
基于农艺性状的甘蓝型油菜三系骨干亲本多样性分析
2019-04-16 10:08:26张东锁胡胜武
西北农业学报 2019年3期
张东锁,臧 珊,胡胜武,郭 媛
(西北农林科技大学 农学院,陕西杨凌 712100)
油菜是世界上重要的经济作物之一,也是中国最重要的油料作物,含有丰富的油脂及蛋白质,是现代重要的食用油和工业油来源[1]。甘蓝型油菜(Brassicanapus, AACC, 2n=38)是由二倍体亲本白菜(Brassicarapa, AA, 2n=20)和甘蓝(Brassicaoleracea, CC, 2n=18)天然杂交后染色体组加倍得到的异源四倍体物种[2]。甘蓝型油菜起源于欧洲,于20世纪30至40年代分别由欧洲和日本引入中国,目前已经成为中国油菜的主要栽培类型[3]。甘蓝型油菜杂种优势显著,杂种优势的幅度可以达到30%~60%[4],杂交育种是油菜新品种选育的主要途径,育种材料的遗传变异和种质资源遗传多样性的研究对于选择亲本材料,合理选配杂交组合非常关键。目前,多种细胞标记、生物化学(同工酶)标记和DNA分子标记(SSR、SRAP、AFLP等)等技术被广泛地应用于作物遗传多样性的研究[5],然而,在育种的实施过程中,选育的品种往往要具备很多的符合生产实践的重要性状,表型性状的鉴定和描述仍然是检验遗传变异最基本的方法和途径[6]。全面分析种质资源的遗传多样性模式,不仅需要从基因层面探讨其多样性,同时也需要表型多样性的数据[7]。前人已经对小麦[8]、大麦[9]、番茄[10]、茶树[11]等作物的表型多样性进行了研究,并在揭示表型变异规律和分类方面取得了有价值的结果。对于油菜育种者而言,育种过程中的农艺性状和品质性状至关重要[1]。陈卫江等[12]利用株高、单株总荚数、千粒质量等10个农艺性状对24份甘蓝型油菜细胞质雄性不育保持系和恢复系亲本进行了遗传距离测定分析,并对杂交亲本的遗传距离与杂种优势的相关关系进行了探讨。结果表明,亲本遗传距离与杂种优势间存在一定程度的正相关。Hu等[13]利用9个农艺性状对国内外的63份甘蓝型油菜进行聚类分析,结果发现可以明显的将中国和欧洲的甘蓝型油菜划分为两大类,为拓展国内资源提供了理论依据。Körber等[14]利用518份甘蓝型油菜自交系研究了幼苗发育的多样性及其与农艺性状间的关系,在8种检测的种质类型中发现了表型多样性的差异。Chen等[15]对世界各地搜集到的488份甘蓝型油菜核心种质的表型性状进行了评估,分析12个产量与农艺性状、10个品质性状的遗传变异,该信息为育种亲本的选择提供了有用的支撑。此外,该研究挖掘到多份优异基因资源和特异遗传材料,可直接用于生产和遗传研究。
本试验对30份甘蓝型油菜三系骨干亲本材料的11个农艺和品质性状进行了鉴定,并对其遗传多样性进行分析,划分育种骨干亲本的杂种优势群,旨在为油菜杂交育种的亲本选配和生产利用等提供有效的信息和重要的参考依据。
1 材料与方法
1.1 材 料
供试的甘蓝型油菜骨干亲本材料共有30份,包括15份恢复系和15份保持系材料(基于Pol/Shaan2A CMS分类的恢保关系),均为低芥酸和低硫苷材料(表1)。这些材料包括‘秦优7号’‘秦油10号’‘陕油8号’‘陕油803’等审定油菜杂交种的父本,或中双系列、浙双系列等常规油菜品种的高代选系。以上材料均由西北农林科技大学油菜研究中心生物技术育种课题组提供。

表1 供试油菜品种编号、来源、类型和品质Table 1 Code, origin, type and quality of rapeseed accessions
注:“00”代表低芥酸,低硫苷。类型代表基于Pol/Shaan2A CMS分类的恢保关系。
Note:“00” represents low content of erucic acid and low content of glucosinolates. Type represents restorers and maintainers of Pol/Shaan2A CMS.
试验于2016至2017年在陕西杨凌西北农林科技大学农学院标本区(北纬34°16′,东经108°)试验地进行,采用随机区组试验设计,3次重复,每个株系种植5行,行长2.0 m,行间0.50 m,植株间距0.15 m,每行15~20株,四周设置保护行,土壤肥力中等,其他田间管理按常规进行。
1.2 农艺和品质性状调查
成熟期在每小区中部随机选择5株进行调查,调查记载的性状包括:株高(PH)、分枝部位(SPFPB)、一次有效分枝数(NB)、主花序有效长度(LTM)、主花序有效角果数(NSTR)、单株角果数(NSP)、每角粒数(NS)、角果长度(SL)、千粒质量(TSW)和单株产量(SYP)10个相关性状。收获种子后,参照文献[16]的方法,利用近红外仪(Foss NIR Systems Inc, Denmark)对小区混合种子进行含油量(OC)测定。
1.3 数据分析
采用SPSS11.0软件[17]的单因素方差分析(One Way ANOVA)程序对恢复系和保持系的11个农艺性状和品质性状进行比较。单株产量为理论值,由全株有效角果数、每角粒数、千粒质量的乘积得来。对每个小区下的株高、分枝部位、一次有效分枝数、主花序有效长度、主花序有效角果数,单株角果数、每角粒数、角果长度、千粒质量、单株产量和含油量等11个性状的原始数据用Excel整理后,采用随机区组试验设计进行11个农艺性状和品质性状的方差分析(Analysis of Agronomic and Quality Variance,AAQVA),11个性状的平均值用于聚类分析。根据马氏距离计算材料间的成对距离。利用DPS数据处理软件对各项平均值进行多元统计分析;聚类分析采用Ward’s法(离差平方和法)[18]进行系统聚类。所有材料的主成分都是从相关系数中提取出来的。利用DPS软件对11个性状进行有效矩阵分析。用SPSS11.0软件的散点图绘制前2个主成分的坐标图。
2 结果与分析
2.1 方差结果分析
30份参试甘蓝型油菜被分成2组,一组为恢复系材料,一组为保持系材料。对11个农艺性状和品质性状的方差分析(AAQVA)结果发现(表2):在恢复系材料中,11个农艺性状的变异系数从大到小依次为单株产量、分枝部位、单株角果数、角果长度、主花序有效角果数、每角粒数、一次有效分枝数、主花序有效长度、千粒质量、含油量、株高,说明供试材料在恢复系材料差异在单株产量、分枝部位、单株角果数等性状上变异丰富,在千粒质量、含油量、株高方面差异小。除株高和含油量变异系数分别为8.09%和8.33%外,其余性状变异系数都在10%以上,最大为单株产量,其变异系数为33.57%。在保持系材料中,11个农艺性状的变异系数大小与恢复系的基本一致,最大为单株产量(39.65%),最小为株高(6.87%)。然而,所有测试性状在恢复系与保持系上没有显著性差异(表2和表3)。
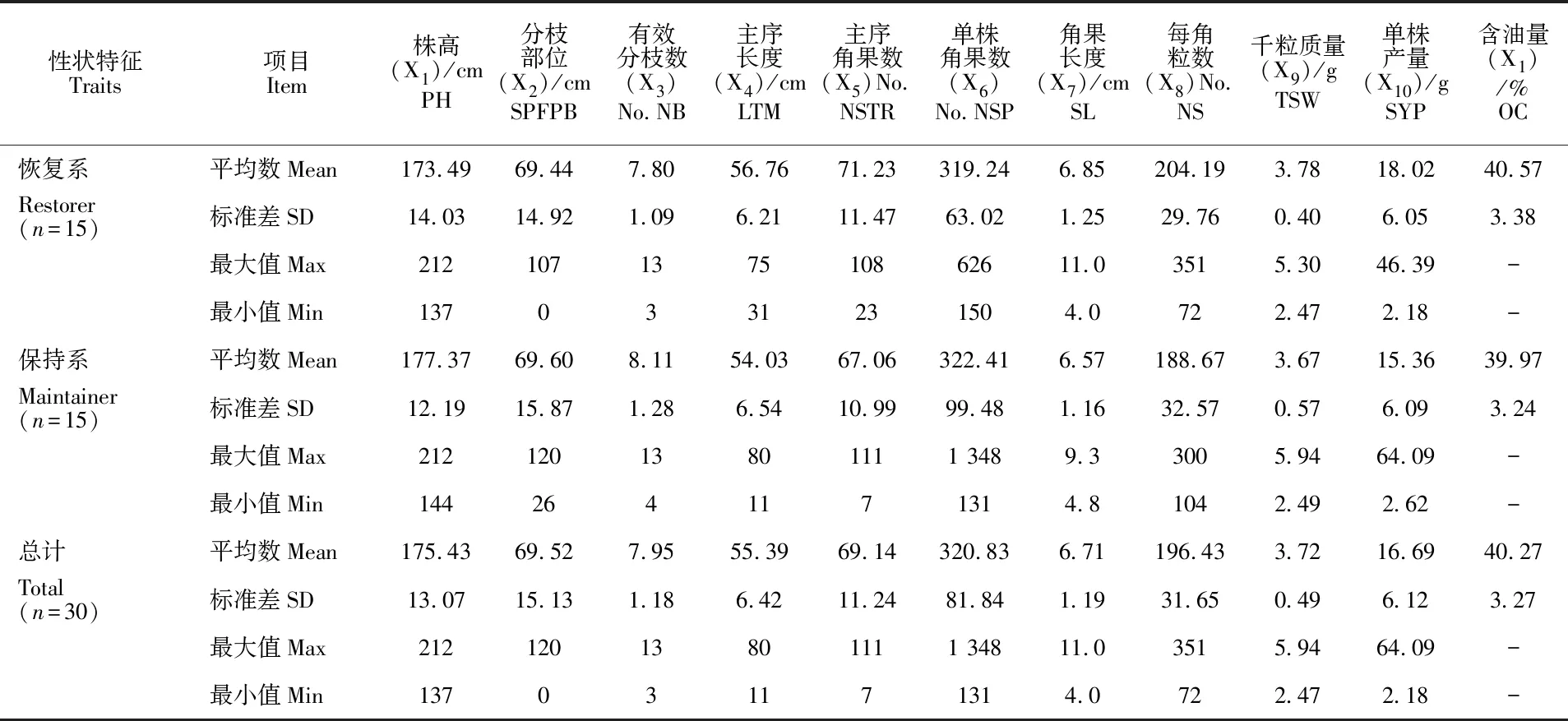
表2 恢复系与保持系11个农艺性状和品质性状的基本统计数据Table 2 The basic statistic data of 11 agronomic and quality traits between restorer and maintainer lines
注:PH.株高;SPFPB.分枝部位;NB.一次有效分枝数;LTM.主花序有效长度;NSTR.主花序有效角果数;SL.角果长度;NSP.单株角果数;NS.每角粒数;TSW.千粒质量;SYP.单株产量;OC.含油量。
Notes:PH.plant height;SPFPB.setting position of the first primary branch;NB.number of primary branches per plant;LTM.length of terminal raceme;NSTR:number of siliques of terminal;SL.siliques length;NSP.number of siliques per plant;NS.number of seed per silique;TSW.thousand seed mass;SYP.seed yield per plant;OC.oil content.

表3 恢复系与保持系11个农艺性状和品质性状的变异系数Table 3 The variation coefficient of 11 agronomic and quality traits between restorer and maintainer lines %
2.2 聚类分析
根据11个农艺性状的原始数据,利用Ward’s最小方差方法对30份油菜供试材料进行聚类分析,并构建聚类图(图1)。当遗传距离的临界值为39.73时,将30份参试材料分为三大类(Cluster Ⅰ~Ⅲ):Cluster Ⅰ包括16个材料,其中12个为恢复系,如Q10C、QY211R、Z821R、S11R、QSC等;4个为保持系,2010B7、New B1、Zhong 5和Zhong 9。Cluster Ⅱ包括10个材料,其中7个为保持系,如2012B1、H15、Zhong 7等;3个为恢复系,HYZ01R、2000-5R和D1526。Cluster Ⅲ包括4个材料,ZY72、Zhong 2、ZY18和CZ49,均为保持系。结果说明,参试恢复系和保持系材料间存在一定的遗传差异。
2.3 主成分分析
利用DPS数据处理软件对11个农艺性状进行有效矩阵分析,并绘制散点图(图2)。PCA结果表明,PC1占总变异的27.38%,PC2占总变异的22.11%。PC1和PC2的公式如下:
PC1=-0.2915X1+0.0697X2-0.0518X3+0.2405X4+0.2628X5+0.4878X6+0.3515X7+0.0599X8+0.2755X9+0.2510X10+0.5258X11
PC2=0.3015X1+0.4600X2+0.3322X3+0.3259X4-0.2079X5+0.1214X6+0.3622X7-0.3332X8-0.3294X9-0.2587X10+0.0730X11
主成分分析(PCA)结果与聚类分析结果相似。从PCA分析结果(图2)中可以看出,30份材料主要被划分为3大类。除Zhong 5(No.17)、2010B1(No.21)、H15(No.22)与2010B7(No.24)4个材料外,其余11个恢复系材料都被划为一起,与在聚类分析结果(Cluster Ⅰ)基本一致;大部分保持系材料也被划在一起;Q10C(No.1)、Zhong 9 Ⅲ(No.19)和CZ49(No.29)3个材料在聚类分析中归为第3类。

图1 基于农艺性状构建的30份油菜材料的聚类Fig.1 Dendrogram of 30 rapeseed materials generated by agronomic traits

图2 从30份油菜材料的11个农艺性状中提取的前2个主成分坐标图Fig.2 Bioplot of first two major principal components extracted from 11 agronomic traits of 30 rapeseed accessions
2.4 甘蓝型油菜保持系和恢复系亲本间农艺性状差异分析
外部形态特征的变异在一定程度上反映了遗传变异的大小。由表4可以看出,保持系和恢复系材料内变异系数为86.40%,保持系和恢复系材料间仅为13.60%。由表5可以看出,保持系内差异最大为15.35%,恢复系内差异次之为7.46%,保持系与恢复系间差异最小为1.80%。总体来说,保持系和恢复系材料内部差异大于它们之间的差异。

表4 恢复系与保持系油菜农艺性状变异分析Table 4 Analysis of agronomic traits variance of rapeseed accessions of restorer and maintainer

表5 不同甘蓝型油菜亲本品种间的差异分析Table 5 Pairwise differences of population average
注:对角线数据为区内成对材料差异的平均值;对角线下数据为修正后的区间差异平均值。
Note:Diagonal elements:average number of pairwise differences within population(PiX);below diagonal:corrected average pairwise difference(PiXY-(PiX+PiY)/2).
3 讨 论
油菜骨干亲本的遗传多样性分析是杂交育种和杂种优势利用的重要基础工作。甘蓝型油菜种植历史较短,中国现有的甘蓝型油菜的原始资源大多来源于欧洲和日本,导致遗传背景相对比较狭窄,品种间遗传多样性普遍不高,可利用资源匮乏[19]。
形态学标记是指通过观察或者简单的仪器测量获得的性状,为作物种质资源的分类和多样性分析提供了基本依据[6]。农艺性状具有表现直观、便于识别、易于掌握以及与生产直接相关等特点,因而在种植资源的分类、育种亲本材料的选择、组配、以及育成品种的识别上具有重要的应用价值。谢永俊等[20]对收集到的15个国家的225份甘蓝型油菜的农艺性状以及含油量等进行评估与分析,筛选出22份优异国外种质资源,并在此基础上选育出了11个优质油菜新品种。Tian等[21]利用农艺性状对96份油菜材料进行了地理位置的聚类,结果表明:96份甘蓝型油菜可以聚类到3个地理位置上。高必军等[22]对甘蓝型油菜的主要农艺性状与单株产量的关系进行了分析,方差分析结果表明:试验所测定的油菜单株产量等10个性状在38个参试品种间存在显著差异。本研究选择本课题组多年选育成功的30份油菜三系骨干亲本株系,包括15份保持系和15份恢复系甘蓝型油菜,利用聚类分析、主成分分析及方差分析对它们11个农艺性状和品质性状进行研究,结果显示:参试亲本材料间存在一定的遗传差异,大部分保持系和恢复系材料被分开,同时,保持系和恢复系材料内部差异大于它们之间的差异。
形态标记虽易观察、操作方便,但表型数量较少,易受环境和作物生育阶段的影响;而DNA分子标记不受环境影响和基因表达的时空影响,遗传标记数量多,遍布整个基因组,稳定性好,多态性明显。因此,将形态学性状为主的表型分析与分子标记或其它方式相结合,已经成为目前研究种植资源的有效手段[23]。何余堂等[23]通过形态学鉴定和RAPD分子标记相结合的方法,对不同地理来源的82份白菜型油菜(B.campestrisL.)资源进行了分析,揭示了白菜型油菜在中国的起源与进化。蒲晓斌等[24]对73份西南地区芥菜型油菜资源遗传多样性进行了分析,基于15个植物学性状测量数据的聚类分析将其分为4大类,共14个亚类;RAPD标记多态性分析结果的聚类分析显示为3大类,共15个亚类,结果表明西南地区芥菜型油菜资源具有丰富的遗传多样性,聚类结果主要遵循地域和生态环境规律。Ana等[25]通过利用表型数据及RAPD标记对30份冬油菜品种进行了遗传多样性的评估。对本研究供试材料,臧珊[26]利用SSR和SRAP分子标记对其遗传多样性进行了研究。对农艺性状和SSR/SRAP分子标记遗传多样性结果比较发现,二者存在一定的一致性,两种方法都可以将供试材料聚类为3类,大部分保持系和恢复系材料被分开,群体内部遗传变异大于群体间的遗传变异。同时,二者也存在一定的差异,表型聚类中,第Ⅰ类16个材料在分子标记中仍有13个被聚在同一类,第Ⅱ类10个材料在分子标记中仍有6个被聚在同一类。可见,在2种聚类方法中被聚在同一类的材料居多。其次,2种方法都发现保持系和恢复系材料内部差异大于它们之间的差异,目前,不同于玉米,油菜中杂种优势群划分和杂优模式研究开展的较少[27-29],这可能是导致油菜保持系和恢复系之间遗传差异较少的原因,因此,需要加强该方面的研究,定向扩大遗传差异。综上,表型性状和分子标记对油菜三系骨干亲本的遗传多样性的鉴定存在一定的差异,但表现更多的是一致性,以上研究结果建立了对供试骨干亲本的整体认识和评价,对育种设计、材料创制和选择、杂交组合选配具有重要的指导价值。
猜你喜欢
园艺与种苗(2021年10期)2021-11-28 23:31:51
农业工程学报(2021年23期)2021-03-11 01:11:30
西藏农业科技(2019年3期)2019-11-04 00:35:14
西藏农业科技(2019年1期)2019-07-25 00:37:02
安徽科技学院学报(2018年5期)2019-01-18 00:36:42
中国种业(2018年8期)2018-08-28 06:40:46
西藏农业科技(2018年4期)2018-04-25 06:39:28
种子(2017年12期)2018-01-17 10:55:45
西南农业学报(2016年6期)2016-04-16 05:12:46
种子科技(2015年8期)2015-05-24 16:14:56
