
Improved ε-Poly-lysine Production by Intergeneric Hybridization between Streptomyces griseofuscus and Brebvibacterium fl avum
2019-04-12 05:34LIShuDUZongjunCHENGuanjun
食品科学 2019年6期
LI Shu, DU Zongjun, CHEN Guanjun*
(State Key Laboratory of Microbial Technology, Marine College, Shandong University (Weihai), Weihai 264209, China)
Abstract: In order to improve the ε-poly-lysine (ε-PL) production of Streptomyces griseofuscus, intergeneric hybridization between S. griseofuscus and Brebvibacterium fl avum for enhanced level of the endogenous precursor lysine in the cells was investigated for the fi rst time in this study. One hybrid designated LS-32 was selected by using melanin as genetic marker,which produced 2.82 g/L of ε-PL in a shake fl ask, 1.73 folds as high as that of S. griseofuscus. In fed-batch fermentation, the ε-PL production of LS-32 was 56.4 g/L, which was increased by 38.9% as compared to that of S. griseofuscus. Randomly amplified polymorphic DNA (RAPD) analysis confirmed the intergeneric hybridization between S. griseofuscus and B. flavum. The activities of several key enzymes in LS-32 were greater than in S. griseofuscus. Furthermore, the concentrations of intracellular amino acids such as Asp, Glu and Arg in LS-32 were much higher than in S. griseofuscus.This research proves the importance of enhancing the endogenous lysine for improving ε-PL production and that intergeneric hybridization is a new breeding strategy for ε-PL production.
Keywords: ε-poly-lysine; endogenous lysine; intergeneric hybridization; fermentation
ε-Poly-L-lysine (ε-PL) is a homo-poly-amino acid in which L-lysine monomers are linked by peptide bonds between the carboxyl and the epsilon-amino groups[1-2].ε-PL is the most well-known for its antimicrobial activity towards microorganism. Besides, ε-PL has many other good qualities such as water solubility, biodegradability and nontoxic toward humans and environment, so ε-PL and its derivatives have been of interest for some valuable industrial applications, such as food preservatives, emulsifying agent,dietary agent, biodegradable fibers, drug carriers, etc.[3-4].Therefore, the commercial production of ε-PL is considered highly promising.
So far, a majority of studies have focused on strain breeding of ε-PL producers to improve ε-PL production.Hiraki et al.[5]employed an L-lysine analog named S-(2-aminoethyl)-L-cysteine (AEC) as the selective marker to breeding. Streptomyces albulus, and a mutant designated 11011A with higher ε-PL production of 2.11 g/L in shake fl ask fermentation was obtained. Li Shu et al.[6]improved the glucose tolerance of S. graminearus by genome shuffling while simultaneously enhancing its ε-PL productivity from 0.45 g/L to 0.86 g/L. Subsequently, the ε-PL yield of S. graminearus was further improved as high as 1.12 g/L by an interspecific hybridization through stochastic protoplast fusion[7]. Wang Liang et al.[8]adopted atmospheric and room temperature plasma (ARTP) mutagenesis combining with streptomycin resistance to screening Streptomyces sp.FEEL-1 and one strain designated AS3-14 was selected with ε-PL yield of 2.91 g/L in shake-flask fermentation.Furthermore, by adopting streptomycin and gentamicin as double antibiotic-resistant, ε-PL production of AS3-14 was strengthened as 3.83 g/L[9].
In the metabolic pathway of Streptomyces cells, L-lysine has been proved to be the precursor of ε-PL and exogenous lysine can be directly utilized for ε-PL synthesis by microorganism[10-11]. Based on this mechanism, Chen Xusheng et al.[12]expected a great increase of ε-PL production in S. albulus M-Z18 by adding exogenous lysine into fermentation broth, but the result was not so significant and ε-PL production was only 6.2% higher than that of the control. In contrast to that, no effects were observed on the ɛ-PL production by the supply of 11 mmol/L lysine in fermentation of Streptomyces sp. USE-51. Even worse of that, the ample supply of 110 mmol/L lysine caused a great reduction of ɛ-PL production for USE-51, from 3.2 g/L to 0.6 g/L[13]. Therefore, in order to obtain a significant increase of ε-PL production, we hypothesized that meaningful work should be focused on enhancing the synthesis of endogenous lysine in cells, rather than adding exogenous lysine.
Brebvibacterium fl avum (B. fl avum) is a commonly used industrial strain at present to produce L-lysine. So combining synthesis of endogenous lysine of bacteria with actinomycetes may be a new breeding strategy for improving ɛ-PL production. In this research, we investigated an intergeneric hybridization between S. griseofuscus and B. fl avum for the fi rst time to verify this hypothesis, and fi nally, the expected result was obtained.
1 Materials and Methods
1.1 Materials and chemicals
S. griseofuscus was screening out from soil and deposited in China Center for Type Culture Collection under the accession number of M209211[14]. B. fl avum was obtained from Xiwang Group Company (Zouping, China). Chemical reagents, which were used for enzyme activities detection,such as lysozyme, ATP, ADP, NADH, NADP, acetyl CoA,and 6-phosphate dehydrogenase, etc. were purchased from Sigma (USA) and Sinopharm Chemical Reagent Co. Ltd.(China); All other chemicals used in this work were of analytical grade[15].
1.2 Instruments and equipment
Stirred-tank bioreactor was purchased from Shanghai Baoxing Bio-engineering Equipment Company in China;Amino acid analyzer was purchased from German MembraPure GmbH; Microscopic camera was purchased from German Leica Company; PCR instrument was purchased from Bio-Rad Company in America; SK15 High-speed freezing centrifuge was purchased from Sigma Corporation of America; 1260 High performance liquid chromatography was purchased from Agilent Corporation of America.
1.3 Methods
1.3.1 Culture conditions
Bettyna (BTN) solid medium was as described previously[14]. B. flavum and spores of S. griseofuscus were spread on BTNagar plates for incubation under 30 ℃ for 3 d and 7 d, respectively. Regeneration medium (RM) and fermentation medium (M3G) were as described previously[7].Protoplasts of B. fl avum and S. griseofuscus and hybrids grew on RM agar plates and generated spores after incubation for 10 d under 30 ℃. Spores of hybrids and S. griseofuscus were transferred into shake flasks containing M3G medium in orbital shakers at 30 ℃, 200 r/min for 72 h. Each strain was grown in three separate shake flasks, and the mean values were reported.
1.3.2 Protoplast preparation and protoplast fusion
Protoplast preparation was as described previously[16]with some modification. B. flavum and spores of S. griseofuscus were respectively inoculated into M3G medium. After 24 h of incubation, mycelia of each strain were harvested by centrifugation at 4 000 × g, and then washed with PB buffer for two times. The mycelia suspension was exposed to lysozyme solution with concentration of 5 mg/mL for 1.5 h at 30 ℃. The protoplast formation was observed by microscope. After there was no residual mycelia in vision,protoplasts of both strains were centrifuged by 4 000 × g to remove lysozyme, and then suspended in PB buffer. For S. griseofuscus, the protoplasts were suffered from a high temperature of 70 ℃ for inactivation, and made sure that the inactivated rate was 100%. For protoplasts of B. fl avum, we maintained them alive. Then protoplasts of both strains were mixed together and fused by 40% aseptic PEG6000. After gentle shaking for 15 min at 40 ℃, the suspension of fusants was diluted with PB buffer and regenerated on RM agar plates for 10 days.
1.3.3 16S rDNA identification and randomly amplified polymorphic DNA (RAPD)
PCR primers for 16S rDNA amplification were as follows: forward primers: 5′-CGTGCTTAACACATG CAAGTCGAAC-3′; reverse primers: 5′-AGC AATGCTGATCTGCGATTACTAG-3′. Bacterial genome was extracted by a test kit (Sangon Biotech Co. Ltd.). Thirty arbitrary primers were purchased from Sigma (USA) and four of them were proved available: primer S5 (TCAGCGCATC);primer S9 (CGAGCCATCC); primer S17 (GTGTAACCTG);primer S26 (ACGGTTGTAT).
1.3.4 Fed-batch fermentation
Fed-batch fermentation was carried out in a 5-L stirredtank bioreactor. 3.2 L of aseptic fermentation medium was inoculated with 240 mL of seed culture. The initial pH was adjusted at 6.8 with NH3·H2O. The dissolved oxygen (DO)was kept at 10% by the automatic control of the stirrer speed.The change of pH during cultivation was detected by a pH electrode and NH3·H2O solution was added automatically to maintain pH at 3.8. The residual glucose was maintained at 10 g/L by automatically pulsed sterile glucose with a peristaltic pump into the culture medium. NH4+-N concentration was maintained at 0.2-0.5 g/L by the addition of an aseptic (NH4)2SO4solution with the concentration of 600 g/L.
1.3.5 Analytical methods
Dry cell weight (DCW) was determined by harvesting fermentation broth, fi ltering the mycelia and drying at 100 ℃for 10 h. The glucose concentration was determined by a biosensor analyzer SBA-40C (Shandong Academy of Sciences). ε-PL concentration was determined as the procedure described by Kahar et al.[17]. NH4+-N was analyzed by means of a colorimetric method using Nessler reagent.Intracellular amino acids were determined by harvesting culture sample, being treated in TCA solution, heating in boiling water for 30 min and detecting with amino acid analyzer (MembraPure GmbH). Enzymatic activities consisted of hexokinase (HK), phosphoenolpyruvate carboxylase (PEPC), aspartokinase (AK), citrate synthase(CS) and ε-PL synthetase (Pls) were measured according to Hamano[9], Murmu[18], and Wang Liang[19]et al. respectively.
1.4 Data and fi gure analysis
The photos were handled by software of Photoshop CS6.Datas and fi gures were compiled and constructed by Origin 8.0. Tables were created in Microsoft Office Word 2010.
2 Results and Analysis
2.1 Intergeneric hybridization between S. griseofuscus and B. fl avum
Protoplast fusion is an efficient breeding technique,which is characterized by the need not to rely on the homology of the microorganism, therefore the DNA recombinant can be extended to interspecies or even between genus cross, for example intergeneric hybridization between Monascus anka and Asperyillus oryzae[20], Lactobacillus delbrueckii and Bacillus amyloliquefaciens[21]. S. griseofuscus is a kind of prokaryotic microorganism, with the ε-PL production of 1.63 g/L in shake-flask fermentation, which was able to secrete melanin on solid agar (Fig. 1a) but not in submerged culture. While B. flavum is another kind of prokaryotic microorganism, with the L-lysine production of 55.2 g/L in shake-flask fermentation, could not secrete melanin on solid agar (Fig. 1b). Although S. griseofuscus and B. flavum belong to different genus, they are all prokaryotes. After protoplast preparation, these two strains were fused by PEG6000.
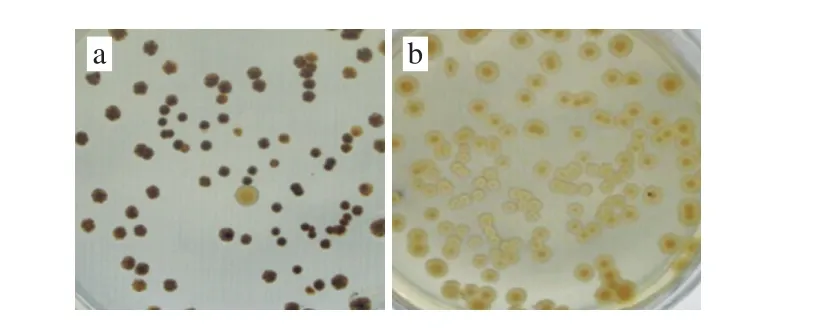
Fig. 1 Colony color of S. griseofuscus (a) and B. fl avum (b) growing on solid agar
Process of protoplast fusion could be observed by an optical microscope as shown in Fig. 2. Protoplasts first attracted each other and get together under the influence of PEG, and then cell fusion gradually completed due to the fl uidity of cell membrane. In microscopical vision, protoplasts fusion could be clearly seen including two-cell fusion, threecell and multiple-cell fusion. Among these hybridization,protoplasts fusion may occurred between S. griseofuscus species, or between B. flavum species, or between these two genus, respectively. A fact that should be noted is that we could not distinguish the S. griseofuscus and B. flavum protoplasts under microscope owing to the same appearance.Because prokaryotic microorganism has no nucleus,chromosome hybridization can be easily carried out after membrane fusion. Minority of fusion took place from the entire genome, forming diploids or polyploids[20]; Majority of fusion were recombined by partial genome[22]and few of them could not be fused[23], only forming heterokaryons.

1. two-cell fusion; 2. three-cell fusion; 3. four-cell fusion; 4. multiple-cell fusion.Fig. 2 Observation of protoplast fusion between S. griseofuscus and B. fl avum under microscopy
2.2 Hybrids screening and ɛ-PL production in shake-f l ask fermentation
Although protoplasts of S. griseofuscus were inactivated by heat, their chromosomes still remained active[20,22-23]and were able to fuse with live ones of B. flavum, generating hybrids which could secrete melanin again. Therefore,melanin of S. griseofuscus was an obvious genetic marker,which made us easily distinguish hybrids from parent species on RM plates (Fig. 3).
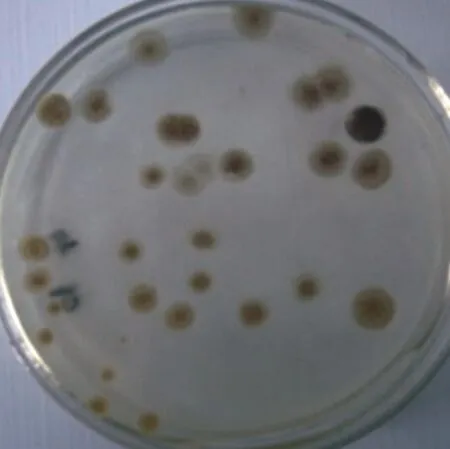
The arrowed colony was the hybrid with melanin as genetic marker.Fig. 3 Colonies growing on RM agar after protoplast fusion
After intergeneric hybridization, 54 hybrids were selected from RM plates with melanin as the genetic marker for the first-round screening. And for the secondround screening, ε-PL production of these 54 hybrids was tested through shake flask fermentation. As our expectation, three hybrids with significantly improved ε-PL production were obtained as 2.43, 2.72 g/L and 2.82 g/L,respectively, comparing with the 1.63 g/L of parent strain S. griseofuscus. The highest ε-PL producer designated LS-32 was subsequently proved to be genetically stable in eight continuous generations of subcultures, whose ε-PL production was (2.85 ± 0.12) g/L in shake fl ask fermentation.
2.3 16S rDNA identification and RAPD
16S rDNA sequences of hybrid LS-32, S. griseofuscus and B. fl avum were analyzed to compare their homology by amplifying the 16S rDNA with common primers for bacteria(Fig. 4). After genetic sequencing, complete 16S rDNA gene sequences of the above strains were detected as 1 484,1 465 bp and 1 452 bp, respectively (data not shown). Using the BLAST program in NCBI, sequence identities between LS-32 and S. griseofuscus was 99.6% while between LS-32 and B. fl avum was 84.3%. This result suggested that the hybrid LS-32 essentially belongs to S. griseofuscus species, but may be fused with partial genes from B. fl avum species, which made a contribution to the enhanced ε-PL production.

Fig. 4 Amplified fragments of 16S rDNA genes from three strains
In order to further confirm the parents of LS-32, RAPD was employed in this research. Thirty arbitrary primers were adopted to amplify genes under identical conditions, but most of them showed no specificity (Fig. 5E) except four. Adopting primer S9 (Fig. 5B), two fragments about 150 bp appeared in the same position for S. griseofuscus and LS-32, but not for B. flavum, suggesting that S. griseofuscus is a parent of LS-32, which was consistent with the result of 16S rDNA identification. Using primer S17 (Fig. 5C), a fragment about 200 bp appeared in B. fl avum along with in the LS-32, but not in S. griseofuscus, indicating that hybrid LS-32 had specific gene from B. fl avum. Besides, primer S5 (Fig. 5A) and S26(Fig. 5D) also showed the same results as primer S17, with fi ngerprints of 150 bp and 400 bp, respectively. RAPD results indicated that S. griseofuscus and B. flavum were indeed involved in intergeneric hybridization. Here, we must notice a fact that although 16S rDNA identification and RAPD proved that LS-32 was fused from S. griseofuscus and B. flavum,they could not elucidate which genes were responsible for the enhancement of ε-PL production, and this meaningful work needs further investigation.
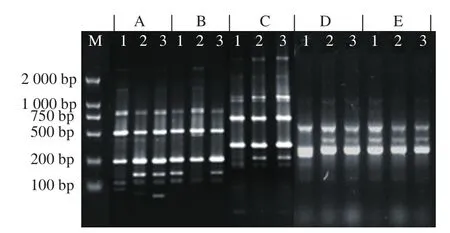
Fig. 5 RAPD fi ngerprints of primers for three strains
2.4 Fed-batch fermentation of hybrid LS-32
The industrial strains should not only have good performance in shake flask fermentation, but also exhibit high productivity in large-scale fermentation. Therefore, in order to test the real ε-PL productivity of LS-32, fed-batch fermentation was carried out in a 5-L fermentation tank with S. griseofuscus as the control. The changes of pH for both strains showed almost no difference, and dropped from 6.8 to 3.8 in the early 20 h (Fig. 6). The residual glucose was maintained at 10 g/L by automatically pulsed sterile glucose with a peristaltic pump into the culture medium. NH4+-N concentration was maintained at 0.2-0.5 g/L by the addition of an aseptic (NH4)2SO4solution. But for the consumption of glucose and NH4+-N, the two strains showed distinct difference, in that the feeding rates for LS-32 were more rapid than those of S. griseofuscus (Fig. 6), meaning that the metabolic intensity in LS-32 is stronger than that of the parent strain. As our expectation, the hybrid LS-32 produced much more ε-PL as 56.4 g/L at 192 h of fermentation process(Fig. 6b), about 38.9% higher over S. griseofuscus (40.6 g/L,Fig. 6a). Meanwhile, the final DCW of LS-32 reached 60.2 g/L, also higher than S. griseofuscus of 48.3 g/L(Fig. 6). The value of ε-PL/DCW represented the ε-PL synthetic ability and for hybrid LS-32 this value was 0.937,while for parent strain S. griseofuscus was 0.845. This comparison indicated that LS-32 obtained a stronger ε-PL synthetic ability by intergeneric hybridization between S. griseofuscus and B. fl avum, which proved the effectiveness of intergeneric hybridization as a new breeding strategy.

Fig. 6 Time-course curves of fed-batch fermentation of S. griseofuscus (a)and hybrid LS-32 (b)
2.5 Comparison for enzymatic activities between LS-32 and S. griseofuscus
In ε-PL synthetic route of Streptomyces, several enzymes were considered as the key enzymes, including HK, PEPC, AK, CS and Pls, etc[24-26]. So their activities were detected for comparison at 120 h of fed-batch fermentation.As a result, the activities of all above mentioned enzymes in LS-32 were higher than those in S. griseofuscus at different levels (Table 1). HK is the key enzyme in glycolytic pathway and its activity in LS-32 was 1.47-fold higher, which may be responsible for the rapider glucose consumption. CS is a key enzyme in TCA circle and catalyzes the first step of reaction. The activity of CS in LS-32 was 1.63-fold higher than that inS. griseofuscus, indicating that the metabolic activity of TCA circle in LS-32 was much stronger. This result had been also supported by anotherε-PL producing strain USE-51, who was reported that adding external citric acid into medium could obtain a significant increase forε-PL production[13]. In bacteria,L-lysine is mainly synthesized in diaminopimelic acid (DPA) pathway. DPA is formed via aspartic acid produced by combining oxaloacetate (OXA)with the NH4+-N as nitrogen source, where AK is the key enzyme for this reaction. Meanwhile, the carbon dioxide fixation reaction in bacteria is another source of OXA because PEPC catalyzed a reaction from PEP to OXA[27-28].Therefore, the higher enzymatic activities of AK and PEPC in LS-32 lead a greater fl ux to DPA pathway. Subsequently,polyreaction fromL-lysine toε-PL is catalyzed by Pls, whose activity was 77.1 U/mg in LS-32, 80.9% higher than that inS. griseofuscus, which may be the direct reason for enhancedε-PL productivity.

Table 1 Comparison of enzymatic activities of LS-32 and S. griseofuscus U/mg
2.6 Comparison for intracellular amino acids between LS-32 and S. griseofuscus
Activities of several key enzymes in LS-32 were proved to be more active than those inS. griseofuscus(Table 1),indicating a more active metabolism in the hybrid. Therefore,concentrations of intracellular amino acids inS. griseofuscusand LS-32 were compared. Some amino acids which exhibited a distinct difference were shown in Fig. 7, including Arg, Glu, Asp, Lys, Ser and Gly, whose concentrations in LS-32 were 53.4%-149.5% over that ofS. griseofuscus. Arg had the highest proportion among all the intracellular amino acids. In bacteria, Arg could be converted into NH3and ATP under the catalytic action by ADI system, while NH3was able to neutralize the H+, and ATP provided energy for H+-ATPase[29]. This result was consistent with the fact thatε-PL fermentation must depend on acidic environment(pH 3.5-4.0). The incensement of intracellular Asp and Glu contents was also beneficial to strengthen the synthesis ofε-PL, because Asp is the precursor of Lys in DPA pathway,while Glu provides -NH2for Lys synthesis[30]. As a result,Lys in hybrid LS-32 was 82.6% higher than that inS.griseofuscus(Table 1), which directly enhanced theε-PL productivity. Besides, concentrations of Ser and Gly in LS-32 were also obvious higher thanS. griseofuscus, implying that these two amino acids may be relevant to higherε-PL production, but the detailed mechanism needs further study.

Fig. 7 Comparison of the contents of intracellular amino acids in LS-32 and S. griseofuscus
3 Conclusion
In this research, an intergeneric hybridization betweenS. griseofuscusandB. flavumwas first investigated to improveε-PL production. Through protoplast fusion, one hybrid designated LS-32 was obtained withε-PL production of 56.4 g/L in fed-batch fermentation, which was 38.9%higher than that ofS. griseofuscus. RAPD analysis indicated that bothS. griseofuscusandB. fl avumwere indeed involved in intergeneric hybridization. Activities of several key enzymes and some intracellular amino acids in LS-32 were more active or much higher than those inS. griseofuscus, which was the possible reason for the enhancedε-PL production.This research emphasized the importance of enhancing endogenous lysine for improvingε-PL production and proved that intergeneric hybridization was a new breeding strategy for improving phenotype ofε-PL-producing strains.
