
广西海岸带天然林群落多样性及种间联结性
2019-04-26 00:39:26张艺帆谢伟东邓荣艳单笑旋
广西林业科学 2019年1期
张艺帆,谢伟东,邓荣艳,单笑旋
(广西大学林学院,南宁 530004)
海岸带作为海陆之间相互作用的地带,承接着联系人类与海洋生态系统的作用,是一个能量与物质激烈交换的地区[1]。同时,海岸带又是生态脆弱、自然灾害多发的地带[2]。其生态环境优美、自然资源丰富,但滨海旅游、城市、工业、港口、农业和海水养殖等大规模开发利用,导致自然植被的破坏或退化严重,由此引发了一系列生态问题,海岸带植被的恢复应得到重视[3]。广西防城港目前尚存少量保存较为完整的地带性原生滨海植被,近年来,对该地区的研究主要集中在红鳞蒲桃(Syzygium hancei)群落资源分布和群落特征以及土壤理化性质的研究[4-6],开花物候特征略有涉猎[7-9],物种多样性的研究较少[5-6,10-11],种间联结性研究尚未见报道。这些研究仍不够系统深入,植被群落结构特征仍不明确,物种多样性的变化规律及其驱动机制还待探明。在东兴市竹山村天然林群落开展物种多样性以及种间联结的研究,有助于了解滨海植被群落结构与功能以及群落的演替趋势,对滨海植被保护和恢复重建提供理论依据。
1 材料与方法
1.1 研究地概况
研究地位于广西防城港东兴市竹山村(107°53′~108°15′E,21°31′~21°44′N,),该群落面积约为5.56 hm2,沿潮上带边缘呈带状分布,宽度40~60 m,长度约1 400 m。该区域为滨海平原,地势平坦,海拔一般在10 m以下,地带性土壤为赤红壤和砖红壤,属南亚热带季风气候区,全年气候温和湿润,雨热同期,年均气温23.2℃左右,年日照时数在1 500 h以上,雨量充足,年均降水量达到2 738 mm。
1.2 样地设置与调查
本群落为当地“风水林”得以保存下来,被粉单竹(Bambusa chungii)侵蚀严重,根据群落大小分布,按照相邻格子法设置9个20 m×20 m的大样方,再将每个大样方分为4个10 m×10 m的小样方,记录乔木(胸径≥1 cm)的种类、高度、枝下高、胸径和冠幅等信息。在每个大样方按对角线设置5个5 m×5 m小样方进行灌草调查,记录灌木(胸径<1 cm)种类、株数、平均高度、盖度等;草本层的种类、株数、平均高度、盖度。并调查乔木层的郁闭度及灌草层的总覆盖度。
1.3 多样性的测度
1.3.1 物种重要值
重要值(IV)=相对多度+相对频度+相对优势度
1.3.2 物种多样性指数
采用物种丰富度指数(S)、Shannon-Wiener指数(H′)、Simpson指数(D)、Pielou均匀度指数(Jsw)[12-14]
S=n
H′=-∑PilnPi
D=1-∑Pi2
Jsw=H′/lnS
式中:n为出现在样方中的物种数,Pi为种i的相对重要值。
1.4 物种联结性测定
1.4.1 总体关联性测定
Schluter[15]提出的方差比率法检验多物种间的关联性,计算公式为

式中:S为总的物种数;N为总的样方数;Tj为样方j内出现的物种数;ni为物种i出现的样方数;t为样方中种的平均数。
VR=1,复合种间关联性是独立的;VR>1,多物种间表现为正的相关;VR<1,多物种间表现为负的相关。
为检验相关性的显著性,采用统计量W(W=N×VR)进行χ2检验,如果种间独立,则W落入χ20.95(N)<W<χ20.05(N)的概率有 90%[16]。
1.4.2 成对物种关联性检验
基于样方调查中所获得的数据,建立2×2列联表,计算其中的a(包含两个种的样方数)、b(只存在B种的样方数)、c(只存在A种的样方数)、d(两个种都不存在的样方数)值,由于是不连续取样,一般使用Yates的连续校正系数来纠正[8]。

式中,n为小样方总数,χ2符合自由度为1的分布,根据χ2(0.05)、χ2(0.1)的理论值,和计算得χ2实测值对照,得出当χ2<3.841时,种间联结不显著;当3.841≤χ2<6.635时,种间联结显著;当χ2≥6.635时,种间联结极显著。χ2本身没有负值,判定正、负联结的方法是当ad>bc时,种间具正关联;若ad<bc,种间具负关联。
1.4.3 物种共同出现百分率PC[17]
PC=a/(a+b+c)
1.4.4 种间关联系数AC[17]
AC=(ad-bc)/(a+b)(b+d)(ad>bc)
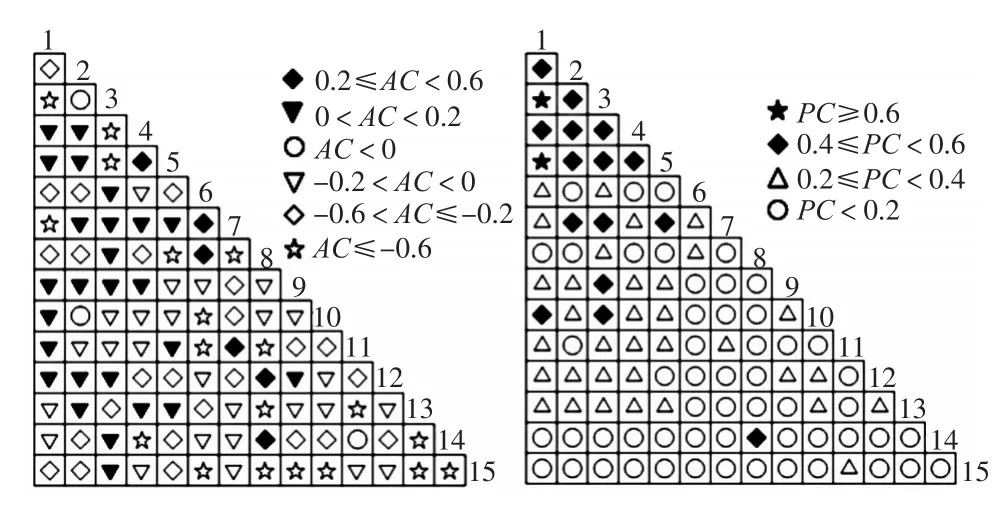
AC=(ad-bc)/(a+b)(a+c)(ad AC=(ad-bc)/(d+b)(d+c)(ad<bc,d 经调查统计,群落内共有植物84种,分属41科70属,其中乔木层树种25科34属40种,灌木层树种30科54属66种(包括乔木幼苗幼树20科25属30种),草本层植物8科8属8种。该群落分层比较明显,大致可分为乔木层、灌木层、草本层。对群落内的植物的重要值进行统计计算,列出每个层重要值在5%以上的主要种(表1)。 表1 主要物种组成及重要值Tab.1 Important values of major plant species 结果表明,群落乔木层物种组成较丰富,优势种是粉单竹,重要值为44.97%,假苹婆和红鳞蒲桃是次优势植物,重要值分别为和27.73%和27.33%,重要值大于10%的树种有8种。灌木层中重要值排在前5位的依次是罗伞树、细枝龙血树、南山花、鸭脚木和龙船花,其中以罗伞树和细枝龙血树为共优势种,重要值分别为50.93%和43.74%,重要值大于10%的树种有7种,其中罗伞树、鸭脚木和滨木患是乔木层的优势树种的幼树,说明该群落在演替过程中具备较强的天然更新能力。草本层种类和数量都很少,处于退化状态,其中华山姜占绝对优势,重要值达到186.36%,沿阶草是次优势种,重要值为72.24%,其余物种重要值大小分布较均匀,彼此差异不大。 群落内物种多样性表现为灌木层>乔木层>草本层;群落内乔木层和灌木层的S、H′、D、Jsw都处于较高的水平,灌木层丰富度最高,且多为乔木树种的幼树,数量多密度大,种群呈现出较强的更新能力;草本层的各多样性指数水平均较低,呈零星分布,多生长在群落边缘(表2)。 表2 群落内物种多样性Tab.2 Species diversity in the community 乔木层的方差比率VR=0.396<1,总体上种间存在一定的负联结,检验统计量W=14.257,落在χ20.95(36)与χ20.05(36)之外,表明乔木层优势树种间总体上表现为显著的负联结关系;灌木层的VR=2.070>1,表明总体上种间存在一定的正联结,W值为 74.515,落在χ20.95(36)与χ20.05(36)区间之外,表明灌木层优势树种总体上的正联结关系达到显著水平(表3)。 3.4.1 乔木层优势种的联结性 乔木层优势种群的χ2检验数据和连接系数AC、共同出现率PC半矩阵图如表4和图1所示。 χ2统计量显示,乔木层主要优势树种种对间的联结关系整体都不显著,仅有3个种对达到显著水平,占2.8%,且都为负相关,其余种对联结均不显著。根据图1的AC值可以看出,在15个树种组成的105个种对间正联结种对数为32个,占总种对数的30.5%;负联结种对数为70个,占66.7%;3个种对表现为相互独立(AC=0),占2.8%。负联结的数量和强度均大于正联结,正联结强度大都不超过0.2,整体上联结强度值AC处于-0.2~0.2之间的有51对,占总种对数的48.6%。从PC值结果看,PC值大于0.6的仅有2对,其余大多数都表现为弱联结性。 表3 种群间的总体联结性Tab.3 Overall association between populations 表4 乔木层主要优势树种χ2统计量半矩阵Tab.4 Semi-matrix of χ2value of dominant tree populations 图1 乔木层优势种群种间联结AC值和PC值半矩阵图(种序号同表4)Fig.1 Semi-matrix of AC and PC value of dominant tree populations(The species No.is the same as Tab.4) 显著负联结的种对有罗伞树—海杧果、罗伞树—银柴、密花树—滨木患,联结性较强,共同出现率也较低(PC<0.2),在群落中形成各自种群,占据不同的生态位。假苹婆与鸭脚木不显著负联结,联结系数表现为较强的负联结(AC≤-0.6),而共同出现率较高(PC≥0.6),假苹婆是阳性树种,鸭脚木是中性偏阳种,可以在相同的生境条件下较好地利用不同的资源。假苹婆和罗伞树的共同出现率很大(PC≥0.6),但表现为不显著的正联结和弱的联结强度(0<AC<0.2),呈现上下层的共生关系。 3.4.2 灌木层优势种的联结性 对灌木层优势树种进行χ2检验以及共同出现率PC和联结强度AC的测算(表5和图2)。χ2统计量显示,灌木层12个优势树种组成的66个种对间,南山花和鸭脚木之间存在极显著的正联结,南山花和细枝龙血树、九节以及肉实树3个种对达到显著正联结水平,罗伞树和滨木患之间表现为显著负联结。根据图2的AC值可以看出,正联结种对数为41对,负联结种对数为23对,滨木患—肉实树、筐条菝葜—假鹰爪、筐条菝葜—棕竹3个种对相互独立(AC=0)。整体上联结强度值AC处于-0.2~0.2之间的有29对,占总种对数的43.9%,联结较松散。 表5 灌木层主要优势树种χ2统计量半矩阵Tab.5 Semi-matrix of χ2value of dominant shrub populations 图2 灌木层优势种群种间联结AC值和PC值半矩阵图(种序号同表5)Fig.2 Semi-matrix of AC and PC value of dominant shrub populations(The species No.is the same as Tab.5) χ2检验结合连接系数AC值和物种共同出现率PC值分析,在灌木层的66个种对中,表现为正联结的有41对(占62.1%),负联结23对(占34.8%),无联结种对3对(占4.5%)。正联结中,极显著正联结的1对(占1.5%),显著正联结的有3对(占4.5%),不显著的有37对(占56.1%);负联结中,显著负联结的有1对(占1.5%)不显著的有22对(占33.3%)。联结种对没有达到显著水平的有59对,占总种对的89.3%,说明大多数种对的联结没有达到显著水平,种间联结较松散,基本趋于独立分布。 广西东兴市竹山村滨海植物群落S、H′和D均表现为灌木层>乔木层>草本层,这与林分郁闭度较高和上层植物分布不均匀有关;Jsw总体表现乔木层最高,灌木层次之,草本层最低。随着S的升高,H′,D和Jsw也随之升高。造成该群落层次多样性格局形成的原因可能由于在乔木层中红鳞蒲桃占据统治地位,其他树种相对比较固定,而灌木层除了灌木树种外,还包括了绝大多数乔木树种的幼苗和幼树,这都造成灌木层的种类和数量多于乔木层,从而具有较高的物种多样性,由于上层树种的盖度较大,遮蔽了阳光,所以林下草本数量和种类都很稀少。 群落的种间关联性与群落的稳定性密切相关,种间总体上正关联能够反映出群落具有较强的稳定性[18-20]。本文的结果与广东黑石顶自然保护区阳生性常绿阔叶林阶段[21]以及水口山国有林场马尾松群落[22]相似。广西东兴市竹山村滨海天然林群落总体上表现为较多的负关联,显著率较低,种间联结较松散。乔木层主要种群总体上表现为显著负关联,正负关联比<1,且多数物种间的联结性未达到显著水平,说明乔木层不稳定,种间竞争比较激烈;灌木层则表现为显著正关联,正负关联比>1,且主要优势种群南山花和鸭脚木之间正相关达到极显著水平,和细枝龙血树、九节以及肉实树之间达到显著水平,说明灌木层具有一定的稳定性,优势种之间搭配合理。 多个研究表明,呈现正联结的物种对生境具有相近的适应性,两个物种间的正联结强度越高,表明它们之间的生境需求越相似,生态位重叠越高[21,23-25];或是一个种对另一个种的定居提供了条件[26],负联结表明两物种间生境需要有明显差异或者两者之间存在激烈的竞争关系[27]。本文的研究也证明了这一点,乔木层中罗伞树和海杧果、罗伞树和银柴、密花树和滨木患均表现为显著的负联结,它们的生态位重叠较小,可能它们对于生境需求的适应性不同或是相互分离的生态位所致,在群落中分布相互独立,如海杧果、银柴和滨木患为喜光阳性植物,而罗伞树和密花树喜温暖湿润气候,耐荫性较强;灌木层中南山花和鸭脚木、细枝龙血树、九节以及肉实树等4个种对间表现为显著的正联结,这些种一般多生长在林下湿润环境中,具有一定耐荫性,它们对生境的要求具有相似的要求,生态位重叠较高。 本文参照了一些研究的方法[28-33],先用方差比率法对种群整体的联结性进行检验,再以χ2检验为基础引入种间联结系数AC和共同出现率PC,种间联结性分析表明,这3个指数在反映种对间的关联性及关联程度上较为一致。 从理论上来说,群落演替初期,物种之间关联程度较低,竞争比较激烈,形成负关联,随着群落演替的进行,种间正关联强度会逐渐增加,物种之间的竞争排斥得以减弱,最终由于生态位的分化和群落中的植物种之间的协调作用而达到演替顶级,显示出明显的正关联特性[20-21,33-35]。因此,在滨海植被恢复与人工林营造过程中,应采取一些合理的抚育措施,选取正联结较强的树种,来缓解种间和种内竞争,例如假苹婆和罗伞树、肉实树和罗伞树,在林区进行采伐作业时,则可以将与优势木本植物联结性较小或者负关联的伴生树种进行剔除,推动群落朝着有利于物种稳定与共存的方向发展。2 结果与分析
2.1 群落组成及重要值

3.2 物种多样性分析

3.3 群落总体联结性
3.4 主要种群的联结性




3 结论与讨论
猜你喜欢
植物研究(2023年5期)2023-09-09 08:01:22河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42建筑与预算(2022年6期)2022-07-08 13:59:18绿色科技(2019年3期)2019-03-21 05:11:12文苑(2018年22期)2018-11-19 02:54:44珠海(2016年11期)2016-12-10 02:09:18中国蜂业(2016年1期)2016-01-30 12:06:13杂草学报(2015年2期)2016-01-04 14:57:55建筑与预算(2014年5期)2014-04-11 07:29:28中国海洋大学学报(自然科学版)(2014年12期)2014-02-28 12:22:02
