叶宽间呈极显著正相关;叶宽与叶长宽比间呈极显著负相关。

表49个野生居群西南红山茶花表型性状间的相关性
注:*和**表示两性状间分别呈显著和极显著相关水平(下同)。
Note:* and**indicate significant difference at 1% and 5% levels. The same below.
从表5可知,花径、柱头裂位、萼片绒毛、叶脉数、叶长及叶宽均与经度、纬度和海拔间均呈极显著负相关;雌雄蕊相对高度与经度、纬度和海拔间均呈极显著正相关;花瓣数、萼片数和叶长宽比与海拔间均呈极显著负相关,柱头开裂数与海拔间呈极显著正相关,与经度和纬度间的相关性均不显著;花色与经度、纬度和海拔间相关不显著。

表59个野生居群西南红山茶花表型性状与地理生态因子间的相关性
2.4西南红山茶花表型性状的聚类
从图1看出,在欧氏距离20处可将36个西南红山茶单株划分为3个大类,第Ⅰ类包括27、28、10、……和8等29个单株;第Ⅱ类仅有2个单株(17和18);第Ⅲ类包括5个单株(32、34、33、35和36)。在欧氏距离10的阀值处,第Ⅰ类又可分了10个亚类,第Ⅲ类可分为2个亚类。总体看,36个单株的1~31号单株并没有依据居群划分进行聚类,不同居群的西南红山茶单株分散在各大类中,不存在区域上的特异性,32~36号的P8和P9居群可能由于经纬度和海拔明显不同于其他居群,所以欧氏距离25处就与其他单株不同类。
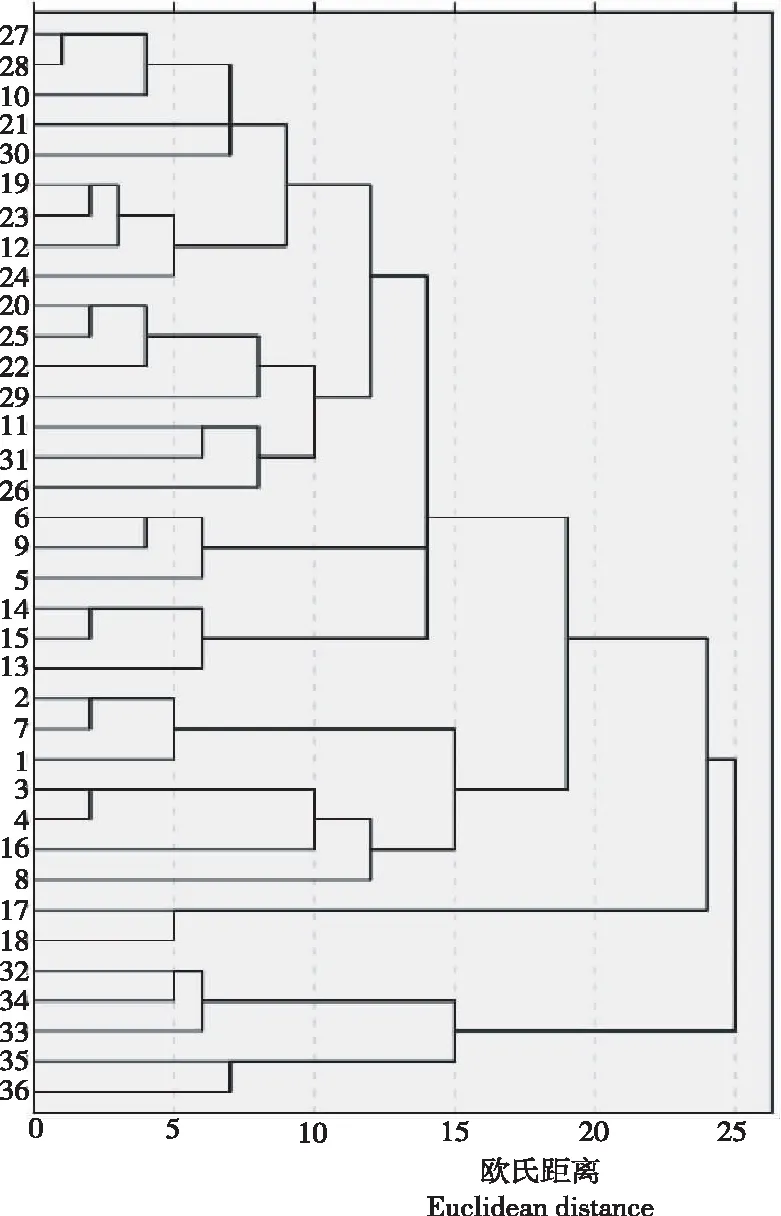
图19个野生居群西南红山茶36个单株的欧氏距离聚类情况
3结论与讨论
表型多样性是遗传多样性与环境多样性的综合体现,既具有稳定性,又具有变异性,是遗传多样性研究的重要内容[18],利用表型性状研究植物的遗传变异是最直接、简便易行的方法,因此通常依据植物表型性状进行种质资源的分类、鉴定及育种材料选择[19]。对9个野生居群西南红山茶36个单株的15个表型性状的变异分析发现,其花的性状变异丰富,15个性状的平均变异系数为13.73%。其中,P1居群的变异系数(24.87%)最高,P3居群的变异系数(5.89%)最低。9个野生居群不同性状间呈不同程度的差异性。
从西南红山茶花颜色的分布频率和多样性指数看,西南红山茶花色呈多样化,其中以水红为主,占50%,其次是茶白、浅粉色和粉红,分别占16.66%、11.11%和8.33%,玫瑰红、胡粉色、粉白、鱼肚白和藕合色各占2.78%。花色多样性指数为1.594,说明西南红山茶花色多样,丰富度较高。
9个野生居群西南红山茶36个单株花性状与经度、纬度和海拔均有不同程度的相关性。聚类结果显示,36个单株的1~31号单株并没有依据居群划分进行聚类,不同居群的西南红山茶单株分散在各大类中,不存在区域上的特异性,32~36号的P8和P9居群可能由于经纬度和海拔明显不同于其他居群,所以在欧氏距离25处就与其他单株不同类。
目前,西南红山茶多处于野生状态,资源丰富、天然杂交的自然变异形成了多种变异类型,广泛的变异和丰富的遗传基础为西南红山茶遗传改良提供了丰富的材料。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
广西植物(2020年13期)2020-09-12
广西植物(2020年13期)2020-09-12
广东园林(2020年3期)2020-07-03
文苑(2020年6期)2020-06-22
大观(书画家)(2018年6期)2018-07-08
花卉(2015年11期)2015-09-14
植物资源与环境学报(2012年2期)2012-09-06
