
不同轮牧方式对荒漠草原植物群落特征的影响
2019-04-23 07:14王晓芳马红彬1沈艳1许冬梅1谢应忠1李建平1李小伟1
草业学报 2019年4期
王晓芳,马红彬1,*,沈艳1,,许冬梅1,,谢应忠1,,李建平1,,李小伟1,
(1.宁夏大学西北土地退化与生态恢复省部共建国家重点实验室培育基地,宁夏 银川 750021;2.宁夏大学农学院,宁夏 银川 750021)
放牧是草地的主要利用方式[1]。放牧方式总体上可以分为自由放牧和划区轮牧[2],适宜的放牧方式有利于植物种类和多样性的维持与发展[3]。围封禁牧作为一项重要的草地恢复重建措施[4],使植被在没有家畜放牧干扰时得到一定程度的恢复[5],促进多年生草本植物的生长[6],植物地上生物量[7-8]、盖度、密度都会有不同程度提高[9-10]。短期封育也提高了对地上植被发展具有指示作用的芽库密度[11],而放牧抑制了牧草的分蘖与生长[4]。但研究也发现,长期的封育并不利于草地的更新发展[12],长时间封育抑制了植物的再生和幼苗的生长,甚至会导致植被的灌丛化[13],适当的放牧有利于植被更新[14]。合理的放牧改变了群落主要物种的生物量[15],会刺激植物的生长,使得牧草发生补偿或者超补偿生长,草地生产力增加[16]。适度放牧下,植物由地上竞争转为地下竞争[17],对植物功能性状优化有积极促进作用,有利于草原多样性的维持,对草原的退化有遏制作用[18]。
划区轮牧应是北方草地可持续集约放牧的主推方式[19]。划区轮牧在充分利用牧草资源的同时[20],能够显著提高群落物种多样性[21-22]和优势种比例[23],增加短花针茅(Stipabreviflora)草原种群空间分布的复杂性[24]。与传统自由放牧相比,划区轮牧提高了草地生态系统的生产力、稳定性和恢复力等系统功能,优势植物的现存量、生产力都比连续放牧有所提高[25]。与其他草原类型相比,荒漠草原稳定性较差,有关其利用方式也随着草畜矛盾的不断加剧而受到人们更多关注[26]。研究发现,在相同的载畜率条件下,划区轮牧在放牧时间利用与空间配置上更合理,使牧草在频度、盖度、重要值、生物量等方面都比连续放牧有所提高[27-28]。荒漠草原划区轮牧具有一定优势,在草地状况较差时,划区轮牧比连续放牧更有利于植被的恢复,适宜的轮牧方式可以维持草地生态系统持续发展。长期封育的草地并非处于最健康状态[29],在适宜载畜率下,划区轮牧与围封禁牧一样可以减少草地生态系统退化[30]。可见,合理的放牧方式对草原生态系统的维护具有重要作用[8]。
宁夏荒漠草原地处我国农牧交错带,是当地重要畜牧业基地和生态环境保护的主要区域,其生境脆弱,在人为的干扰下易出现逆行演替。2003年起宁夏实施了全面的草原禁牧封育政策,禁牧以来草原生态系统恢复显著,如何进行科学放牧,在维护草原生态系统的基础上兼顾畜牧业生产发展需求是需要进一步探讨的问题。为此,本研究以宁夏荒漠草原为对象,在前期放牧强度研究基础上[31],对禁牧和不同轮牧方式下的草原植物群落组成、群落特征、多样性和稳定性进行研究,旨在为该区草原合理放牧利用提供理论依据。
1 材料与方法
1.1 研究区概况
研究区设在宁夏盐池县花马池镇四墩子行政村曹泥洼自然村,地理位置为北纬37°47′,东经107°25′,海拔1380 m左右,属于典型大陆性气候,1月平均气温-8.9 ℃,7月平均气温 22.5 ℃,年均气温7.7 ℃,≥0 ℃的年积温3430.3 ℃,≥10 ℃的年积温2949.9 ℃,年平均无霜期162 d左右。多年平均降水量289.4 mm,年蒸发量2131.8 mm。试验区地貌为缓坡丘陵,地带性土壤为灰钙土,质地为沙壤和粉沙壤,pH值为7.5~8.5。地带性植被为荒漠草原,主要分布有短花针茅、糙隐子草(Cleistogenessquarrosa)、赖草(Leymussecalinus)、牛枝子(Lespedezapotanimii)、白草(Pennisetumflaccidum)等多年生植物和狗尾草(Setariaviridis)、猪毛菜(Salsolacollina)等一年生植物。
1.2 试验设计
2013年试验开始前,在研究区选择地形、土壤及植被一致的地段,设置围封禁牧(对照,CK)、连续放牧(continuous grazing,CG)、二区轮牧(two-paddock rotational grazing,TG)、四区轮牧(four-paddock rotational grazing,FG)和六区轮牧(six-paddock rotational grazing,SG)5个处理,共14个小区。其中围封禁牧和连续放牧各有1个小区、二区轮牧有2个小区(TG1、TG2区)、四区轮牧有4个小区(FG1、FG2、FG3和FG4区)、六区轮牧有6个小区(SG1、SG2、SG3、SG4、SG5和SG6区),小区采用随机排列方式(图1)。每个小区面积9.3 hm2(623 m×150 m),14个小区总面积为130.2 hm2。对各个小区架设围栏,留围栏出入口。

图1 试验样地分布Fig.1 Schematic diagram of grazing and experimental design
根据前期研究结果[31],各处理载畜率均为 0.75只·hm-2。放牧家畜选择健康、体重相近的2龄健康滩羊母羊91只,随机分为5组,其中,连续放牧7只羊、二区轮牧14只羊、四区轮牧28只羊、六区轮牧42只羊,围封禁牧小区禁止放牧。羊只体重组间差异不显著(P>0.05)。放牧时间为2013到2015年,每年5月17日开始放牧,10月7日结束放牧,放牧周期为48 d,轮牧频率为3,每年放牧144 d。
1.3 测定项目与方法
2015年试验期开始前,分种测定各放牧区和对照区植物地上生物量。试验期间,在各放牧小区内设置5个活动围笼,围笼大小为1.5 m×1.5 m×1.5 m。在每个小区放牧前和放牧后测定笼内外植物种类(组成),各植物种频度、盖度、密度和地上生物量,测定后立即将围笼移动一次。在对照区生物量达到最高峰时(8月上旬)测定相应植被指标。测定时样方面积均为1 m×1 m,其中物种组成、盖度、密度和地上生物量5次重复,频度30次重复。地上生物量于65 ℃烘至恒重。
试验期间,处理样地内沿对角线设置5个面积1 m×1 m固定样方,每个放牧周期结束后收集各固定样方中凋落物,带回实验室于65 ℃烘至恒重。在放牧结束时,各处理样地采用土柱法测定0~40 cm地下生物量,土柱直径为17 cm,3次重复。用干筛法进行根土分离,将筛后根系装入自封袋带回室内清洗后于65 ℃烘至恒重。
1.4 数据分析
采用 Excel 软件进行数据初步整理、计算和作图,采用DPS软件进行单因素方差分析(One-way ANOVA),检验植物群落盖度、生物量、凋落物、植物生产力以及植被多样性等指标的差异,选择最小显著性检验方法(LSD)进行多重比较。根据植被调查数据,计算以下指标。
植物地上净初级生产力用净初级生产量计算得出。对照区植物地上净初级生长量是指地上生物量达到最高峰时的产量。放牧区植物净初级生长量是指放牧开始前地上生物量与放牧期间地上生长量之和。放牧期间植物地上生长量为各轮牧周期生长量之和,各轮牧周期植物生长量由下式计算[32]。
植物生长量=(f-c)+(c-f)[(logd-logf)/(logc-logf)]
式中:c为笼外植物地上生物量,即时间为0(小区轮牧前)的生物量;d为时间1(小区轮牧后)的笼内植物地上生物量;f为时间1的笼外植物地上生物量。
物种重要值、Margalef丰富度指数(R)、Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(E)、Simpson优势度指数(C),计算公式如下[33-34]:

式中:S为每个样方中的物种数;N为样方中所有物种的总个体数;Pi为样方中第i种个体数占全部物种个体数N的比例。
用改进后的Godron方法来测定群落的稳定性[35]。首先把样地中不同物种的盖度由大到小排列,换算成相对盖度并从大到小依次累加;然后将样地内物种种类之和取倒数,按照植物种类的顺序也逐步累加,由对应的结果看百分之多少的种类占多大的积累频度;最后将种类百分数与相对频度的积累做一条散点图,并用一条平滑的曲线将其连接,再与直线y=100-x[坐标轴(0, 100)与(100, 0)坐标相连的直线]相交,交点即为所求群落的稳定性。一般的稳定性比值越接近20/80,那么群落就越稳定。
2 结果与分析
2.1 不同轮牧方式下植物群落组成
2.1.1不同轮牧方式下植物群落组成和重要值 不同处理下,植物优势物种及其重要值有所不同(表1)。CK中赖草为优势种,短花针茅、糙隐子草重要值亦较高,3种植物的重要值为39.43%。在放牧区,赖草和糙隐子草的重要值均有不同程度的下降,短花针茅成为优势物种,其重要值随着轮牧分区增加呈下降趋势。糙隐子草、牛枝子、沙芦草等优良牧草的重要值以CK草地最高,CG最低,FG和SG居中,这与这些植物的适口性、家畜的选择性采食有关。老瓜头等杂草在几种轮牧方式下的重要值总体以SG最高,FG次之,CK最低。

表1 不同轮牧方式下群落的物种组成及其重要值Table 1 Species composition and species importance value of plant community at different rotational grazing patterns (%)
-:未出现 No appearance.
2.1.2不同轮牧方式下植物生活型 不同处理下,植物的生活型功能群组成基本一致,调查到的41种植物可分为3种生活型(表2),但植物生活型比例和重要值比例不同。CK、CG、TG、SG、FG总数分别为22、30、32、34、34种,放牧使物种的种类增加。半灌木主要是牛枝子、老瓜头、猫头刺,其物种和重要值比例在各处理下差异性不显著,说明轮牧对半灌木物种数及重要值影响较小;CG草地中多年生草本的重要值比例为17.28%,显著低于其他方式,但一年生草本的重要值比例显著高于其他方式(P<0.05),其他处理间多年生或一年生草本重要值比例无显著差异,表明连续放牧使一年生草本植物增加,多年生草本的优势地位低于其他轮牧方式。

表2 不同轮牧方式下植物生活型Table 2 Life form of plants at different rotational grazing patterns
注:同列不同字母表示处理间显著差异(P<0.05),下同。
Note: Different letters in the same column mean significant differences(P<0.05), the same below.
2.2 不同轮牧方式下植物群落特征
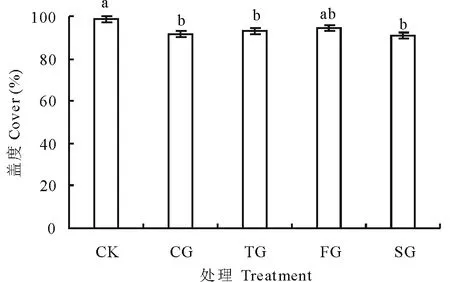
图2 不同轮牧方式下的群落盖度Fig.2 Coverage of plant community at different rotational grazing patterns不同小写字母表示不同处理间显著差异(P<0.05),下同。Different letters mean significant differences (P<0.05), the same below.
2.2.1不同轮牧方式下植物群落盖度及生物量 CK和FG处理的盖度较高,其他处理之间无显著差异(图2)。地上生物量以FG与SG较高,CG最低;地下生物量以CK与SG最高,CG和FG居中,TG最低(P<0.05)。
2.2.2不同轮牧方式下植物凋落物量和净初级生产力 凋落物总量以CK最高,FG和SG居中,CG和TG较低(P<0.05)。放牧试验前,各处理地上生物量差异不显著,但其生长量以FG和SG较高,CK和CG较低。FG与SG的净初级生产力要显著高于其他方式,CG、TG和CK差异不显著(P>0.05,表3)。
2.3 不同轮牧方式下植物群落多样性
轮牧方式对荒漠草原的群落多样性有显著的影响(图5)。其中丰富度指数(R)和多样性指数(H′)的变化规律一致。CG,TG、FG和SG的丰富度指数(R)和多样性指数(H′)都显著高于围封禁牧草地(P<0.05),各轮牧区二者之间差异性均不显著。均匀度指数(E)以CK最高,TG次之,FG最小(P<0.05)。优势度指数(C)以CK最低,且在4种放牧方式下均无显著差异(P>0.05)。
2.4 不同轮牧方式下的群落稳定性
5种轮牧方式下的交点坐标分布均远离(20,80)稳定点,说明群落均处于不稳定的演替阶段(表4)。相比之下,CK群落的稳定性最高,TG次之,FG与SG稳定性接近,CG最低。

图3 不同轮牧方式下群落的地上生物量和地下生物量Fig.3 Aboveground and belowground biomass of communities at different rotational grazing patterns

图4 不同轮牧方式下群落的凋落物量Fig.4 Litter fall biomass of communities at different rotational grazing patterns
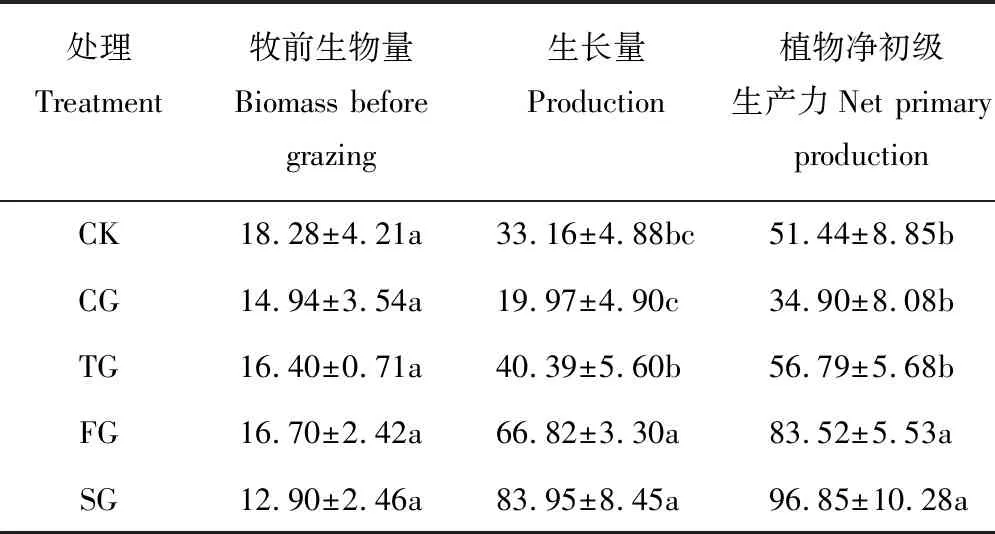
处理Treatment牧前生物量Biomass before grazing生长量Production植物净初级生产力Net primary productionCK18.28±4.21a33.16±4.88bc51.44±8.85bCG14.94±3.54a19.97±4.90c34.90±8.08bTG16.40±0.71a40.39±5.60b56.79±5.68bFG16.70±2.42a66.82±3.30a83.52±5.53aSG12.90±2.46a83.95±8.45a96.85±10.28a
3 讨论

图5 不同轮牧方式下的群落多样性Fig.5 Species diversity of plant community at different rotational grazing patterns
荒漠草原是宁夏草地生态恢复和合理管理的重要区域。其物种群落的组成、生物多样性、草地生产力均受到放牧方式的干扰[29,36]。本试验中禁牧封育植物种类最少,轮牧促进了物种组成增加,说明合理放牧下植物群落可进行一定程度物种组成的调整或生活型的改变,以更好地适应变化的环境[37]。相对围封禁牧,放牧草地由于家畜的选择性采食,使得一些耐牧、适牧的禾本科植物的重要值减少[38]。糙隐子草、牛枝子、沙芦草等优良牧草重要值下降且在连续放牧时最低,而禁牧封育草地没有人为因素的干扰和动物的践踏采食,将更多的资源投入到叶的干物质含量中,提高了光合作用能力,为这些牧草的光资源利用竞争提供了有利条件,也使得群落中毒害草重要值下降[39]。灌木由于家畜采食较少,在各处理间重要值比例差异不大。连续放牧情况下,放牧草地没有休养生息时间,家畜的连续采食抑制了多年生植物生长,为一年生植物提供了更多空间,表现为一年生植物重要值比例上升,多年生植物下降,不利于草地植物群落结构稳定[40]。
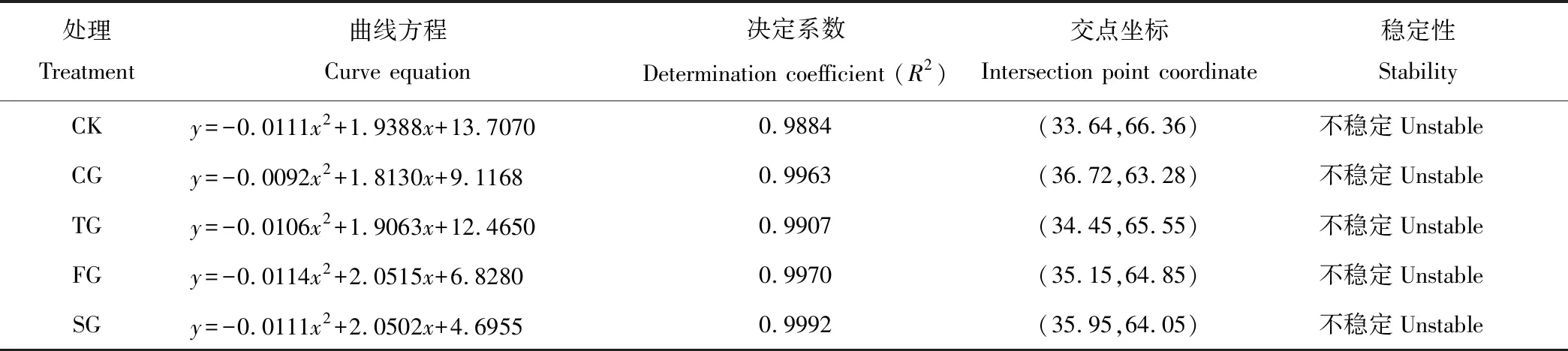
表4 不同轮牧方式下群落的稳定性 Table 4 Stabilities of plant communities under different grazing intensities
封育草地植物地上部没有被采食,光合作用没有受到影响[41],植被盖度、地下生物量维持在较高水平。适度轮牧情况下,家畜采食刺激了植物的再生[42],植物产生一个新的生长上限[43],轮牧中间的休牧时段,也使得植物的超补偿机制得到发挥[44],再者,根据小区植物恢复时间不同,四区和六区轮牧草地地上生物量增加,六区轮牧地下生物量较高,说明适宜的轮牧方式有助于生物量的提高[10,45]。植物的凋落物量受多种因素影响[46],植被组成、物种丰富度以及动物的采食都会导致凋落物量的变化[47]。本研究中,围封禁牧草地植被未被利用,凋落物增加。轮牧草地由于家畜采食后的再生量增加,提高了草地的生产力[25]。生态系统退化的最直接表现是群落地上生物量的减少和群落生产力的下降。所以与围封禁牧、连续放牧相比,合理的划区轮牧方式会促进植物的补偿性生长[48],提高草地的生产潜力。
围封禁牧草地的物种丰富度、多样性和优势度均显著低于轮牧区,均匀度则相反,这与禁牧区物种种类较少、轮牧小区杂类草增加有关(表1)。关于封育对群落多样性的影响,目前研究结论并不一致[9,49-51]。在本研究中,封育草地多样性要低于轮牧草地[52]。放牧家畜通过选择性采食、践踏等活动改变了优势物种,从而间接地影响群落的组成,改变种间竞争[53],削弱了群落优势物种排除其他物种的能力,从而提高了物种多样性,这也说明适度放牧干扰有利于群落多样性维持,符合“中度干扰”假说。
各处理下植物群落均处于不稳定的演替阶段,相对而言,禁牧草地由于没有家畜放牧干扰,群落稳定性增加,连续放牧草地由于家畜持续采食,植物恢复时间很少,群落稳定性下降[54],低于轮牧草地。研究发现,植物生物量、物种多样性与群落稳定性关系复杂[55],多样性与稳定性变化不一定呈正比[56-57]。二区轮牧在本研究中地上生物量、盖度等低于四区或六区轮牧,但稳定性高于这两种轮牧方式,这可能与群落中物种间的竞争强弱有关[57],还需进一步研究。
4 结论
轮牧对草地半灌木影响较小,但使植物物种种类增加、优良牧草重要值和凋落物量下降。增加轮牧分区有利于优良牧草和多年生草本重要值的保持,四区和六区轮牧有利于地上生物量和净初级生产力的增加。地下生物量以禁牧和六区轮牧较高。轮牧增加了草地植物物种丰富度、多样性和优势度,均匀度指数在禁牧草地最高。各处理下植物群落均处于不稳定的演替阶段,相比之下,禁牧群落稳定性最高。综合各轮牧方式下草地植被特征和生产中的便利性,研究认为四区轮牧是研究区最适宜的轮牧方式。
猜你喜欢
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
绿色科技(2021年10期)2021-06-23
林业科技(2020年3期)2021-01-21
今日农业(2020年19期)2020-12-14
南方农业·中旬(2020年9期)2020-11-30
黑龙江工程学院学报(2020年5期)2020-10-21
绿色科技(2017年15期)2017-09-01
中国科技信息(2015年2期)2015-11-16
江苏农业科学(2014年3期)2014-07-16
