
三叠系的阶与牙形刺生物地层
2019-04-09 02:38:20王成源郎嘉彬
世界地质 2019年1期
王成源 ,郎嘉彬
1.中国科学院南京地质古生物研究所,南京210008;2.吉林大学古生物学与地层学研究中心,长春130026;3.吉林大学东北亚生物演化与环境教育部重点实验室,长春130026
1 国际三叠纪的年代地层划分
国际三叠纪划分为早、中、晚三叠世。早三叠世又划分为印度期和奥伦尼克期,中三叠世又划分为安尼期和拉丁期,而晚三叠世划分为卡尼期、诺利期和瑞替期,这样划分已基本定型(图1)。
值得特别注意的是,早、中、晚三个世或统,各个期或阶的时限,差别非常大。晚三叠世的时限最长, 仅卡尼期的时限就比早三叠世的时限长, 而诺利期的时限更是比早、 中三叠世的时限还长很多[1]。
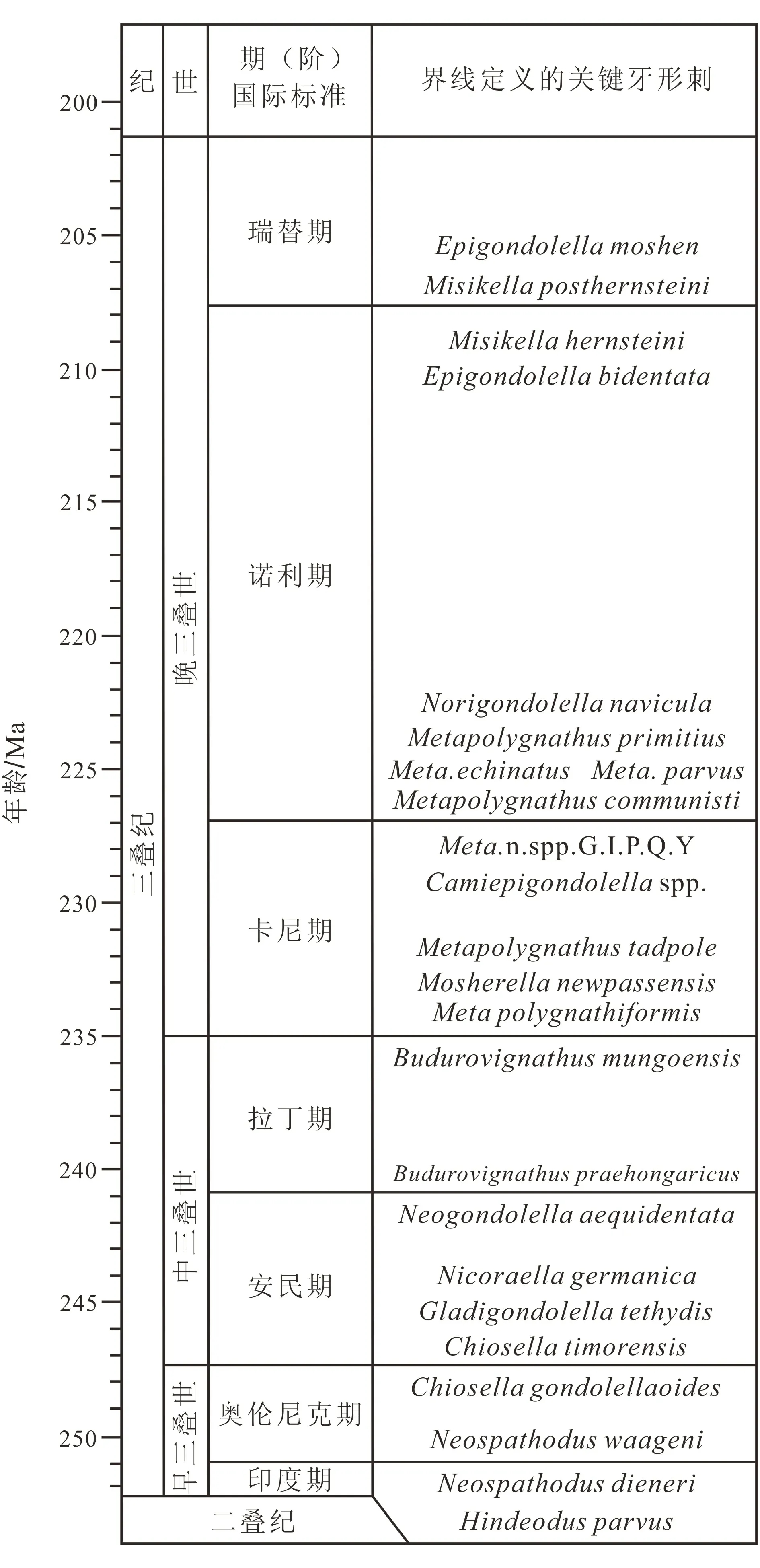
图1 三叠系的划分与牙形刺定义[1]Fig.1 Division of Triassic and definition of conodonts
中国三叠纪海相生物地层的研究,主要集中于早、中三叠世的研究,相对的,晚三叠世的海相生物地层研究较少。因为晚三叠世海相地层的分布仅局限于西藏、青海和黑龙江等边远地区。但对三叠纪生物地层研究而言,这个时期恰恰是时限最长,而研究精度最低,因此,存在较大的深入研究空间。
印度期的时限最短,但已划分出9个牙形刺带,而卡尼期、诺利期和瑞替期的时限都比印度期的时限长数倍;可以预测,中国晚三叠世牙形刺生物地层的进一步研究必将使牙形刺带数量进一步增加。
2 国际三叠系的阶与中国三叠系的阶
全国地层委员会于2001年正式提出有中国特色的年代地层划分方案[2]。2005年童金南,殷鸿福提出了中国三叠纪海相和陆相三叠纪的划分和建阶方案(表1)[3]。对于在海相地层另建中国自己的阶和在陆相地层建立中国自己的阶是有不同观点的。特别是在陆相地层的研究过程中,是否还要强调用中国自己的阶名,而不用国际地质年表的阶名,如印度阶与殷坑阶,奥伦尼克阶与巢湖阶,安尼阶与青岩阶,卡尼阶与亚智梁阶的取舍。国际地层委员会力推建立各个阶的GSSP(金钉子),就是要找到地层对比的共同语言,笔者认为,中国的阶名,凡是阶的底界定义与国际上同一阶的底界定义相同的,还是不用中国的阶名为好。与国际的阶名保持共同一致,没有另建中国自己的阶名的必要。中国三叠纪的区域性阶名,殷坑阶,巢湖阶,青岩阶,亚智梁阶和土隆阶都应当停止使用,统一使用国际地层委员会所认可的三叠系的阶名。中国建立的陆相地层的阶名也应当停止使用,国际地层委员会不主张陆相地层建阶。

表1 中国三叠纪年代地层划分系统[3]Table 1 Chronostratigraphic division of Triassic in China
“金钉子”的工作,就是建立各个阶的底界界线层型;由于阶是年代地层的最基本单位,因而也是建立“金钉子”的最直接和最核心的年代地层单位。这些阶名和底界定义已经成为全球地层工作者的共同语言,应当大力推广。中国在今后争取“金钉子”的过程中,也应尽量使用国际上目前确定的现有阶名,没有必要再推行自己的阶名,因为由“金钉子”确定的全球通用的阶,理应成为所在国的“区域阶”,定义上没有歧义。
例如,三叠系底界的全球层型已确定在中国的长兴,同时定义下三叠统和该统最下部的印度阶的底界,与牙形刺微小欣德刺(Hindeodusparvus)的首次出现一致。过去在中国所建的殷坑阶,定义与印度阶完全相当[4,6]。该“金钉子”建立后,如再继续使用殷坑阶一名,就欠妥当。在新编的《中国地层表》中[7],目前处理为“印度阶(殷坑阶)”,却使用了与奥伦尼克阶定义完全相同的巢湖阶,虽有进步,但仍然不妥。中国学者如使用巢湖阶去竞争奥伦尼克阶底界的全球层型,就不利于争取该“金钉子”在中国落户[8]。由此,笔者建议,《中国地层表》上的阶与《国际年代地层表》用“金钉子”确定的阶在底界定义完全一致、尤其是上覆阶底界定义也完全一致的情况下,统一使用国际上的阶名,不必再使用中国自己的阶名。殷坑阶和巢湖阶的阶名,都不应再使用,而使用国际上的统一标准:印度阶和奥伦尼克阶。
国际地层委员会当前的重要任务就是确定全球年代地层阶一级单位的“金钉子”,并完善《国际年代地层表》。自2002年起,该表每年至少更新一次,反映国际年代地层研究的最新进展。进入21世纪,互联网得到快速发展,国际地层委员会的官网也在2008年转移到中国科学院南京地质古生物研究所[9],这是对中国全球年代地层研究成果的肯定,中国就更应积极推广《国际年代地层表》的成果及其在中国的应用。
建立区域性的地层表的目的是要在条件成熟时,使区域性的阶逐步成为全球性的阶,但在中国有一种倾向,有的学者为突出建有大量中国阶名的“中国地质年表”的重要,在将国际与中国地质年代对比时,把《中国年代地层表》放在“国际与中国地质年代对比表”的左行[10];在《中国地层表》中,还建有大量的陆相地层阶一级的年代地层单位[11, 12]。众所周知,陆相地层变化大,其界线难以精确界定和进行大范围、长距离的地层对比,也找不到演化快的“主导门类化石”。这种局限性极大地降低了“陆相阶”的地层学意义。《国际年代地层表》中几乎所有的“金钉子”都是建立在海相地层中。国际地层委员会从来没有鼓励、也没有支持在陆相地层建阶。中国虽已在新生界建立了大量的陆相地层的阶,但没有一个阶得到国际地层委员会的认可,或在世界上得到公认。实际上,中国中、新生代除新近系和第四系外,其他各系(如三叠系、侏罗系、白垩系和古近系)等都有完整的海相地层发育(参见《中国地层表》)。笔者因此建议,凡是中国有相应的海相地层的,一律考虑在海相地层建阶,通过不断研究,逐步淘汰相应的“陆相阶”。同时要加强海陆交互相和陆相地层的生物地层的研究,侧重生物地层标志化石的确认,使陆相地层能较好地与海相地层对比。
作为年代地层中的基本单位,建阶是立典性的工作,因为阶是由两个地质时间界面所限定的地层,与形成这些地层的环境(相)完全不相干,地质时间也没有国界,所以全球阶应当通用于全世界。理论上,只要是这两个时间界面内形成的地层,无论它们是海相、海陆交互相、陆相、变质岩相还是火成岩相,都应是同名的阶,这是界线层型的属性所限定的[13],不应在不同相区的地层中分别建阶。
年代地层单位中显生宇的系,如寒武系、奥陶系、志留系和泥盆系等,都建立在欧洲,并早已被中国学者所接受。但是元古宇的一些全球系,如埃迪卡拉系、成冰系、拉伸系、窄带系、延展系、盖层系、固结系、造山系、层侵系和成铁系等仍未完全接受,建议逐步废除南华系、青白口系、蓟县系、长城系和滹沱系等中国自建的系名。目前,《中国地层表》中统一级的年代地层单位就已与《国际年代地层表》中的统一致。国际地层委员会致力于年代地层的精细划分,通过确定阶一级的“金钉子”,使全球年代地层的研究精度提高到阶一级。阶一级的年代地层单位,也要像系一级和统一级的年代地层单位一样,名称和定义世界统一,全球通用。笔者因此希望中国学者应努力做到采用《国际年代地层表》中所有已正式确定的阶名和阶的定义,真正使用国际上的统一标准。
《国际地层表》和《国际年代地层表》中阶的中译名至少有3种以上的不同的译法,阶的中译名也需要统一,建议采用最新的译法[14]。
3 三叠系阶的底界
3.1 瑞替阶底界
瑞替阶的底界,至今没有统一的意见,但放射虫,牙形刺,菊石,腕足类和瓣鳃类都是这一界线的重要门类。Carter依据放射虫的研究,倾向于将瑞替阶底界置于Betracciumdeweveri带和Proparvicingulamoniliformis带之间,从这个界线起,至少有3个放射虫的种出现,都是瑞替阶底界的标志[15,16]。
依据牙形刺的研究,瑞替阶的底界有不同的划分方案,较早的方案是划分在Misikellahernsteini或M.posthernsteini之底,近年来,主要的观点则是划分在EpigondolellamosheriMorphotype B的底界,此界线落在E.bidentata带之内。
菊石类的研究,主要将该界线置于Sagenitesguinguepunctatus带与Sagenitesreticulatus带之间,或Metasibirites带与Parachloceras/Cochloceras带之间[17]。
瑞替阶底界界线层的腕足类也比较重要。Milloš Siblik通过对界线层腕足类的综合研究,提出的界线位置在牙形刺bidentata带之内,M.hernsteini带之下[18]。这条界线在中国西藏南部有较好的剖面,非常值得进一步的研究。
现有资料证实,牙形刺并没有延伸到三叠纪顶界或侏罗纪的证据。以前报道的日本和非洲的科迈隆有侏罗纪牙形刺,都是地层时代鉴定错误的结果。
3.2 诺利阶底界
诺利阶底界的“金钉子”(GSSP)目前还没有确定,有两个候选剖面,一个是加拿大布列颠哥伦比亚的黑熊岭(Black Bear Ridge)剖面,代表北美区;另一个是意大利西西黑禹的Pizzo Mondello剖面,代表特提斯区。
北美区的诺利阶底界置于菊石Stikinoceraskerri带的底部[19,20],牙形刺Primatellaprimitia带之内,相当于Primatellaasymmatrica-Norigondolellasp.亚带之底,此亚带内有多个Primatella的种存在[21]。而Metapolygnathusex gr.M.echinatus的首次出现,非常接近菊石Stikinoceraskerri带的底界[22]。
特提斯区意大利Pizzo Mondello剖面的牙形刺与北美区的有很大不同:Carnepigondolellaex gr.samlieli的时限较高;Epigondolellaex gr.E.quadvat,P.primitia等出现较早;M.comunisti很常见。M.parvus和M.echinatus都是诺利阶底界的标志化石。
Orchard依据加拿大黑熊岭剖面的牙形刺,对卡尼阶与诺利阶的分界进行了详细的研究[21]。这个剖面厚度仅有5 m,但是卡尼阶顶部和诺利阶底部的地层中含有极为丰富的牙形刺和菊石。Orchard将此剖面的牙形刺,自上而下,划分出3个带和9个亚带,平均每个亚带地层仅有半米多厚,这样的研究精度和水平是世界上较高的[21]。
3.3 卡尼阶底界
卡尼阶底界的“金钉子”(GSSP)已确立在意大利的Prati di Stuores剖面[23,24],以菊石Daxatinacanadensis的首次出现为准,以牙形刺Metapolygnathusex gr.M.polygnathiformis的首次出现为铺助标准,两者在层型剖面上几乎是同时出现的,但牙形刺标本较少。
卡尼阶底界界线,在印度的Spiti剖面,美国内华达州的New Pass剖面,加拿大黑熊岭剖面,也有较好的代表。特别是黑熊岭剖面,Orchard对卡尼阶的底界有详细地研究[21,25,26]。
值得注意的是,这一界线的GSSP虽已确定。但在中国至今也没有找到菊石Daxatinacanadensis,并且国内对于牙形刺Metapolygnathusex gr.M.polygnathiformis标本的鉴定也不一致。中国的一些脊椎动物专家仍把含牙形刺Polygnathiformis的层位归为中三叠世拉丁期[27,28]。王成源等则将这一层归属到卡尼期[29]。
由于这一界线层型已经确定,界线划分的菊石、牙形刺的标准也已确定,只要化石鉴定准确,就应按GSSP的标准划分地层时代。
3.4 拉丁阶底界
拉丁阶底界的“金钉子”(GSSP)也已经确定,在意大利的北部Bagolino村附近的Caffaro River层[30],以菊石Eoprotrachycerascurioni的首次出现为准。牙形刺Budurorignathuspraehungaricus出现在下覆的菊石Nevaditessecedensis带的上部,此牙形刺种来源于Neogondolellaeguidentata。
Neogondolellaconstricta,Parogondolellaexcels,N.alpina出现于这一界线之下,有的可穿过界线。另一些对确定拉丁阶底界有用的种包括Neogondolellapostcornuta,“Ng.”trammeri和Gladigondolellamalayensis,主要见于特提斯区。拉丁阶底界界线的年龄为210~242 Ma[31,32]。
3.5 安尼阶底界
安尼阶底界界线层的最好剖面目前公认为罗马尼亚的Desli Caira剖面。这里奥伦尼克期最晚期的菊石Deslicarites层之上是安尼期最早期的Paracrochordiceras-Japonites菊石层,然后是Aegeiceras菊石层。现在,安尼阶底界的牙形刺标准是以Chiosellatimorensis的首次出现为准,但GSSP还没有最后选定。此种特征明显,分布广泛,在这一界线层,Neospathodusspathi,Gladigondolellacarinata,Triassospathodushomeri穿过界线不久,就立即消失[25,26]。中国有竞争力的剖面为贵州的关刀剖面,该剖面的牙形刺序列比较完整,但缺少菊石化石的记录[33],因此该界线的GSSP最终有可能选在罗马尼亚。
3.6 奥伦尼克阶底界
奥伦尼克阶包括2个亚阶:斯帕斯亚阶(Spathian)和斯密斯亚阶(Smithian)。
中国下三叠统奥伦尼克阶的牙形刺研究,以贵州甲戎剖面最为详细,可分为8个牙形刺带,史密斯亚阶4个带,斯帕斯亚阶4个带[34--36]。
斯帕斯亚阶的底界并没有最后确定,暂以Nv.pingdingshanensis的首次出现为准,斯密斯亚阶或奥伦尼克阶的底界以Nv.w.eowaageni的首次出现为准,但界线层型剖面点仍没有确定(图2)。
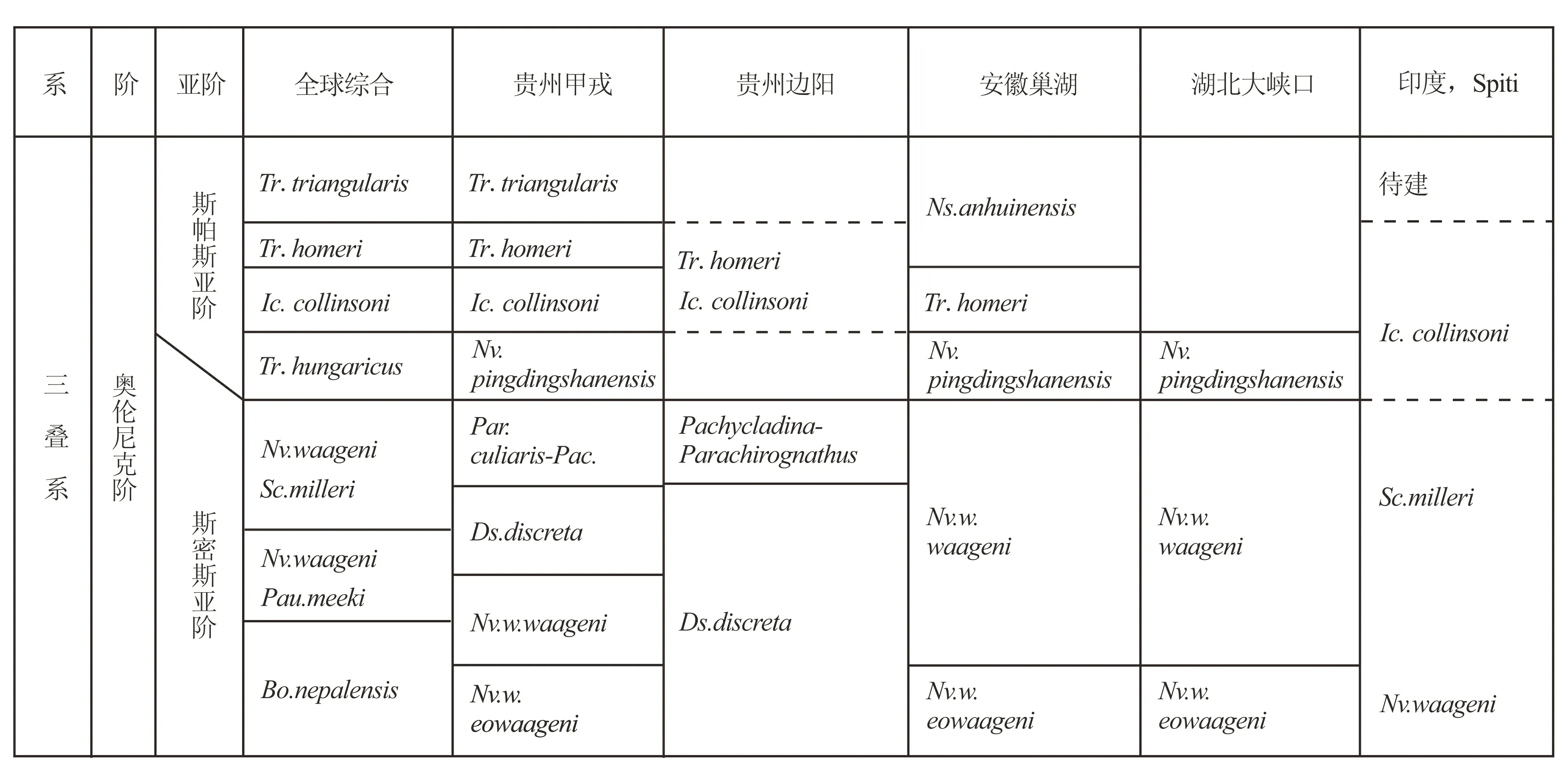
图2 奥伦尼克阶牙形刺带对比(据Chen et al.,2015[35], 略修改)Fig.2 Correlation of conodont zones in Olenekian stage
3.7 印度阶或三叠系的底界
二叠系—三叠系界线或印度阶底界的“金钉子”(GSSP)在中国长兴D剖面的27c之底确立,以Hindeodusparvus的首次出现为准[2]。Hindeodusparvus的分布是世界性的,低纬度、高纬度,浅水相、深水相均有此种的纪录。它的先躯种是Hindeoduspraeparvus。现在印度期早期的牙形刺已分为H.parvus,H.lolata,I.staesche,I.isacica4个带。
长兴期最晚期的带,自上而下一般划分为H.changxingensis带和Clarkinazhangi带,而Orchard将最高层位的牙形刺列为praeparvus带和latidentatus带[1]。
Kozur & Pjatakova在建立parvus种(当时称Archiognathodusparvus)时,就在文章题目中指出,parvus是三叠系底部的重要的标准化石[37]。殷鸿福等也提出将H.parvus作为三叠系之底的带化石[2]。长兴剖面上的H.parvus最早是由张克信[38]发现的,而长兴剖面GSSP的点位,最早是由王成源[4,39,40]确定的,已被普遍接受。
4 中国三叠纪牙形刺生物带
4.1 上三叠统牙形刺生物带
2018年Rigo et al.对特提斯区上三叠统的牙形刺生物带提出系统的分类, 将特提斯区上三叠统划分出22带,这是最新的划分方案[41]。与Orchard[21]的方案相比有以下不同:其中,诺利阶的底界置于Metapolygnathusparvus带之底,瑞替阶的底界确定为Misikellaposthersteni带之底,这与Orchard[21]的方案相一致,本文采用他们从图瓦里亚阶到瑞替阶的牙形刺带,但朱利安亚阶的牙形刺带采用中国自己的带(表2)。

表2 中国上三叠统牙形刺生物带Table 2 Conodont biozones of Upper Triassic in China
按现有的总结特提斯区上三叠统可划分为23带,整个三叠系可划分为50个牙形刺带。但有几个带在中国还没有发现,是今后要努力寻找的。
4.2 中三叠统牙形刺生物带
拉丁阶拉丁阶自上而下可分为2个亚阶:龙高巴丁亚阶(Longobarding),弗萨尼亚阶(Fssanian)(表3)。
在拉丁期牙形刺有以下4个事件值得注意:①接近菊石reitzi带的底部,Neogondolellacornutapostcornuta和ParagongdolellaalpineGroup开始出现;②菊石reitzi带的上部,Neogondolellapseudolonga,Paragondolellafueloepi,Paragondolellatrammeri开始出现;③在菊石secedensis带上部Neogondolellapraehungarica与Buduroviganthusgabriellae的先驱分子(这里暂时归入Neogondolellasp.A)一起出现;④Budurovignathustruempyi和B.hungaricus一起出现在Bagolino 柱状图66 m的层位。N.praehungarica和P.fueloepi的出现正是在拉丁阶底界定义的下方,完全可以作为拉丁阶底界的标志[30,31]。
中三叠统拉丁阶底界的“金钉子”是2005年确定在意大利北部阿尔卑斯山南侧的Bagakino村南部Caffaro河南岸(45°49′9.5″N,10°28′15.5″E),层位是在Buchenstein 组底部之上5 m的瘤状灰岩之中,菊石Eoproterachycerascurioni出现的最低层位,即Eoproteracherascurioni带的底界。
拉丁阶底界的GSSP第二个标志是出现在安尼阶最顶部的牙形刺Neogondolellapraehungarica,此界线距今大约241 Ma。
安尼阶安尼阶自上而下分为4个亚阶:伊利莲亚阶(Illyrian),佩尔松亚阶(Pelsonian),毕塞妮亚阶(Bithynian),艾格亚阶(Aegean),目前亚阶的划分和牙形刺带的划分只是大致的对比,存疑较多。伊利莲亚阶和佩尔松亚阶上部可与Neogongdolellaconstrictacornuta带-Neogondolellaconstricta带-Paragondolellabifurcata带对比;早、中佩尔松亚阶相当于Niccoraellakockli带;毕赛妮亚阶大致相当于Nicoraellagermanica带,艾格亚阶可与Chiosellatimorensis带大致对比。
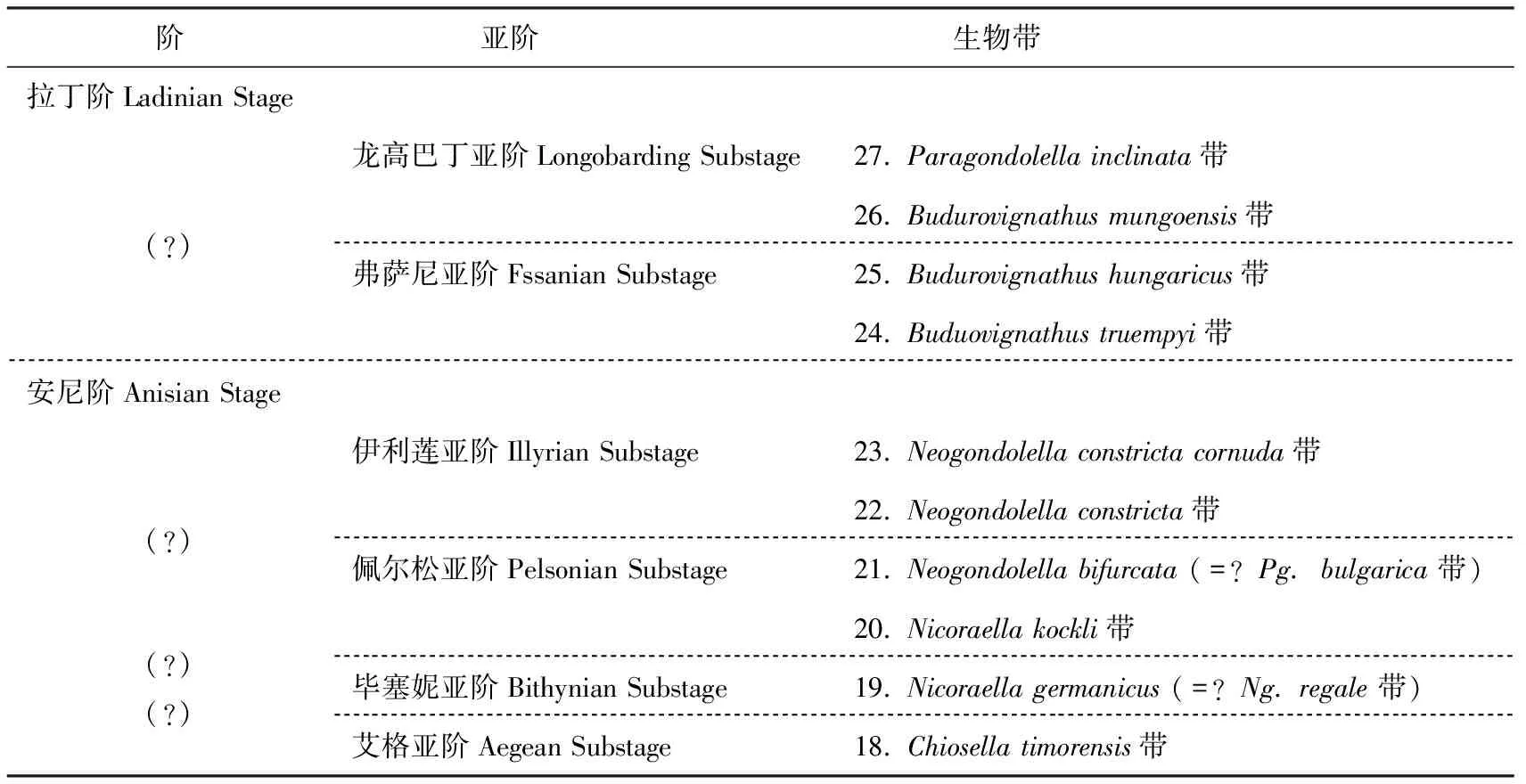
表3 中国中三叠统牙形刺生物带Table 3 Conodont biozones of Middle Triassic in China
4.3 下三叠统牙形刺生物带
奥伦尼克阶奥伦尼克阶包括2个亚阶:斯帕斯亚阶 (Spathian)和斯密斯亚阶(Smithian)(表4)。
中国下三叠统奥伦尼克阶的牙形刺序列,以贵州甲戎剖面最为完整,Chen et al.将其分为8个牙形刺带, 斯密斯亚阶4个带, 斯帕斯亚阶4个带[36]。
值得特别注意的是斯密斯亚期和斯帕斯亚期之间海水氧化还原条件的大幅度变化,在斯密斯亚期晚期,广泛分布的缺氧事件,一直持续到斯帕斯亚期早期的Novispathoduspingdingshanensis带,缺氧事件可能是导致斯密斯亚期—斯帕斯亚期过渡时期的牙形刺小型化的重要原因,但在斯帕斯亚期早期之后,进入Icriospathoduscollinsoni带时,氧化还原条件发生巨大变化,迅速进入富氧的红纹石灰岩相(Griotte facies),这是由于海水温度下降、含氧量增加形成的特殊岩相[42]。

表4 中国下三叠统牙形刺生物带Table 4 Conodont biozones of Lower Triassic in China
Krystyn et al. 及Orchard & Krystyn建议用Novispathoduswaagenis.l作为印度阶和奥伦尼克阶的分界[43,44],而Zhao et al.推荐用Novispathoduswaagenieowaageni作为印度阶与奥伦尼克阶的分界[45--48]。在贵州甲戎剖面和西澳大利亚用Novispathoduswaagenieowaageni更适合[36]。Chen et al.依据贵州甲戎剖面建立了Discretelladiscreta带[36]。依据此带贵州甲戎剖面可以与边阳剖面对比。在这两个剖面Discretelladiscreta带都是位于Pachycladina-Parachirognathus组合带之下。Discretelladiscreta最早报导于美国内华达州[49]。此种同样见于加拿大西部和加拿大北极区[50,51]、克什米尔[52]、印度Spiti[53]、贵州关刀和边阳剖面[33,54]、亚洲东北部俄罗斯滨海地区[55]、安曼[56],虽然Discretelladiscreta的分布是世界性,但很少有人建带。Discretelladiscreta带可以与Nv.w.eowageni对比。Yan et al.建议在缺少Nv.w.eowageni的情况下,可以用Discretelladiscreta作为划分印度阶与奥伦尼克阶的标志[54]。
在斯帕斯亚阶,有人建立了Pachycladina-Parachirognathus组合带[33,54],此组合带的时限可能较长,以属建带本身就是不可取的。因为对属的种群并不了解。此组合带内缺少Neospathodus,Novispathodus,Barinella和Neogondolella的种,可能是生态环境所致。Pachycladina-Parachirognathus组合带见于贵州关刀[33]、边阳[54]、广西太平[57]、西秦岑[58],西芷[59],此带在巢湖地区相当于Novispathoduswaageniwaageni带的上部和菊石Anasibirites带[48]以及贵州边阳剖面的Pachycladina-Parachirognathus组合带[54],也相当于湖北大峡口剖面的Novispathodusw.waageni带的上部,以及印度Spiti地区的Scythogondolellamilleri带[17,60]。Pachycladina和Parachirognathus都见于潮间带。但Pachycladina见于高能潮间带,而Parachirognathus见于低能潮间带。
贵州甲戎剖面Pachycladina-Parachirognathus见于斜坡之顶,可能也有下滑坡的再沉积。
王成源认为用Sc.milleri取代Pachycladina-Parachirognathus组合带,可能更好。
Orchard & Zonneveld依据加拿大Wapiti 湖区的资料,将斯密斯期的牙形刺自下而上建立4个带:B.nepalensis,S.Lachrymiformis,S.meeki,S.Mosheri, 后一个带,又分为2个亚带,S.phryna亚带在下,S.milleri亚带在上[51]。此方案目前还没有在世界上普遍使用,与现有的划分对比也不十分清楚。
斯帕斯晚期的牙形刺依据Orchard 和Zonneveld的总结,牙形刺分异度较低,各剖面差异较大,在高纬度地区, 有些Scythogondolella的种,如Sc.mostleri,Sc.Milleri,Nv.pingdinmgshanensis可以延伸到斯帕斯亚阶的顶部[51]。
斯帕斯中期的牙形刺依据Orchard & Zonneveld 的总结,是牙形刺发育的高峰期,常见的牙形刺有Ns.posterolongatus,Nv.waageni, 很多Neospathodus的种和Conservatella,Discretella,Guangxidella,Paullella,Scythogongdolella,Wapitiodus的种[51]。
斯帕斯早期的牙形刺,依据Orchard & Zonneveld的总结,Nv.waagenieowageni可以作为底界的标志,出现在界线附近的种包括Neospathoduscncavus,Ns.chii,Ns.postlongatus,Ns.pakistanensis,Borinellanepalensis,Eurygnathoduscostatus,而Ns.cristagali,Ns.dieneri时限较长,跨过印度阶和奥伦尼克阶的界线。Discretell,Spathicuspus?的种出现的稍晚些[51]。
印度阶印度阶分为迪耐尔亚阶(Dienertian)和格里斯巴钦亚阶(Griesbachian)(表4)。
Hindeodusparvus在浙江长兴煤山剖面的首现点位由王成源首先确定的,位于界线层2的中部[4,39,40]。即三叠系底界“金钉子”的位置。
Hindeodusparvus带在世界各地都有广泛分布[61--78], 既见于浅水相也见于深水相,是非常好的带化石。
但在长兴煤山剖面Hindeodusparvus带,仅见于27c,只有 4 cm厚,而且这4 cm厚的地层可能包含有Hindeodusparvus带和Hindeoduslobata带两个化石带,但是在该段地层中发现有硬底构造,这是该剖面所面临的新问题,这可能表明剖面有极小的沉积间断[79],但也存在不同的看法[80]。
1976年在中国发现了三叠纪牙形刺,确认了早、中、晚三叠世的8个牙形刺带的存在[81]。1991年中国三叠纪牙形刺生物带已划分为26个[82--84]。至2013年中国三叠纪牙形刺生物带又增至30个生物带, 寒武系至三叠系7个地质时代共划分出237个带[85,86]。目前三叠纪已划分出50个生物带,寒武系至三叠系共划分出257个生物带,增加了20个生物带,今后还有增加的可能。
5 结论
(1)三叠系3统7阶的时限差别极大,上三叠统时限最长,仅诺利阶的时限就比早三叠世和中三叠世时限之和还长。三叠世牙形刺生物地层牙形刺带的数量还有增加的可能,特别是晚三叠世。
(2)阶是全球的地质年代单位,应全球统一,使用共同的名称和定义。不要按国别建阶,更不能按岩相或生物群建阶。陆相地层不适宜建阶,中国三叠纪陆相地层建立的阶都应放弃不用。
(3)中国三叠系牙形刺生物带已划分出50个带,牙形刺在地层划分对比中起主导化石作用,越来越重要。
猜你喜欢
汉语世界(The World of Chinese)(2021年1期)2021-02-22 06:43:25
——工程地质勘察中,一种做交叉剖面的新方法
中国设备工程(2021年2期)2021-01-28 07:51:10
数学物理学报(2020年1期)2020-04-21 06:00:22
电子测试(2017年12期)2017-12-18 06:35:46
青海政报(2017年8期)2017-07-31 18:05:02
北京航空航天大学学报(2016年6期)2016-11-16 01:50:44
海峡姐妹(2016年7期)2016-02-27 15:21:26
地震研究(2014年3期)2014-02-27 09:30:50
