
温度和集胞藻浓度对轮虫生活史参数的影响
2019-04-04 02:48耿红姚慧张鹏宋发军林爱华孟艳艳张欢
中南民族大学学报(自然科学版) 2019年1期
耿红,姚慧,张鹏,宋发军,林爱华,孟艳艳,张欢
(1中南民族大学 生命科学学院,武陵山区特色资源植物种质保护与利用湖北省重点实验室,武汉430074;2中国科学院 水生生物研究所,武汉 430072)
湖泊富营养化是当今全球环境领域面临的突出问题[1],严重时可爆发蓝藻水华. 蓝藻水华会破坏水体食物链,导致生态系统崩溃. 轮虫是淡水生态系统中浮游动物的主要组成类群,在食物链中起着承上启下的枢纽作用[2]. 萼花臂尾轮虫(Brachionuscalyciflours)因繁殖快、世代周期短、易培养等特点被作为生态毒理学研究的模式生物[3]. 目前已有较多研究探讨蓝藻对浮游动物种群的影响,如浮游动物与蓝藻的摄食关系及其影响因素[4],铜绿微囊藻及其毒素对轮虫生活史的影响[5],铜绿微囊藻对浮游动物后代的耐受性影响[6]等,在这些研究中,多数集中于铜绿微囊藻及大型浮游动物种群的研究. 而有研究表明,蓝藻对浮游动物的影响会随蓝藻种类及浮游动物类群的不同而不同[7]. 集胞藻是单细胞非固氮蓝藻,生长周期短,易培养,不产生毒素,目前有关集胞藻对轮虫种群的影响研究几乎未见报道.
水体温度直接影响生物有机体的体温,体温的高低又决定了浮游动物新陈代谢、生物体的生长、发育、繁殖和行为等,从而影响浮游动物的丰度[8,9]. 全球变暖及其引起的湖泊水体温度升高已受到国际社会和学者的广泛关注[10],尽管已有研究表明温度对轮虫的个体大小、生长以及生活史特征有显著的影响,但是很少有研究关注温度与其他因子的交互作用对轮虫生长及繁殖影响;且少量的关于温度和食物浓度对轮虫种群增长及繁殖的研究报道结果却仍存在不一致性[11]. 本文以萼花臂尾轮虫(B.calyciflorus)为受试生物,采用动态生命表实验,研究不同温度下,不同浓度的集胞藻对轮虫种群生活史参数的影响,来探讨温度和集胞藻及其交互作用对轮虫生长繁殖的影响. 旨在丰富浮游动物与蓝藻间的相互关系,为臂尾轮虫属的生态参数研究提供基础数据资料.
1 材料与方法
1.1 轮虫的来源和培养
实验选用萼花臂尾轮虫为研究对象,萼花臂尾轮虫采自湖北省武汉市南湖水体,在实验室克隆培养半年以上,轮虫置于(25±1)℃恒温光照培养箱中进行群体培养,光照强度为2200 lx,光照周期为12L:12D,所用培养基配方为EPA[5].
1.2 藻种来源和培养
蛋白核小球藻(Chlorellapyrenoidosa)为培养轮虫所用的绿藻食物,实验所用蓝藻为集胞藻(Synechocystissp. PCC6803),实验所用的藻种均由中国科学院水生生物研究所淡水藻种库提供. 绿藻培养基为HB-4,蓝藻培养基为BG11[5],置于(25±1)℃的恒温光照培养箱中进行充气培养. 选取指数增长期的绿藻和蓝藻进行离心(6000 r/min,10 min)、显微镜下计数并用EPA培养基稀释到实验所需浓度.
1.3 动态生命表实验
实验设3个温度梯度,分别为15、25、35 ℃,每个温度下按照集胞藻浓度的不同再分别设4组,根据预实验结果,每组绿藻和蓝藻的浓度如下:第一组0 cells/mL集胞藻+105cells/mL蛋白核小球藻,第二组104cells/mL集胞藻+105cells/mL蛋白核小球藻,第三组105cells/mL集胞藻+105cells/mL蛋白核小球藻,第四组106cells/mL集胞藻+105cells/mL蛋白核小球藻.
实验前,将轮虫放置于15、25、35 ℃下预培养一周,同时将处于指数增长期的集胞藻和蛋白核小球藻进行离心、计数,放置4 ℃冰箱备用.
实验开始时,取带卵的非混交雌体,放置于烧杯中,在15、25、35 ℃下培养;选取龄长<3 h的轮虫幼体,分别放置于24孔板中进行生命表实验,每孔放1只轮虫. 总共有12组(3 温度 × 4 浓度),每组初始轮虫个数为18~23只. 每隔8 h观察一次,记录轮虫带卵数,母体存活数,新生幼体数,并将新生幼体从培养体系中移除,24 h更换一次EPA培养液,实验至全部母体死亡为止.
1.4 生活史参数计算方法
根据动态生命表实验数据计算轮虫主要生活史参数[13]:
特定年龄存活率lx:X年龄组开始时存活个体百分数;
特定年龄繁殖率mx:X年龄组平均每个个体所产的雌性后代数;
净生殖率R0:种群经过一个世代后的净增长率,R0=∑lxmx;
世代时间T:完成一个世代所需要的时间,T=∑lxmxx/R0;
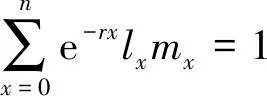
1.5 数据分析与处理
实验数据初步处理在Excel中进行,计算得到轮虫生活史参数之后,采用非参数双因素方差分析(Scheirer-Ray-Hare test)检验温度和集胞藻浓度对轮虫主要生活史参数的影响,利用多重比较(posthoc test)分析轮虫生活史参数在不同温度和不同浓度下各组别的差异,多重比较的结果用字母标注法标出,方差分析和多重比较均在R软件中完成.
2 结果与分析
2.1 萼花臂尾轮虫的存活率和繁殖率
随着温度升高,轮虫存活率开始下降的时间提前,存活率下降速度加快,轮虫的存活时间变短(图1). 集胞藻浓度对轮虫存活率的影响随温度的不同而有所不同,15 ℃时,轮虫的存活率呈现随集胞藻浓度的增加而升高的趋势;在25 ℃和35 ℃下,随集胞藻浓度上升,轮虫存活率反而明显下降. 温度对轮虫的繁殖率影响较大,在15 ℃下,轮虫的初次繁殖高峰期从第5 d开始,在25、35 ℃下轮虫的初次繁殖高峰期明显提前. 集胞藻浓度对轮虫繁殖率的影响也有一定的规律性,当温度为15 ℃时,轮虫繁殖率随集胞藻浓度的升高呈现增加的趋势,但当温度升高为25 ℃和35 ℃时,集胞藻浓度的增加不仅没有促进轮虫繁殖率的提高,反而抑制了轮虫的繁殖.

图1 3种温度下不同实验组萼花臂尾轮虫存活率(lx)和繁殖率(mx)曲线Fig.1 Age-specic survival rate (lx) and fecundity (mx) of Brachionus calyciflorus at three temperatures in different treatments.
2.2 萼花臂尾轮虫的生活史参数
当温度为15 ℃时,轮虫的净生殖率随集胞藻浓度的升高而增加,高浓度组(105、106cells/mL)轮虫的净生殖率显著高于对照组(0 cells/mL);当温度升高为25 ℃和35 ℃时,轮虫的净生殖率随集胞藻浓度的升高出现下降的趋势,其中105cells/mL集胞藻实验组轮虫的净生殖率显著低于对照组(图2). 比较相同集胞藻浓度下,轮虫在不同温度时的净生殖率变化,发现当集胞藻浓度较低(0、104cells/mL)时,轮虫的净生殖率随温度的升高而呈现增加的趋势;其中,集胞藻浓度为0 cells/mL时,15 ℃下轮虫的净生殖率最低;集胞藻浓度为104cells/mL时,25 ℃下轮虫的净生殖率最高;但当集胞藻浓度升高时(105、106cells/mL),轮虫的净生殖率随温度升高而呈现下降的趋势,当集胞藻浓度为106cells/mL时,35 ℃下轮虫的净生殖率显著低于对照组.

注:图中字母标注为多重比较的结果,不同字母表示不同处理组之间存在显著差异;以下同图2 3种温度下不同实验组萼花臂尾轮虫净生殖率变化Fig.2 Changes of net reproductive rate of Brachionus calyciflorus at three temperatures in different treatments
温度对轮虫的世代时间有显著影响,随温度的升高,轮虫世代时间明显缩短,25 ℃和35 ℃条件下,各实验组轮虫的世代时间都显著低于15 ℃. 相同温度下,不同实验组轮虫的世代时间也随集胞藻浓度的升高而呈下降趋势;当温度为15 ℃时,这种下降达到显著性水平,对照组轮虫的世代时间显著长于其他实验组轮虫(图3).
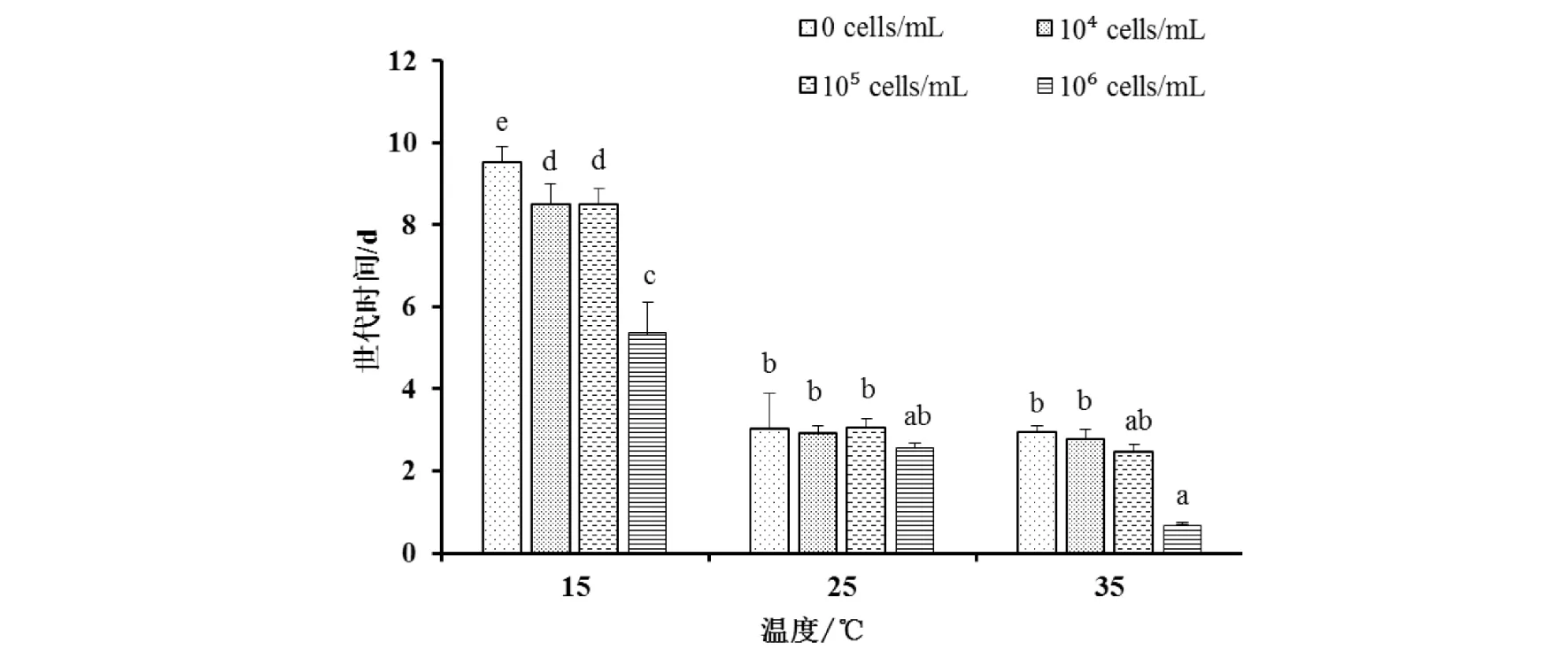
图3 3种温度下不同实验组萼花臂尾轮虫世代时间变化Fig.3 Changes of generation time of Brachionus calyciflorus at three temperatures in different treatments
图4显示了不同温度下,不同实验组轮虫的内禀增长率变化,当温度为15 ℃时,轮虫的内禀增长率随集胞藻浓度的升高而显著增加,与对照组(集胞藻浓度为0 cell/mL)相比,集胞藻浓度为104cells/mL时,轮虫的内禀增长率增加了67.4%,集胞藻浓度为105、106cells/mL时,轮虫的内禀增长率分别增加了119.6%、198.9%. 但当温度升高至25 ℃和35 ℃时,轮虫的内禀增长率随集胞藻浓度升高的变化呈现波动性,但仍能看出,轮虫的内禀增长率并没有随集胞藻浓度的升高而明显增加,当集胞藻浓度为105cells/mL时,轮虫的内禀增长率反而显著低于对照组,其余实验组轮虫的内禀增长率与对照组没有显著差异,35 ℃下,106cells/mL集胞藻浓度组的轮虫内禀增长率增加幅度最大,也只有10.2%,由此可见,随着温度升高,集胞藻对于轮虫内禀增长率的促进作用是明显减弱的.
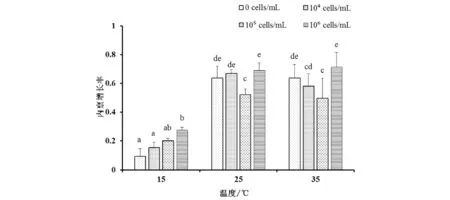
图4 3种温度下不同实验组萼花臂尾轮虫的内禀增长率变化Fig.4 Changes of intrinsic growth rate of Brachionus calyciflorus at three emperatures in different treatments
温度和集胞藻浓度对轮虫净生殖率(R0)、世代时间(T)和内禀增长率(rm)的方差分析结果见表1所示,温度、集胞藻浓度以及两者的交互作用对轮虫生活史主要参数都有极显著影响.

表1 温度与集胞藻浓度对轮虫生活史参数的方差分析Tab.1 Analysis of variance of temperature and Synechocystis concentration on lifeTable parameters of rotifer
注:*代表极显著,P<0.001
3 讨论
目前,虽然蓝藻对轮虫种群影响的机制尚不是很清楚,但一般认为,蓝藻主要通过产生蓝藻毒素[8,13]、形成群体或其他形态[7]以及低的营养价值[14]对轮虫种群产生影响. 有关蓝藻对轮虫种群的影响研究多集中于铜绿微囊藻上.有毒铜绿微囊藻所产生的微囊藻毒素对轮虫的生长繁殖有负面的抑制作用[13], 无毒微囊藻由于缺乏脂肪酸及其他脂类物质,也会使轮虫的生殖能力下降[14]. 而本研究结果则显示,当温度为15 ℃时,萼花臂尾轮虫种群的净生殖率和内禀增长率都随集胞藻浓度的升高而显著增加,表明在一定条件下,集胞藻可以为轮虫种群提供营养,是用于轮虫培养的潜在饲料[15],这也进一步说明蓝藻和轮虫间的相互关系随蓝藻种类的不同而不同[7],因此进行不同种类蓝藻对轮虫种群的影响研究是很有必要的.
温度是影响轮虫生活史特征的重要生态因子,本研究中,随温度的升高,轮虫的世代时间显著缩短,这与前人的研究结果是一致的[7,8]. 温度对轮虫净生殖率的影响随集胞藻浓度的不同而不同,当集胞藻浓度较低(0、104cells/mL)时,轮虫的净生殖率随温度的升高趋于增加;但当集胞藻浓度升高时,轮虫的净生殖率呈现相反的趋势. 由此看出,随温度的升高,集胞藻浓度对轮虫净生殖率的促进作用逐渐减弱,甚至出现抑制作用(当集胞藻浓度为106cells/mL时). 种群内禀增长率综合了特定年龄存活率和繁殖率、首次生殖年龄和生殖间隔等,因此,温度常对其具有显著影响[11],这在本研究中也得到了证实. 实验中轮虫种群的内禀增长率随温度的升高而增加,但增加的程度则随温度和集胞藻浓度的不同而不同. 以106cells/mL的集胞藻浓度为例,与对照组相比,15 ℃下轮虫种群的内禀增长率增加高达198.9%;而当温度升高到25 ℃和35 ℃,与对照组相比,轮虫种群的内禀增长率增加只有8.5%和12.1%. 由此可见,温度的升高降低了集胞藻对轮虫种群的促进作用. 在实验过程中,曾观察到较高温度下集胞藻的聚集现象.可能是由于在高温度下,集胞藻的高密度聚集,对轮虫的摄食活动造成机械干扰[7],从而影响轮虫种群的净生殖率和内禀增长率. 温度升高对水体浮游动物的实际影响将取决于整个水体生态系统不同环境因素之间复杂的相互作用,考虑到浮游动物在水生食物网中的重要性以及人们对全球气候变暖的持续关注,显然需要进一步研究温度与其他生态变量的综合作用对轮虫及其他浮游动物种群的影响.
猜你喜欢
发明与创新·中学生(2022年3期)2022-03-14
伦理学研究(2021年6期)2021-12-29
科学养鱼(2020年10期)2020-11-23
伦理学研究(2020年4期)2020-09-07
科学养鱼(2020年12期)2020-04-22
课外语文·中(2020年2期)2020-03-24
知识窗(2019年4期)2019-04-26
农民致富之友(2016年24期)2017-04-26
海洋世界(2016年2期)2016-03-11
科学24小时(2015年3期)2015-09-10
