
铁橡栎的潜在分布区及其对气候变化的响应
2019-04-02 06:45:10刘瑞斌姜小龙戴锡玲
植物资源与环境学报 2019年1期
刘瑞斌, 姜小龙, 戴锡玲, 邓 敏,①
〔1. 上海师范大学生命科学学院, 上海 200234; 2. 上海辰山植物园(中国科学院上海辰山植物科学研究中心), 上海 201602〕
中国西南地区为同纬度生物多样性最高的区域[1-2]。该区域板块运动活跃、气候复杂多变,是保存孑遗物种的“博物馆”和植物区系分化的“摇篮”[1-3]。新近纪以来的气候变化,特别是第四纪冰期-间冰期循环,导致全球生物分布格局明显改变。基于孢粉对古气候和古植被重建的研究结果显示:末次盛冰期(last glacial maximum)草原植被扩张至现代的常绿阔叶林区的北部,常绿阔叶林则退缩至现代的热带区域,同时伴随着热带植被类型的消失。进入相对温暖和湿润的全新世中期(mid-holocene),东亚暖温性森林分布区域的纬度向北推进了约4°[4]。过去的气候变化对现代亚热带植被的分布具有深远影响[5]。在未来全球持续升温的背景下,中国西南地区增温幅度较大,降水量却未明显增加[6]。这种气候条件对中国西南干热河谷的植被提出了新的挑战。加之河谷区海拔一般较低,适于耕作,土地已被高强度利用,并且由于大规模水电站的建设,使中国西南干热河谷植被受到严重破坏。目前,有关中国西南干热河谷物种分布动态的研究报道较少,因此,研究干热河谷代表类群的分布格局对地质时期气候变化的响应,可为未来全球气候变化下该物种如何应答提供重要线索,并为干热河谷生态系统的保护和恢复提供重要基础资料。
目前,物种分布区模型已广泛运用到多个研究领域,如生态学、谱系地理学以及保护生物学。虽然物种分布预测已发展出多种模型,如BIOCLIM[7]、DOMAIN[8]、GARP[9]和MaxEnt[10-11],但BIOCLIM模型忽略了生物气候变量间的关联性和等效性,需要增加生物气候变量才能准确模拟物种分布[7];在生物气候变量较少时,DOMAIN模型模拟效果较好,但在区分存在与否的临界阈值时有一定误差[12]。相比之下,MaxEnt模型仅依赖于物种分布数据,从符合条件的分布中选择熵最大的分布作为最优分布,首先确定特征空间,即物种已知分布区域,然后寻找限制物种分布的约束条件(生物气候变量),构筑约束集合,最后建立二者之间的相互关系,因此,MaxEnt模型是目前应用最广、预测效果较好的模型[10,13-14]。
铁橡栎(QuercuscocciferoidesHand.-Mazz.)隶属于壳斗科(Fagaceae)栎属(QuercusLinn.),为常绿小乔木或灌木,主要分布在中国西南地区(北纬22°~30°、东经97°~105°)海拔600~2 500 m的干热河谷[15],具有耐旱、耐贫瘠以及适应性较强等特点,是中国西南广义干热河谷特殊植被类型的建群种和优势树种。其种子主要通过重力或啮齿类动物传播,传播距离十分有限。在当前干热河谷植被遭受破坏日益加剧的情况下,了解过去以及未来气候波动下铁橡栎的分布动态以及限制其分布的主导生物气候变量,可为该物种的种质资源保护及生态恢复研究提供科学依据。
本研究在对铁橡栎分布数据全面收集和整理的基础上,基于MaxEnt模型模拟铁橡栎的潜在分布范围,并分析影响铁橡栎分布的主导生物气候变量,以期揭示以铁橡栎为代表的干热河谷植物的分布演化格局,为这一特殊生态系统的保护和修复提供重要指导和基础资料。
1 材料和方法
1.1 分布记录获取
铁橡栎的分布记录来源包括本研究组的野外采集记录、全球生物多样性信息网(GBIF,http:∥www.gbif.org)、中国数字植物标本馆(CVH,http:∥www.cvh.org.cn)以及密苏里植物园标本馆。剔除重复、鉴定错误及分布记录不详的数据,最终获得91条准确的分布记录(即91个分布点),其中,21条来自本研究组野外实地采集记录,其余70条来自以上网站和标本馆(表1)。
1.2 生物气候变量的下载与筛选
从世界气候网站(http:∥worldclim.org)下载现代(1950年至2000年)以及采用通用气候系统模式CCSM4模拟生成的末次盛冰期、全新世中期和未来(2070年)4个时间段的19个生物气候变量,数据的空间分辨率均为2.5′。其中,2070年的气候数据基于温室气体排放情景为典型浓度目标8.5(RCP8.5)模型下获得。
为了避免高度相关的生物气候变量造成模型的过拟合现象[16],利用Dismo (http:∥CRAN.R-project.org/package=dismo)将19个与温度和降水有关的生物气候变量进行了相关性分析。剔除相关系数大于0.8的生物气候变量,最终筛选出年均温(bio1)、昼夜温差月均值(bio2)、等温性(bio3)、温度季节变化(bio4)、年均降水量(bio12)、最干月份降水量(bio14)和最热季度降水量(bio18)7个生物气候变量。随后,利用R语言基础包绘制各分布点现代气候数据的频率直方图及正态分布曲线。
表1用于MaxEnt模型预测的中国西南干热河谷铁橡栎的分布记录
Table1DistributionrecordsofQuercuscocciferoidesHand.-Mazz.indry-hotvalleyofSouthwestChinausedforMaxEntmodelprediction
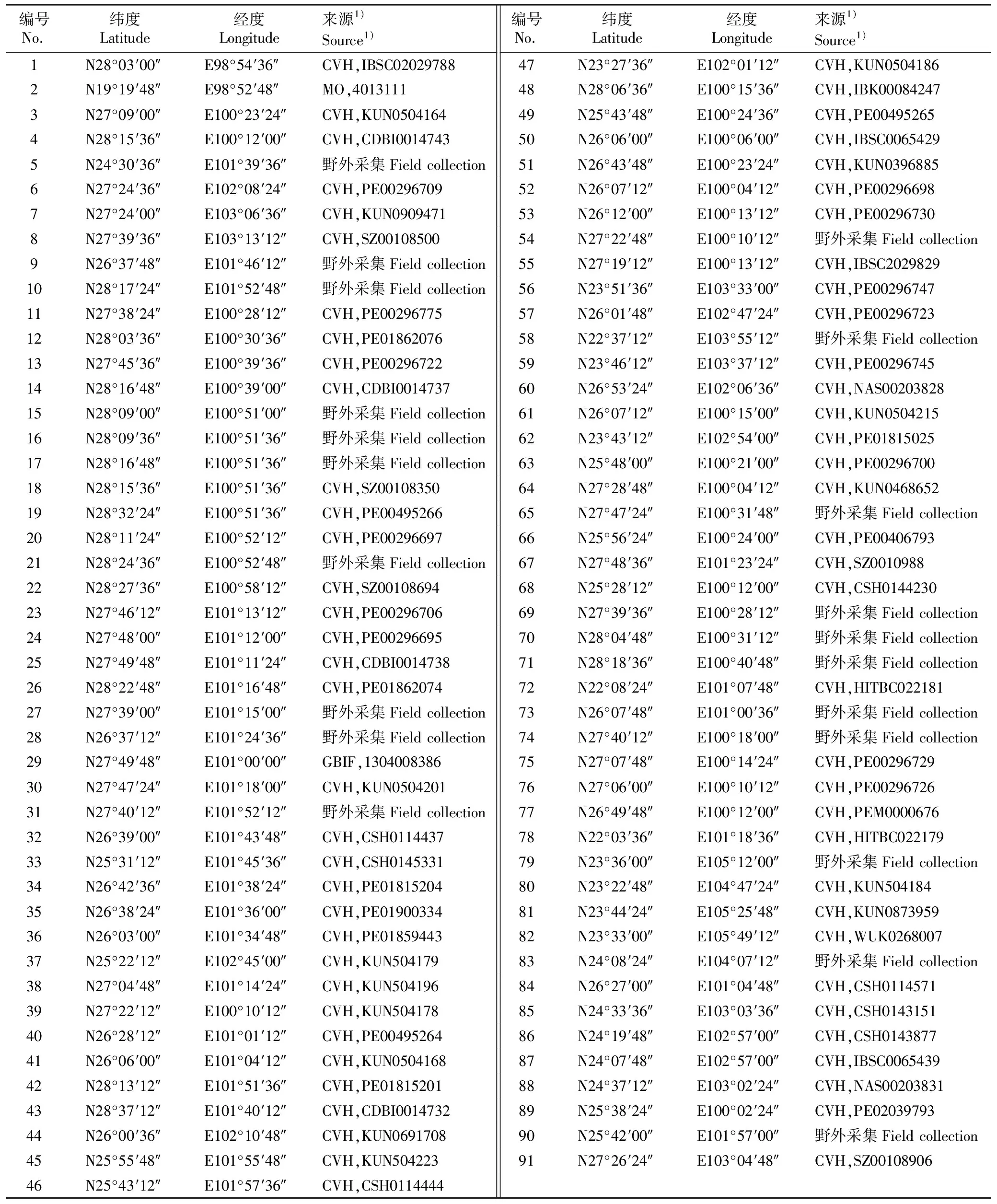
编号No.纬度Latitude经度Longitude来源1)Source1)编号No.纬度Latitude经度Longitude来源1)Source1)1N28°03′00″E98°54′36″CVH,IBSC0202978847N23°27′36″E102°01′12″CVH,KUN05041862N19°19′48″E98°52′48″MO,401311148N28°06′36″E100°15′36″CVH,IBK000842473N27°09′00″E100°23′24″CVH,KUN050416449N25°43′48″E100°24′36″CVH,PE004952654N28°15′36″E100°12′00″CVH,CDBI001474350N26°06′00″E100°06′00″CVH,IBSC00654295N24°30′36″E101°39′36″野外采集Field collection51N26°43′48″E100°23′24″CVH,KUN03968856N27°24′36″E102°08′24″CVH,PE0029670952N26°07′12″E100°04′12″CVH,PE002966987N27°24′00″E103°06′36″CVH,KUN090947153N26°12′00″E100°13′12″CVH,PE002967308N27°39′36″E103°13′12″CVH,SZ0010850054N27°22′48″E100°10′12″野外采集Field collection9N26°37′48″E101°46′12″野外采集Field collection55N27°19′12″E100°13′12″CVH,IBSC202982910N28°17′24″E101°52′48″野外采集Field collection56N23°51′36″E103°33′00″CVH,PE0029674711N27°38′24″E100°28′12″CVH,PE0029677557N26°01′48″E102°47′24″CVH,PE0029672312N28°03′36″E100°30′36″CVH,PE0186207658N22°37′12″E103°55′12″野外采集Field collection13N27°45′36″E100°39′36″CVH,PE0029672259N23°46′12″E103°37′12″CVH,PE0029674514N28°16′48″E100°39′00″CVH,CDBI001473760N26°53′24″E102°06′36″CVH,NAS0020382815N28°09′00″E100°51′00″野外采集Field collection61N26°07′12″E100°15′00″CVH,KUN050421516N28°09′36″E100°51′36″野外采集Field collection62N23°43′12″E102°54′00″CVH,PE0181502517N28°16′48″E100°51′36″野外采集Field collection63N25°48′00″E100°21′00″CVH,PE0029670018N28°15′36″E100°51′36″CVH,SZ0010835064N27°28′48″E100°04′12″CVH,KUN046865219N28°32′24″E100°51′36″CVH,PE0049526665N27°47′24″E100°31′48″野外采集Field collection20N28°11′24″E100°52′12″CVH,PE0029669766N25°56′24″E100°24′00″CVH,PE0040679321N28°24′36″E100°52′48″野外采集Field collection67N27°48′36″E101°23′24″CVH,SZ001098822N28°27′36″E100°58′12″CVH,SZ0010869468N25°28′12″E100°12′00″CVH,CSH014423023N27°46′12″E101°13′12″CVH,PE0029670669N27°39′36″E100°28′12″野外采集Field collection24N27°48′00″E101°12′00″CVH,PE0029669570N28°04′48″E100°31′12″野外采集Field collection25N27°49′48″E101°11′24″CVH,CDBI001473871N28°18′36″E100°40′48″野外采集Field collection26N28°22′48″E101°16′48″CVH,PE0186207472N22°08′24″E101°07′48″CVH,HITBC02218127N27°39′00″E101°15′00″野外采集Field collection73N26°07′48″E101°00′36″野外采集Field collection28N26°37′12″E101°24′36″野外采集Field collection74N27°40′12″E100°18′00″野外采集Field collection29N27°49′48″E101°00′00″GBIF,130400838675N27°07′48″E100°14′24″CVH,PE0029672930N27°47′24″E101°18′00″CVH,KUN050420176N27°06′00″E100°10′12″CVH,PE0029672631N27°40′12″E101°52′12″野外采集Field collection77N26°49′48″E100°12′00″CVH,PEM000067632N26°39′00″E101°43′48″CVH,CSH011443778N22°03′36″E101°18′36″CVH,HITBC02217933N25°31′12″E101°45′36″CVH,CSH014533179N23°36′00″E105°12′00″野外采集Field collection34N26°42′36″E101°38′24″CVH,PE0181520480N23°22′48″E104°47′24″CVH,KUN50418435N26°38′24″E101°36′00″CVH,PE0190033481N23°44′24″E105°25′48″CVH,KUN087395936N26°03′00″E101°34′48″CVH,PE0185944382N23°33′00″E105°49′12″CVH,WUK026800737N25°22′12″E102°45′00″CVH,KUN50417983N24°08′24″E104°07′12″野外采集Field collection38N27°04′48″E101°14′24″CVH,KUN50419684N26°27′00″E101°04′48″CVH,CSH011457139N27°22′12″E100°10′12″CVH,KUN50417885N24°33′36″E103°03′36″CVH,CSH014315140N26°28′12″E101°01′12″CVH,PE0049526486N24°19′48″E102°57′00″CVH,CSH014387741N26°06′00″E101°04′12″CVH,KUN050416887N24°07′48″E102°57′00″CVH,IBSC006543942N28°13′12″E101°51′36″CVH,PE0181520188N24°37′12″E103°02′24″CVH,NAS0020383143N28°37′12″E101°40′12″CVH,CDBI001473289N25°38′24″E100°02′24″CVH,PE0203979344N26°00′36″E102°10′48″CVH,KUN069170890N25°42′00″E101°57′00″野外采集Field collection45N25°55′48″E101°55′48″CVH,KUN50422391N27°26′24″E103°04′48″CVH,SZ0010890646N25°43′12″E101°57′36″CVH,CSH0114444
1)CVH: 中国数字植物标本馆,其后编号为标本的馆藏条形码Chinese Virtual Herbarium, subsequent code represents bar code of specimen; MO: 密苏里植物园标本馆,其后编号为标本的馆藏流水号Missouri Botanical Garden Herbarium, subsequent code represents serial No. of specimen; GBIF: 全球生物多样性信息网,其后编号为标本的馆藏条形码Global Biodiversity Information Facility, subsequent code represents bar code of specimen.
1.3 模型预测及准确性评价
采用MaxEnt 3.3.3k软件对铁橡栎不同时期的潜在分布区进行模拟。为提高模型预测准确性,随机选取75%的分布数据作为训练数据集(training data),剩余的分布数据作为测试数据集(testing data)。选择刀切法(jackknife),设置10次重复运算的平均预测结果作为本研究的预测结果,其他参数采用软件默认设置。利用ArcGIS v10.0.0(http:∥www.esri.com/software/arcgis/)对模型预测结果进行适生等级划分及可视化处理,参照文献[17]的方法,根据适宜度(S)将适宜区划分为4个等级:0%≤S<25%,不适宜区;25%≤S<50%,低度适宜区;50%≤S<75%,中度适宜区;75%≤S≤100%,高度适宜区。利用Arctoolbox中的重分类(reclassify)统计各级栅格数量。采用MaxEnt 3.3.3k软件绘制受试者工作特征(receiver operating characteristic,ROC)曲线检验模型准确性。ROC曲线下的面积(area under receiver operating characteristic curve,AUC)表示模型的预测能力。AUC值的取值范围为[0.5,1.0],AUC值越大,模型预测能力越强,衡量标准为:0.7≤AUC<0.8,较准确;0.8≤AUC<0.9,很准确;0.9≤AUC≤1.0,极准确。通过最大训练敏感度加特异性逻辑值作为物种缺失/存在阈值来确定物种分布,并采用栖息地稳定性(NStab)、栖息地分布面积比(Na)和栖息地扩张程度(Ne)3个指标比较末次盛冰期以来铁橡栎分布范围的变化。其中,NStab的计算公式为NStab=1-|NPre-N|,式中,NPre表示分布点在现代的栖息地适宜性,N表示分布点在末次盛冰期或未来的栖息地适宜性;Na表示分布面积变化的程度,计算公式为Na=现代的分布面积/过去的分布面积,Na值接近1表示物种具有稳定的分布,远高于或低于1表示物种分布区经历了从过去到现代的扩张或收缩;Ne表示物种分布区从过去到现代的扩张百分率,计算公式为Ne=〔1-(过去与现代生态位重叠面积/现代分布面积)〕×100%。
1.4 环境因子重要性评价
综合使用贡献率(contribution rate)、置换重要值(permutation importance)和Jackknife检验来分析生物气候变量对铁橡栎现代分布格局的重要性。
2 结果和分析
2.1 MaxEnt模型预测的准确性评估
MaxEnt模型对铁橡栎现代潜在分布区的预测结果显示:训练AUC值为0.995±0.000,测试AUC值为0.994±0.002,表明MaxEnt模型对铁橡栎现代潜在分布区的预测极准确。
2.2 铁橡栎现代潜在分布区
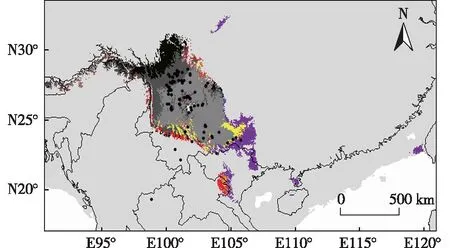
预测结果(图1)表明:铁橡栎现代(1950年至2000年)潜在分布区主要位于滇中高原、滇北—川南金沙江流域和滇东南南盘江流域。91个铁橡栎分布点中,大部分位于高度适宜区,少量位于中度和低度适宜区,仅有2个分布点位于不适宜区。其高度适宜区主要位于滇中高原和滇北—川南金沙江流域,约占适宜区总面积的16.98%。中度适宜区主要位于滇西南、滇东南和川南,约占适宜区总面积的19.50%。
此外,物种分布模型分析结果(表2)表明:编号2的铁橡栎分布点在末次盛冰期、现代和未来的栖息地适宜性,编号58的铁橡栎分布点在现代和未来的栖息地适宜性,编号72的铁橡栎分布点在末次盛冰期、现代和未来(2070年)的栖息地适宜性,以及编号89的铁橡栎分布点在现代和未来的栖息地适宜性均低于现代物种存在阈值(0.103±0.016)。

: 不适宜区Unsuitable area; : 低度适宜区Low suitable area; : 中度适宜区Moderate suitable area; : 高度适宜区High suitable area. ●: 铁橡栎分布点Distribution point of Quercus cocciferoides Hand.-Mazz.
表2基于MaxEnt模型获得的铁橡栎各分布点在不同时期的栖息地适宜性和栖息地稳定性1)
Table2HabitatsuitabilityandhabitatstabilityofeachdistributionpointofQuercuscocciferoidesHand.-Mazz.duringdifferentperiodsbasedonMaxEntmodel1)

编号No.NLGMNPreNFNStab(LGM)NStab(F)编号No.NLGMNPreNFNStab(LGM)NStab(F)10.130.530.610.600.93470.280.360.230.920.8720.050.010.000.961.00480.140.700.680.440.9830.470.700.700.781.00490.490.580.590.910.9940.070.570.680.500.90500.330.680.560.650.8850.540.600.320.940.72510.470.700.700.781.0060.600.630.530.970.91520.460.590.580.870.9970.300.440.370.870.93530.320.590.430.730.8480.300.400.470.900.93540.370.700.680.670.9890.400.670.560.730.89550.270.690.580.580.88100.540.640.590.900.95560.420.480.410.940.93110.310.710.690.600.98570.660.680.650.980.97120.180.710.680.470.97580.310.080.040.770.96130.290.680.660.610.98590.440.680.450.760.76140.110.650.640.471.00600.630.640.590.990.95150.180.670.640.510.97610.390.480.390.910.91160.180.670.640.510.97620.560.620.380.950.77170.140.660.630.480.97630.490.580.590.910.99180.140.660.630.480.97640.480.720.700.770.98190.070.540.560.530.98650.170.590.600.590.99200.170.660.630.510.97660.510.590.590.921.00210.110.600.580.510.98670.480.620.580.860.96220.100.560.520.540.96680.500.580.580.921.00230.430.650.610.780.96690.310.710.690.600.98240.430.700.660.730.96700.180.710.680.470.97250.430.700.660.730.96710.110.630.630.471.00260.180.650.600.530.95720.060.080.040.990.96270.460.620.590.840.97730.580.610.610.970.99280.180.700.400.480.70740.270.720.700.550.98290.320.610.590.710.98750.420.670.670.751.00300.460.700.660.760.96760.300.530.540.770.98310.550.650.580.900.93770.530.710.680.820.98320.400.670.560.730.89780.160.140.060.980.91330.340.680.490.670.81790.500.330.330.831.00340.620.670.580.950.92800.540.370.380.830.98350.240.700.410.540.70810.510.360.250.850.89360.190.580.300.620.72820.490.280.130.800.85370.200.660.350.540.69830.440.580.520.860.93380.500.630.620.870.99840.210.690.550.510.86390.370.710.700.650.98850.620.640.500.980.86400.150.700.450.450.75860.620.630.530.990.90410.490.550.520.930.96870.550.630.530.920.90420.540.670.580.870.91880.620.640.500.980.86430.290.590.560.700.97890.260.100.100.841.00440.650.650.621.000.97900.300.320.400.970.92450.430.680.550.750.87910.350.510.430.840.92460.300.320.400.970.92
1)NLGM: 铁橡栎分布点在末次盛冰期的栖息地适宜性Habitat suitability of distribution point ofQuercuscocciferoidesHand.-Mazz. in the last glacial maximum;NPre: 铁橡栎分布点在现代的栖息地适宜性Habitat suitability of distribution point ofQ.cocciferoidesat present;NF: 铁橡栎分布点在未来(2070年)的栖息地适宜性Habitat suitability of distribution point ofQ.cocciferoidesin the future (in 2070);NStab(LGM): 铁橡栎分布点在末次盛冰期的栖息地稳定性Habitat stability of distribution point ofQ.cocciferoidesin the last glacial maximum;NStab(F): 铁橡栎分布点在未来的栖息地稳定性Habitat stability of distribution point ofQ.cocciferoidesin the future.
2.3 铁橡栎过去和未来的潜在分布区
MaxEnt模型预测的铁橡栎在不同时期的潜在分布区见图2。结果显示:末次盛冰期铁橡栎的高度适宜区主要位于中国的滇中和滇东南,中度和低度适宜区则主要位于缅甸东北部,越南北部,以及中国的滇西南、广西大部、海南岛及广东以东的南海大陆架(图2-A)。该时期铁橡栎高度适宜区的面积较现代减少30.31%,栖息地分布面积比(Na)和栖息地扩张程度(Ne)分别为1.29和44.91%,呈收缩的迹象(图3)。
全新世中期中度和高度适宜区的范围与现代的中度和高度适宜区的范围基本一致,但此时期滇东南和华南不再适宜铁橡栎分布(图2-B)。该时期的Na和Ne值分别为1.09和18.53%,铁橡栎的分布范围显现出相对稳定的状态。
未来(2070年)高度适宜区的范围与现代高度适宜区的范围十分相似(图2-C)。该时期的Na和Ne值分别为0.99和19.29%,铁橡栎的中度和低度适宜区呈轻微向北扩张的迹象(图3)。
: 现代潜在分布区Potential distribution area at present; : 末次盛冰期潜在分布区Potential distribution area in the last glacial maximum; : 未来(2070年)潜在分布区Potential distribution area in the future (in 2070); : 现代和末次盛冰期的重叠潜在分布区Overlapping-potential distribution area at present and in the last glacial maximum; : 现代和未来的重叠潜在分布区Overlapping-potential distribution area at present and the future; : 末次盛冰期和未来的重叠潜在分布区Overlapping-potential distribution area in the last glacial maximum and the future; : 现代、末次盛冰期和未来的重叠潜在分布区Overlapping-potential distribution area at present and in the last glacial maximum and the future.
此外,物种分布模型分析结果(表2)表明:编号2的铁橡栎分布点在末次盛冰期、现代和未来的栖息地适宜性,编号4和19的铁橡栎分布点在末次盛冰期的栖息地适宜性,以及编号72的铁橡栎分布点在末次盛冰期、现代和未来的栖息地适宜性均低于末次盛冰期物种存在阈值(0.091±0.035);编号2的铁橡栎分布点在末次盛冰期、现代和未来的栖息地适宜性,编号58的铁橡栎分布点在现代和未来的栖息地适宜性,编号72的铁橡栎分布点在末次盛冰期、现代和未来的栖息地适宜性,编号78和82的铁橡栎分布点在未来的栖息地适宜性,以及编号89的铁橡栎分布点在现代和未来的栖息地适宜性均低于未来物种存在阈值(0.193±0.009)。末次盛冰期各分布点的栖息地稳定性变幅较大,未来各分布点的栖息地稳定性则变幅较小。
2.4 生物气候变量的重要性分析
结果(表3)表明:用于MaxEnt模型预测的影响铁橡栎现代分布的7个生物气候变量中,贡献率位列前4的生物气候变量依次为最干月份降水量(29.6%)、昼夜温差月均值(27.4%)、等温性(23.5%)、年均温(18.8%),四者的贡献率总和为99.3%。置换重要值位列前3的生物气候变量依次为等温性(41.5%)、最干月份降水量(29.4%)、昼夜温差月均值(26.9%),三者的总和超过97%。
Jackknife检验结果(图4)表明:当分别使用单独生物气候变量和使用除该生物气候变量外的所有生物气候变量时,正规化训练增益、测试增益和受试者工作特征曲线下的面积得到的结果相同,即等温性、最干月份降水量和昼夜温差月均值均位列前3,表明这3个生物气候变量影响铁橡栎分布范围的贡献率较高。
表3用于MaxEnt模型预测的影响铁橡栎现代分布的生物气候变量的贡献率和置换重要值
Table3ContributionrateandpermutationimportanceofbioclimaticvariablesaffectingdistributionofQuercuscocciferoidesHand.-Mazz.atpresentusedforMaxEntmodelprediction
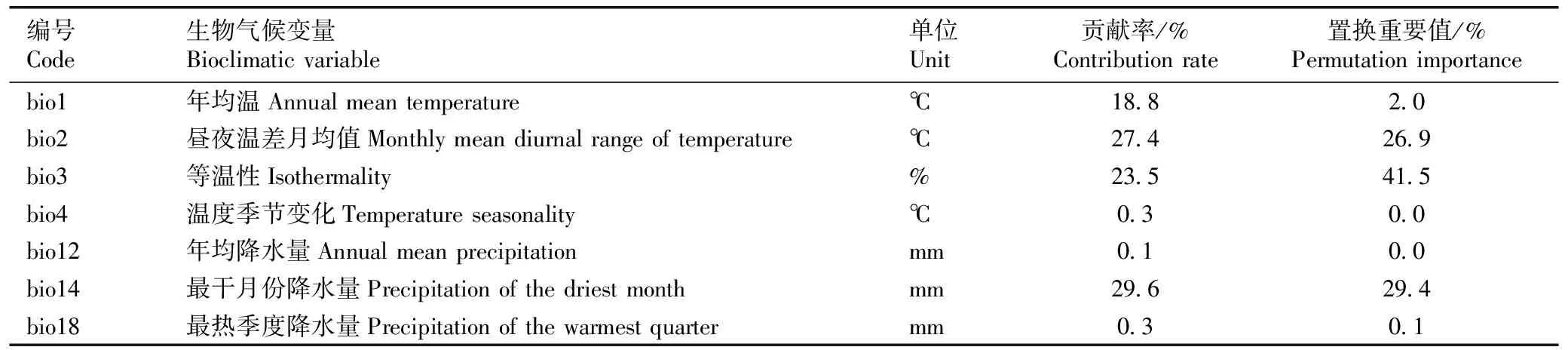
编号Code生物气候变量Bioclimatic variable单位Unit贡献率/%Contribution rate置换重要值/%Permutation importancebio1年均温Annual mean temperature℃18.82.0bio2昼夜温差月均值Monthly mean diurnal range of temperature℃27.426.9bio3等温性Isothermality%23.541.5bio4温度季节变化Temperature seasonality℃0.30.0bio12年均降水量Annual mean precipitationmm0.10.0bio14最干月份降水量Precipitation of the driest monthmm29.629.4bio18最热季度降水量Precipitation of the warmest quartermm0.30.1
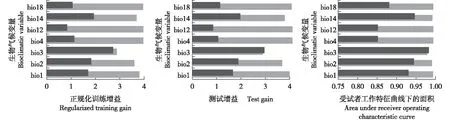
深灰色和灰色条形图分别表示使用单独生物气候变量和除该生物气候变量外的所有生物气候变量的MaxEnt模型预测结果Dark gray and gray bars represent prediction results of running MaxEnt model with only bioclimatic variable and all bioclimatic variables except this bioclimatic variable, respectively. bio1: 年均温Annual mean temperature; bio2: 昼夜温差月均值Monthly mean diurnal range of temperature; bio3: 等温性Isothermality; bio4: 温度季节变化Temperature seasonality; bio12: 年均降水量Annual mean precipitation; bio14: 最干月份降水量Precipitation of the driest month; bio18: 最热季度降水量Precipitation of the warmest quarter.
综合上述分析结果显示:影响铁橡栎现代潜在分布的主要生物气候变量为气温因子(等温性、昼夜温差月均值和年均温)和降水因子(最干月份降水量)。
2.5 响应曲线分析
本研究选取的7个生物气候变量的边缘响应曲线见图5,其频率直方图和正态分布曲线见图6。结果表明:适宜铁橡栎生长的年均温的变化范围为10.7 ℃~20.5 ℃,昼夜温差月均值的变化范围为10.9 ℃~13.5 ℃,最干月份降水量的变化范围为3~14 mm,最热季度降水量的变化范围为400~600 mm,年均降水量的变化范围为800~1 100 mm。

等温性和温度季节变化以数值的100倍为基础Isothermality and temperature seasonality are based on 100 times of their value. : 10次重复的平均值Mean of ten repeats; : 标准偏差Standard deviation.

等温性和温度季节变化均以数值的100倍为基础Isothermality and temperature seasonality are based on 100 times of their value.
3 讨 论
3.1 铁橡栎的历史分布动态
第四纪冰期-间冰期的气候回旋对铁橡栎种群的分布格局产生了一定影响。在末次盛冰期,虽然中国没有形成大陆型冰川,但植物区系也受到影响[18]。这一时期,夏季风明显减弱,冬季风增强,致使中纬度地区平均降温12 ℃~15 ℃,这可能导致了铁橡栎自西北向东南退却,东缘至中国广西西部,南部分布区延伸至中国云南与越南交界处。进入全新世中期,气候重新变得温暖湿润,基于孢粉数据对生物群落的重建表明,该时期的气候特征和亚热带植被与现代十分相似[19]。另外,常绿硬叶栎类和栲类植物的孢粉量有所增多[20-21],推测铁橡栎可能在此时期恢复生长或向北扩张。第四纪以来青藏高原持续抬升,加强了西南季风对中国西南地区的影响,该季风携带的水汽导致局部区域降水量增加,这可能是滇东南不再适宜铁橡栎生长的主要原因。中国西南地区南北地势的差异可能阻碍了滇东南群体的北迁,经过长期的自然选择,该区域的群体形成了适应性进化。
本研究利用MaxEnt模型预测铁橡栎的现代潜在分布区主要集中于滇中高原、滇北—川南金沙江流域和滇东南南盘江流域。铁橡栎作为中国西南干热河谷的典型植被的代表,其分布受河谷气候的影响较大。由于受河谷地形和地理纬度的影响,中国西南干热河谷气候异常干热,干湿季分明,降水量较少且蒸发量大,但光、热资源丰富[22]。这种温度变异性低和较干旱的环境可能较适宜铁橡栎的生长。
综上所述,推测从末次盛冰期至现代,铁橡栎地理分布范围经历了“向南退缩—向北扩张—基本稳定”的过程。滇东南和中南半岛北部地区可能为铁橡栎提供了多个分散的避难所,中国西南干热河谷可能是铁橡栎在冰期-间冰期迁移和扩散的重要通道。未来全球气温和降水都将有所增加[23],铁橡栎的潜在分布范围也可能有所增加。但中国西南干热河谷大多海拔较低,是主要的农耕区[24],铁橡栎群体的生境斑块化明显,受人为干扰较大,因此,该区域干热河谷原生植被在环境适宜的条件下仍可能会衰退和减少,物种生存风险仍将会加剧。
3.2 气候因子的制约作用
植物的地理分布主要受环境因子制约,其中,温度和降水是影响植物地理分布的主要决定因子。中国西南干热河谷冬季和春季受西风南支控制形成干季,而在夏季受印度洋夏季风影响,尽管带来大量水汽,但受纬度和河谷地形的影响,该地区形成了降水量极少、干湿季分明、异常干热的气候特征[22]。但干热河谷特有种对该生境的适应性良好,如:滇榄仁(TerminaliafranchetiiGagnep.)一定程度上在干热的气候条件下种子质量更好,并通过增加有效种子数和种子萌发能力来满足天然更新的需要,体现了干热河谷物种对干热生境的特殊适应[25]。虽然大量栎属植物的生理生态研究结果表明:高温干旱生境会在一定程度上限制栎属植物的生殖、发育和地理分布[17,26-29],但铁橡栎出对干热河谷气候的适应性良好,可能其生长节律与其他生长于中生型生境的栎属植物的适应性存在差异。本研究认为,等温性是影响铁橡栎潜在分布的首要因子。根据筛选生物气候变量的相关性分析可知:等温性与昼夜温差月均值显著相关(相关系数0.70,P<0.05),二者均可作为年温度变化趋势的指示参数。此外,推测滇中高原北部地区的冬季低温可能是限制铁橡栎分布的重要因子。
3.3 干热河谷生态系统的保护
本研究结果表明:滇东南和中南半岛北部地区可能是干热河谷植被冰期的避难所,这一区域的群体可能具有较高的遗传多样性和特殊遗传基因,并且这一区域群体所处的生境与核心分布区明显不同,环境因子已有明显分化,这一区域群体可能存在与核心分布区群体不同的适应性,因此,对滇东南及中南半岛北部的河谷应给予重点保护。在未来气候条件下,铁橡栎的分布虽然可能出现轻微的扩张趋势,但是中国西南干热河谷已遭到较严重的破坏,因此,在代表区域(如滇中高原和滇东南)选取植被保存完好的区域进行保护对干热河谷的生物多样性保护十分重要。
3.4 结论和展望
本研究结果表明:气温因子(等温性、昼夜温差月均值和年均温)和降水因子(最干月份降水量)共同制约铁橡栎的现代分布范围,其中气温因子的制约作用尤为突出,并且铁橡栎在不同地质时期的分布范围变化明显。末次盛冰期,铁橡栎的整个高度适宜区向南撤退,现代的滇北和川南地区在该时期并适宜铁橡栎生长。而进入全新世中期,铁橡栎的分布区则呈现向北回迁和扩张的态势,逐渐演变为现代的模式。虽然,在未来全球变暖的背景下,铁橡栎适宜区可能局部扩张,但由于适生区大部分位于农垦区,人类活动频繁,土地利用强度较高,铁橡栎的分布仍面临很大挑战。另外,零星分布个别群体的滇东南可能是铁橡栎在末次盛冰期的南部避难所。综合上述,在对中国西南干热河谷分布的铁橡栎进行保护的同时,应对滇东南的种群给予重点保护。
目前,在分子层面上探索气候变化对物种遗传结构的影响较为有效,建议结合高通量分子标记和分布区模拟开展群体遗传学研究,揭示气候变化和环境适应对铁橡栎遗传多样性的影响,为铁橡栎和干热河谷生态系统的保护和修复提供更全面的资料。
猜你喜欢
疯狂英语·初中版(2023年7期)2023-08-18 05:01:35
安徽农学通报(2021年22期)2021-12-11 20:37:03
安徽农业科学(2021年14期)2021-09-03 09:55:59
小哥白尼(神奇星球)(2021年3期)2021-07-22 03:18:16
小哥白尼(神奇星球)(2021年4期)2021-07-22 03:17:30
炎黄地理(2019年5期)2019-10-20 01:59:19
Zoological Research(2019年3期)2019-06-28 09:13:12
汉语世界(The World of Chinese)(2018年3期)2018-10-22 01:50:04
厦门航空(2018年4期)2018-04-25 10:49:27
乡村地理(2018年4期)2018-03-23 01:54:40
