
温度诱导对鼠李糖乳杆菌cspc mRNA基因表达和生长速率的影响
2019-04-15 01:48,,,,,,,*
食品工业科技 2019年6期
, ,,,, ,,*
(1.内蒙古医科大学药学院,内蒙古呼和浩特 010110;2.内蒙古医科大学附属医院药剂部,内蒙古呼和浩特 010059)
鼠李糖乳杆菌(LactobacillusrhamnosusGG,LGG)属于乳杆菌属(Lactobacillus),最适生长温度37 ℃,兼性厌氧[1-2]。LGG由两位美国科学家Sherwood Gorbach和Barry Goldin在上世纪80年代发现,该菌分离自健康人体肠道[3],大量研究证明LGG能够定植于肠道,通过调节肠道菌群,起到抵御病原微生物的侵袭等作用,具有预防和治疗腹泻、预防呼吸道感染、预防龋齿、抗过敏的作用,提高机体免疫力等功效[4],LGG在酸奶、饮料、奶粉、奶酪、保健品等均有应用。
通常,LGG可通过液态发酵生产工艺获得,进一步通过冷冻干燥的方法获得菌粉,菌粉是微生物的最佳保存方式,然而冷冻干燥会不可避免的造成乳杆菌细胞膜的损伤,具体的损伤包括:机械损伤、溶质损伤、细胞膜渗透性损伤[5]、蛋白质变性失活、pH平衡的破坏和细胞膜脂肪酸的变化[6-7]。然而,微生物也会通过改变膜的流动性和改变蛋白质的翻译来适应低温环境。冷休克蛋白(cold shock proteins,Csps)是微生物为适应低温环境产生的应激蛋白,大肠杆菌在低温诱导过程中会产生大量GspA蛋白,可占到细胞总蛋白合成的13%,随后又发现了8中同源性蛋白:CspB、CspC、CspD、CspE、CspF、CspG、CspH、CspI蛋白[8]。枯草芽孢杆菌在低温环境下有13个Csps蛋白被诱导产生。在低温条件下Csps蛋白可以作为RNA的分子伴侣,与mRNA结合,稳定RNA,促进菌体在低温条件下转录并翻译[9],其中CspA、CspB、CspG、CspI蛋白在低温诱导下产生,CspC、CspE蛋白在正常培养条件下产生[10,11]。然而,与CspA蛋白不同的是CspC蛋白的表达量随温度的升高而降低,使用基因敲除的方法去除大肠杆菌的cspc基因后,发现菌体的生长明显加快[12]。说明CspC有抑制菌体生长的作用,因此在生长过程中抑制CspC蛋白的表达可能会有利于提高菌体的生长速率。然而对于LGGcspcmRNA基因的研究还未见报道。
本文通过对LGG进行温度诱导处理,通过荧光实时定量PCR(RT-PCR)的方法检测cspc的表达,并且对诱导后的菌体进行生长速率的测定,以考察温度诱导对cspcmRNA基因表达的影响,以及对菌体生长速率的影响,揭示cspcmRNA基因与生长速率的关系,为提高LGG的产量提供一定的理论依据。
1 材料与方法
1.1 材料与仪器
鼠李糖乳杆菌(LactobacillusrhamnosusGG,LGG)GD20150520 内蒙古医科大学药学院生物制药实验室;酪蛋白胨、酵母粉、牛肉粉 北京奥博星生物技术有限责任公司;葡萄糖、乙酸钠、柠檬酸三铵、吐温-80、MgSO4、K2HPO4、琼脂粉 国药集团化学试剂有限公司;cspc基因引物序列(F:5′-TGATAA GGGTTACGGCTTCA′,R:5′-GGCCACGATCAGATTG TTCTAC-3′),产物长度140 bp、16S rDNA引物序列(F:5′-TAGCGTACCATCTGATCCAGTA′,R:5′-GAAGTCGTCGCGTTGACA-3′),产物长度187 bp 美国Invitrogen;RNA提取试剂盒(RNAiso PlusD9108A)、cDNA反转录试剂盒(PrimeScript RT reagent KitDRR037A)、RT-PCR试剂盒 日本Takara。
ViiA7荧光定量PCR仪 美国ABI;Mini-sub cell GT电泳仪 美国Bio-Rad;Mutiskan FC酶标仪 美国Thermo。
1.2 实验方法
1.2.1 鼠李糖乳杆菌的发酵 培养菌种采取四级发酵培养方法,一、二、三级为种子扩大培养基,四级为最终发酵培养基,一级培养基接入0.3 g的冻干菌种,混匀,放置恒温厌氧培养箱37 ℃静置培养16 h。二级培养基由培养好的一级发酵液按4%的接种量接入,培养12 h。以此类推,传代到四级最终发酵培养基。
1.2.2 活菌数的测定 采用平板混菌计数法[13]。
1.2.3 生长曲线测定 四级发酵液按不同的时间点(0、1、2、3、4、6、8、10、12、16、20 h)取样,测定发酵液活菌数,制作生长曲线。
1.2.4cspcmRNA基因表达量法检测 mRNA的表达量根据GenBank上报道的鼠李糖乳杆菌的cspc基因序列,利用Prime5.0软件设计RT-PCR法检测cspc基因的引物。提取鼠李糖乳杆菌总RNA,按试剂盒说明书方法操作,用酶标仪检测RNA纯度,OD260/OD280在1.8~2.0为符合要求的RNA。反转录cDNA,按照试剂盒说明操作,RNA上样量为2.0 μL(约500 ng),反应条件:37 ℃,15 min;85 ℃,5 s结束反应,得到cDNA。
荧光定量PCR,按照试剂盒说明书方法操作,以cDNA为模板,加入cspc基因的引物,通过实时荧光实时定量PCR(Real time PCR,RT-PCR)方法扩增cspc基因。反应条件:预变性95 ℃ 30 s;变性95 ℃ 5 s,退火60 ℃ 34 s读板,共40个循环;从65 ℃到95 ℃制作融解曲线。以16s rDNA为内参基因,以确定cspcmRNA基因的相对表达量。产物做电泳检测分析,胶回收,产物送测序Invitrogen公司,NCBI blast比对分析。
RT-PCR所获得的数据,利用2-ΔCt(ΔCt=cspc基因Ct值-16S rRNA Ct值)公式进行cspcmRNA基因在LGG中相对表达量计算。
1.2.5 温度诱导cspcmRNA基因表达试验 将鼠李糖乳杆菌四级发酵液分别置于37、39、41、43、45 ℃的恒温水浴中分别处理10、15、20、25、30 min,即按照5组(温度)×5组(时间)设计,共25组试验,每组试验设3个重复,37 ℃为对照组。RT-PCR法检测cspcmRNA基因的表达量[14]。
1.2.6 温度诱导后菌体的生长速率测定 在三级发酵接种到四级培养基培养4 h时,即菌体进入对数生长期后,按1.2.5确定的最佳诱导温度和时间进行温度诱导,之后继续培养4 h再次进行温度诱导,之后再继续培养12 h结束发酵。RT-PCR法检测菌体的cspcmRNA基因表达量,活菌数法测定菌体生长曲线,并计算生长速率,生长速率=(lg活菌数t1-lg活菌数t0)/(t1-t0)(t1、t0表示发酵时间点),以未进行温度诱导作为对照[14]。
1.2.7 冻干验证试验 为了考察温度诱导对鼠李糖乳杆菌冻干存活率的影响,在终止发酵后对菌体进行冷冻干燥[14]。鼠李糖乳杆菌四级发酵诱导组、对照组,发酵液经10000 r/min离心20 min,弃去上清液,保留菌体,加入冻干保护剂(冻干保护剂:25%脱脂奶粉、10%乳糖、10%抗坏血酸)混匀,-40 ℃预冷30 min,真空干燥36 h得到冻干菌粉。菌粉悬溶于100 mL PBS中混匀,活菌数法测定混悬液的活菌数,计算冻干菌粉总活菌数(冻干菌粉总活菌数=混悬液活菌数浓度×100 mL)。根据发酵液总活菌数(发酵液总活菌数=发酵液活菌数浓度×发酵液体积),计算冻干存活率。冻干存活率(%)=冻干菌粉总活菌数/发酵液总活菌数×100。
1.3 数据处理
每组实验设3个重复,试验数据均采用平均值和标准误差表示,应用Tukey test检验进行分析,p<0.05表示差异显著,p<0.01表示差异极显著。
2 结果与分析
2.1 鼠李糖乳杆菌生长曲线
鼠李糖乳杆菌生长曲线如图1。
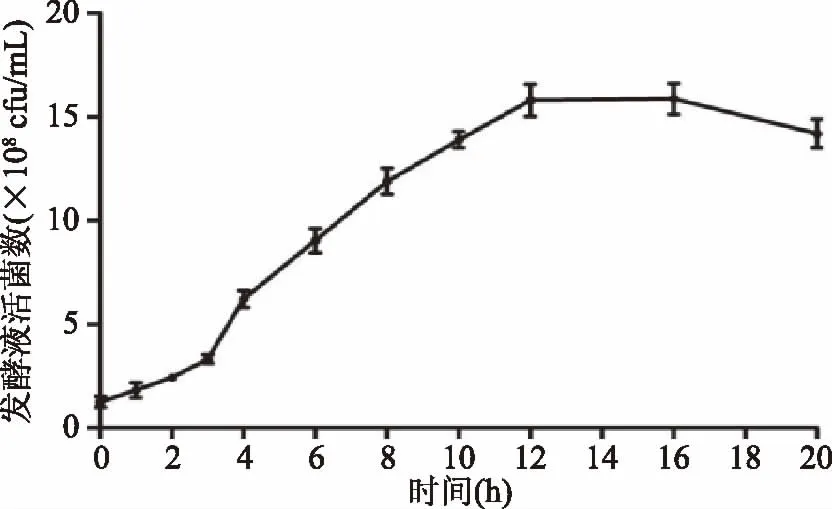
图1 鼠李糖乳杆菌生长曲线Fig.1 Growth curve of Lactobacillus rhamnosus GG
如图1所示,鼠李糖乳杆菌生长良好,0~3 h是菌体的适应期,3~12 h是菌体的对数生长期,12~16 h是菌体的稳定生长期,16 h之后菌体进入衰退期。
2.2 鼠李糖乳杆菌cspc基因克隆及电泳验证
鼠李糖乳杆菌cspc基因克隆电泳结果见图2。
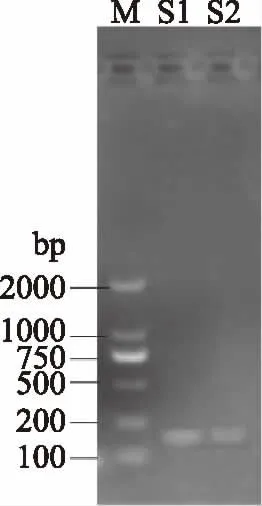
图2 cspc基因RT-PCR电泳结果Fig.2 Electrophoresis results of cspc RT-PCR product注:M:Marker;S1、S2:cspc基因样品1、2。
如图2所示鼠李糖乳杆菌cspc基因克隆片段,大小为140 bp,产物经电泳、胶回收、测序、比对,确认为cspc目标基因。
2.3 温度诱导对cspc mRNA基因表达的影响
CspC蛋白是CspA蛋白家族中的一个成员,与CspA蛋白不同的是,CspC蛋白不需要低温诱导表达,在37 ℃即可表达,研究显示[15-16],CspC蛋白与染色体凝集和细胞分裂有关,并且有抗转录终止的作用。Shenhar等[17]研究表明,CspC蛋白可以促进细菌胁迫应答因子(RpoS)的表达,RpoS是微生物在环境压力产生的适应性因子,因此可以说CspC蛋白是微生物对抗环境压力的重要因子。然而,其具体的生物学功能还不是十分明确。与CspA蛋白不同的是,CspC蛋白的表达量随温度的升高而降低,然而对于鼠李糖乳杆菌cspcmRNA基因的研究还未见报道。本研究通过升高温度抑制LGGcspcmRNA基因的表达,如图3所示,在对照37 ℃时,不同的作用时间对其表达量没有影响,而在温度高于37 ℃时,菌体cspcmRNA基因的表达量明显下调,说明温度升高可以抑制菌体产生cspcmRNA基因,且在43 ℃诱导30 min,菌体cspcmRNA基因的表达量最低,且差异极显著(p<0.01)。
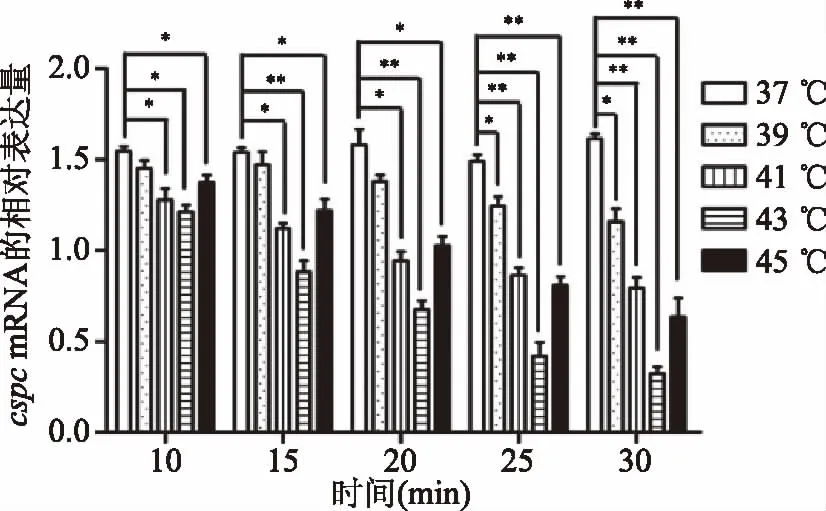
图3 温度诱导后csps mRNA基因的相对表达量Fig.3 Relative mRNA expression of csps after temperature treatment注:*代表差异性显著(p<0.05);**代表差异性极显著(p<0.01),图6同。
2.4 温度诱导后菌体的生长速率测定结果
本研究中,通过升高温度抑制菌体cspcmRNA基因的表达,结果如图4所示,在接种后生长第4和8 h,菌体经43 ℃、30 min诱导,菌体cspcmRNA基因的表达明显降低,而随着温度降至37 ℃,cspcmRNA基因的表达又恢复到原来水平。如图5所示,经过诱导后的鼠李糖乳杆菌生长明显加快,且活菌数明显升高。如图6所示,经过两次诱导后,与对照组相比,诱导组4~6 h以及8~10 h的生长速率明显增高,差异显著(p<0.05),而6~8 h和10~12 h两组的生长速率差异不显著。
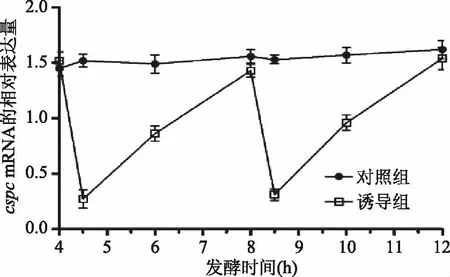
图4 温度诱导条件下菌体cspc mRNA基因的相对表达量Fig.4 Relative mRNA expression of cspc after temperature induction
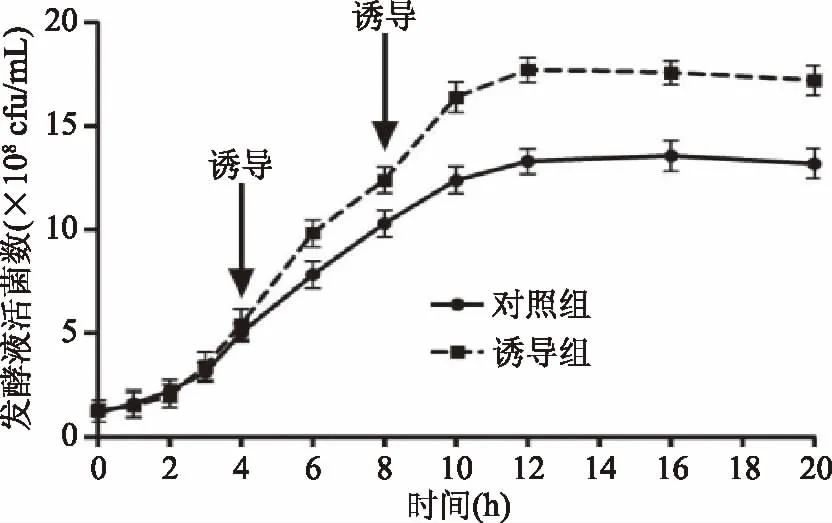
图5 温度诱导条件下菌体的生长曲线Fig.5 Growth curves of cells after temperature induction
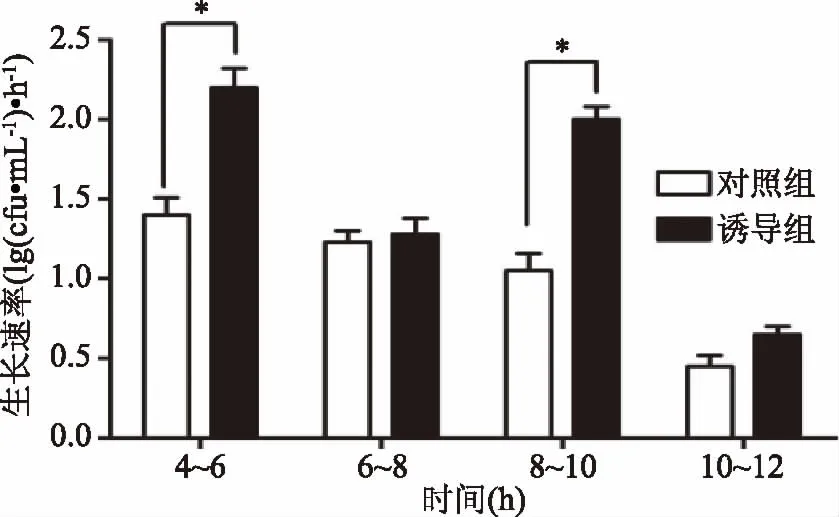
图6 温度诱导条件下菌体4~12 h的生长速率Fig.6 Growth rate in 4~12 h after temperature induction
2.5 鼠李糖乳杆菌冻干验证试验
如图7所示,诱导组与对照组相比,其冻干存活率无明显差异,说明温度诱导只影响菌体的生长速率,并不影响菌体的冻干存活率。
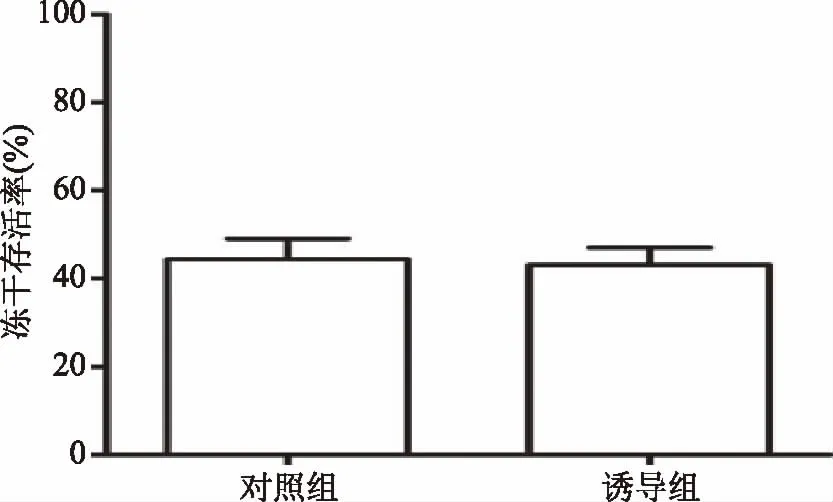
图7 菌体冻干存活率验证结果Fig.7 Testify result of freeze-drying survival rate of bacteria
3 结论
本研究对鼠李糖乳杆菌进行不同温度和时间的诱导处理,考察cspcmRNA基因的表达量,并考察在温度诱导后菌体的生长速率,结果显示,经43 ℃、30 min诱导,菌体cspcmRNA基因的表达明显降低,且诱导后菌体的生长速率明显升高,温度诱导相比基因敲除具有安全的优点,可应用于鼠李糖乳杆菌规模化发酵过程中,以提高菌体的产量。此外,研究显示,通过两次温度诱导,鼠李糖乳杆菌的冻干存活率并未下降,也证明了该方法的可行性。
猜你喜欢
化工管理(2022年9期)2023-01-04
当代水产(2022年1期)2022-04-26
华南理工大学学报(自然科学版)(2022年1期)2022-02-21
中国饲料(2021年17期)2021-11-02
食品与发酵工业(2021年7期)2021-04-27
中国洗涤用品工业(2019年4期)2019-05-11
中成药(2018年4期)2018-04-26
中国调味品(2017年2期)2017-03-20
中国海洋大学学报(自然科学版)(2016年12期)2016-12-24
中国兽医杂志(2016年7期)2016-08-30
