
肿瘤新抗原在恶性肿瘤治疗和预后分析中的应用
2019-04-01 07:17:02李佳忆杜顺达
世界华人消化杂志 2019年5期
陈 瞧, 李佳忆, 杜顺达
陈瞧, 李佳忆, 中国医学科学院北京协和医学院 北京市 100005
杜顺达, 中国医学科学院北京协和医院肝脏外科 北京市 100730
核心提要: 肿瘤免疫治疗的有效性与免疫细胞对抗原的识别能力密切相关, 利用二代外显子测序技术及生物信息学分析研究肿瘤新抗原, 可为肿瘤的精准治疗和预后预测提供新方向和可能.
0 引言
随着新一代测序成本的下降、多肽设计制作技术的成熟和计算机技术的进步, 找出肿瘤细胞不同于正常细胞的抗原成为可能.这类抗原被称为肿瘤新抗原.肿瘤新抗原可以用来培养靶向性更强的T淋巴细胞, 可以用于免疫检查点封锁疗法应答的预测, 也可以用于治疗性疫苗接种策略, 因而在诊断和治疗中的潜在用途越发重要.
1 肿瘤的抗原和新抗原
根据抗原的特异性程度, 肿瘤抗原分为: 肿瘤特异性抗原(tumor specific antigen, TSA)、肿瘤相关抗原(tumor associated antigen, TAA)和癌症生殖/癌症睾丸抗原(cancer-testis antigen, CTA).CTA是指在肿瘤细胞中过度表达但在正常细胞中低水平表达的一类抗原.TAA能够直接参与致癌过程, 如在维尔姆斯肿瘤1(WT1[1]或Her2/neu)[2]中作为有促生长功能的蛋白; 也可以异常磷酸化后作用于蛋白质翻译后的修饰过程[3].TSA也称为肿瘤新抗原(neoantigen), 是由非同义突变和其他遗传改变引起的、在正常宿主基因组中不编码的一类抗原, 可以表现为由体细胞突变而产生的致癌病毒蛋白或异常蛋白[4].因为TSA诱导产生的T细胞能够更好的适应免疫耐受机制, 因此和抗原有更强的亲和力, 意味着TSA能够更好地引导靶向作用[5].CTA由各种肿瘤类型和生殖组织(例如睾丸, 胎儿卵巢和滋养层细胞)表达, 但在成人的其他正常组织中表达有限, 且不存在于不表达MHC Ⅰ类分子的正常生殖细胞上[6].
TSA、TAA和CTA都被认为是免疫治疗的目标.到目前为止, 大多数癌症疫苗都集中在通常过度表达的TAA上, 因为相同的治疗性疫苗能够治疗各种不同的肿瘤.虽然CTA靶点也在肿瘤之间共享, 但已知它们仅在有限数量的肿瘤类型中表达, 这大大降低了CTA靶向免疫治疗的适用性.
然而, 大多数以TAA为靶向的疫苗尝试都只取得了有限的成功, 并且伴随着较为严重的副作用.这可能是因为TAA也是正常的宿主蛋白质, 因此受到中枢和外周耐受机制的影响.与此同时, 研究发现一些起到治疗作用的TAA特异性T细胞很快出现治疗无效, 其中原因可能是尽管一些TAA特异性T细胞确实避免了阴性选择,但TAA的高亲和性TCR被优先消耗, 剩下的其余TCR的亲和力很低, 我们知道T细胞的细胞毒性和活化与TCR结合亲和力相关, 因此一段时间后治疗效果无法维持.而TSA特异性T细胞的TCR亲和力远高于TAA, TSA在恶性肿瘤治疗中逐步受到重视.
2 肿瘤的免疫效应机制及免疫治疗
肿瘤新抗原的发现提供了一种通过机体的免疫系统治疗肿瘤的思路, 即利用对抗原性异物的识别和清除能力, 杀伤或抑制肿瘤细胞, 达到控制恶性肿瘤的目的, 进而延长恶性肿瘤患者的生存期甚至达到治愈的目的.
机体识别和清除抗原性异物的功能被称为免疫监视[7].肿瘤细胞突变产生的新抗原就是一种抗原性异物,当肿瘤细胞产生新抗原时, 机体的免疫系统能够通过免疫监视发挥抗肿瘤的效应.目前发现的抗肿瘤的免疫效应主要由细胞免疫所介导, 主要包括: 依赖于MHC Ⅰ类分子的CD8+ CTL细胞介导的肿瘤细胞杀伤机制、依赖于MHC Ⅱ类分子的CD4+ Th细胞的抗肿瘤免疫辅助作用、非MHC限制的γδT细胞的肿瘤细胞杀伤机制、CD1d诱导活化的NKT细胞的肿瘤细胞杀伤机制、NK细胞的肿瘤细胞杀伤机制、巨噬细胞、中性粒细胞、淋巴因子等的非特异性杀伤和抑制肿瘤作用等.近年来, 又提出了免疫编辑理论, 认为肿瘤与免疫系统相互作用中, 免疫监视只是其中一个阶段, 随后还要经过免疫平衡和免疫逃逸阶段, 这也对免疫治疗提出了挑战.
在此基础上, 经过长达几十年的肿瘤免疫治疗的探索, 目前肿瘤的免疫治疗主要分为: 主动免疫疗法、被动免疫疗法和免疫检查点抑制疗法等.
主动免疫治疗是通过激活患者自身免疫系统, 利用肿瘤细胞或肿瘤抗原物质诱导机体的特异性细胞免疫和体液免疫反应, 从而增强机体的抗癌能力, 阻止肿瘤的生长、扩散和复发, 最终达到清除或控制肿瘤的目的.被动免疫疗法即细胞免疫疗法, 又称为细胞过继免疫疗法, 其原理是将患者体内自身抗肿瘤的免疫细胞进行体外改造, 让这些细胞具备对癌细胞更有效、更精准的免疫能力.改造后的免疫细胞回输到患者体内后, 它们会定向消灭癌细胞.这类疗法主要利用的免疫细胞包括LAK、DC、CIK、DC-CIK、CAR-T、TCR-T、NK、CAR-NK以及肿瘤浸润性淋巴细胞(tumor infiltrating lymphocytes, TIL)等.其中, 很多免疫细胞因为一些特殊要求或严重的毒副反应等[8]被淘汰, 而利用TIL的细胞过继免疫疗法则受到了关注.TIL是来源于浸润肿瘤组织的淋巴细胞, 它能够在体外良好扩增,抗肿瘤作用强于LAK等细胞, 并且对IL-2的依赖性小[9].有研究证实[9]通过在体外用IL-2诱导激活、扩增TIL, 进而回输TIL能够高效而特异地杀伤肿瘤细胞.免疫抑制检查点疗法则是针对CTLA-4及PD-1的单克隆抗体药物有效地抑制了上述抑制性检查点, 从而促进了抗肿瘤免疫, 产生临床疗效.
如今, 筛选肿瘤特异性抗原已经实现了技术上的突破.通过使用NGS技术[10]和构建算法模型, 外显子测序[11]和转录组测序能得到肿瘤细胞的DNA和RNA的序列信息, 找出可能引起免疫细胞识别的肿瘤性突变.生物信息学工具的发展则提高了肿瘤新生抗原的筛选能力, 基因组大数据和计算机算法加速了肿瘤表位预测以及MHC亲和力预测, 进而推动了个体化肿瘤疫苗的发展.
3 新抗原在肿瘤免疫治疗中的应用
随着肿瘤新抗原检测和预测技术的突破, 基于新抗原的精准免疫治疗热度不断提高, 并且在多种肿瘤的免疫治疗中取得了突破性的进展.
3.1 细胞过继免疫疗法中的应用 细胞过继免疫疗法的关键是过继细胞对肿瘤细胞的识别.随着测序技术的进展, 已经实现了个体化筛选肿瘤抗原和T细胞受体.多项个体化细胞过继疗法研究证实, 通过筛选出能够特异识别肿瘤突变抗原的TIL(多为Th1细胞), 能够实现对肿瘤细胞的有效杀伤.
2011年在一项针对黑色素瘤的临床试验中, 93例黑色素瘤患者接受了从肿瘤患者自身提取出的TIL输注,有20例患者的肿瘤完全消退, 并且100%的患者生存时间超过3年[12].可惜的是, 这项研究对于TIL细胞识别的具体突变抗原仍然是未知的.2013年Robbins团队[13]继而在这批接受TIL治疗的患者中选择了一个肝部结节得到有效缓解的患者, 用传统的cDNA文库筛选分离出了一个突变蛋白PPP1R3B, 并进一步证实从这名患者外周血分离出的T淋巴细胞可以识别这种突变蛋白.另外, 他们还用全外显子测序方法, 找出了突变的新抗原, 并用一种验证MHC结合力的算法验证了可以被三种TIL识别的新抗原[14].
通过以上对方法学和新抗原-TIL相互作用的验证,TIL回输疗法成为可能.全外显子测序则给我们提供了一个全新的思路, 即通过测序获得能被TIL识别的抗原和能识别新抗原的TIL细胞, 直接指导个体化治疗.2014年Rosenberg团队[15]的个案报道实现了对肿瘤进行测序及对靶向免疫细胞进行筛选, 并成功指导治疗的过程.他们招募了一个经过多方案化疗仍然持续进展的已有远处转移的胆管癌患者, 用全外显子测序的方法检测出患者癌症细胞中的26个非同义突变基因, 其中一个突变基因表达erbb2突变相互作用蛋白(ERBB2IP).他们发现从该患者肿瘤组织中分离出的肿瘤浸润淋巴细胞(TIL)含有的Vβ22+CD4+T辅助细胞1(Th1)细胞能够特异识别ERBB2IP.随后该团队在体外扩增这种可以特异识别突变蛋白的T细胞, 用过继细胞回输的方法输入含25%浓度的有突变识别特异性的Th1细胞, 在7 mo随访时, 肺和肝的转移灶共缩小了30%.患者病情在随访至13 mo时依然稳定.随后出现肺部进展, 再次向患者输入含>95%浓度的有突变识别特异性的Th1细胞, 在1 mo和6 mo随访时, 研究者观测到患者的肺转移灶再次缩小.
2015年Rosenberg团队[16]对10名转移性消化道肿瘤的患者的肿瘤细胞进行了外显子测序, 并证实了从9名患者体内分离出的CD4+或CD8+细胞可以识别3种经测序得到的新抗原.并且一名患者体内分离出的含白细胞抗原-C*08:02-限制性T细胞受体的CD8+T细胞可以识别在很多消化道肿瘤中共有的突变KRAS G12D.2016年这一团队利用从这些结直肠癌患者中分离出的4种靶向KRAS G12D突变的CD8+T细胞, 在实验室中扩增到一定的数量后, 通过静脉回输含75%浓度的TIL细胞到最后这名患者体内.结果发现, 在回输后40天, 患者所有7个转移性肺结节消退, 且维持了9 mo.9 mo后, 其中一个病变出现进展, 通过手术切除后维持了至少8 mo临床无病状态.因此靶向共有突变的TIL细胞为治疗的广泛应用前景广泛[17].
3.2 个体化疫苗中的应用 癌症疫苗是通过向患者注射癌症特异性元素以引发针对肿瘤的免疫应答, 提高了免疫系统识别和杀伤肿瘤细胞的能力[18].疫苗主要分为树突细胞[19,20]、DNA[21]、RNA[22]和肽类[23], 各自有优势和劣势.在体内, 疫苗的组分激活专职抗原呈递细胞, 包括树突细胞等, 树突细胞接收并处理引入的抗原, 然后迁移到局部淋巴结; 进入淋巴结后树突细胞通过MHC Ⅰ类或Ⅱ类分子在细胞表面展示抗原.TCR与MHC Ⅰ类分子/肽复合物连接后, 新抗原特异性T细胞被激活, 经历增殖并分化成CD8+细胞毒性T细胞, 然后离开淋巴结并靶向结合肿瘤表面显示的抗原.类似地, CD4+T细胞也可以被激活.与传统疫苗类似, 肽疫苗可以由许多肽和佐剂组成, 佐剂是一类旨在加强对疫苗的免疫应答的免疫刺激剂.到目前为止, 肽疫苗的耐受性良好, 治疗相关的不良事件很少.研究者们利用靶向肿瘤特异性新抗原配制和注射突变的合成肽来制造个性化的治疗性癌症疫苗来治疗恶性肿瘤.
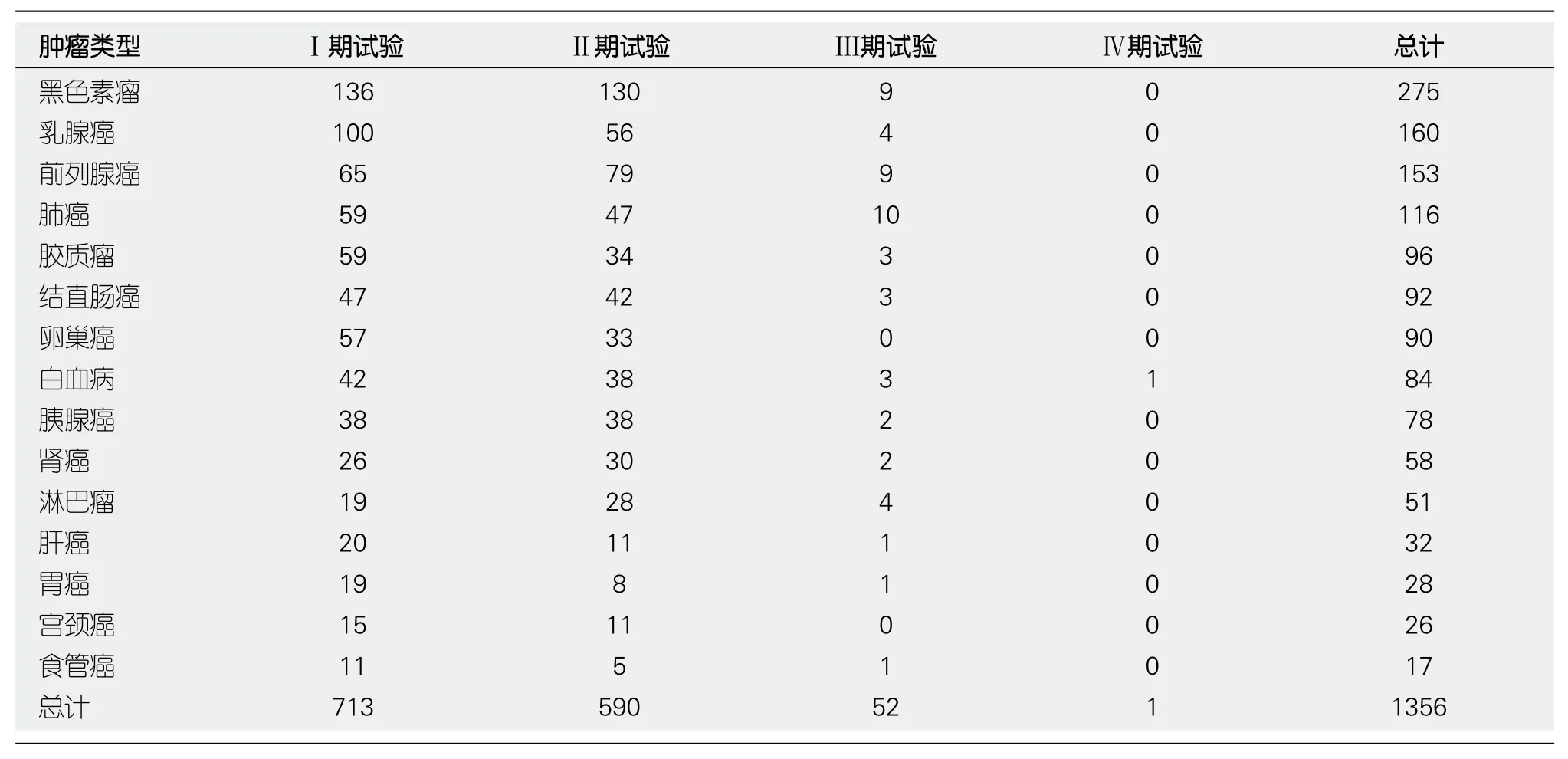
表1 包括处于招募前、招募中、终止、撤销、完成等状态下的所有临床试验(截止到2017-12)
有一部分治疗性癌症疫苗可能由于有缺陷的抗原靶标, 如免疫原性低, 抗原特异性低或肿瘤细胞表达水平低, 或弱佐剂而失败[24].但也有成功研制的疫苗, 如:黑色素瘤疫苗[25-27]、胰腺癌疫苗[28]、多抗原疫苗[29].
关于肿瘤疫苗的研究层出不穷, 但是多数还停留在前期研究或者动物水平表1也有完成了Ⅳ期试验的疫苗, 如坦佩雷大学医院研究的治疗慢性淋巴细胞白血病的肺炎球菌多糖疫苗.
4 新抗原辅助下对肿瘤预后的预测
肿瘤对免疫检查点抑制治疗的反应与新抗原的数目和免疫原性相关[30,31], 免疫检查点抑制治疗前进行基因测序能够评估治疗的反应.2013年在一例对抗-CTLA4 Ipilimumab单抗疗法有良好反应的Ⅳ期黑色素瘤患者中, Van[32]进行外显子测序和T细胞反应分析, 发现了体内出现了靶向针对两个新抗原的T细胞.2016年McGranahan等[31]对接受PD-1抑制剂Pembrolizum单抗治疗的进展期非小细胞肺癌患者进行了外显子测序, 发现肿瘤新抗原数量越多且瘤内异质性越少的患者对免疫检查点抑制治疗敏感性更高, 并且总生存时间显著延长.这显示了新抗原在免疫检查点抑制剂进行免疫治疗的效果及预后预测方面的作用.
肿瘤新抗原还影响肿瘤患者对免疫检查点抑制治疗的耐药性的产生.2017年Anagnostou等[30]对接受抗PD-1/抗CTLA-4抗体治疗后产生耐药性的非小细胞肺癌患者的肿瘤细胞进行全外显子测序, 发现出现耐药性的肿瘤丢失了一部分新抗原并产生了新的突变, 这些丢失的新抗原与剩余的新抗原相比, 有更强的MHC结合力.
此外, 对一些恶性肿瘤的分析也发现, 新抗原与肿瘤患者的无进展生存时间和总生存时间相关.Balachandran等[33,34]通过对胰腺癌肿瘤细胞进行外显子测序、MHC结合力、TCR测序及T细胞亚群分析发现,新抗原数目、新抗原与MHC的结合力以及CD3+CD8+型T细胞多的个体生存期显著延长.Choudhury等[35]在肌层浸润性膀胱癌患者中发现, 表达更高的新抗原数目和更低的TCRβ多样性的患者可以获得更长的无复发生存期.Matsushita等[36]则在肾脏透明细胞癌患者中发现, 同时表达更高的新抗原数目和HLA分子的患者有更好的临床预后.Strickland等[37]也在高级别浆液性卵巢癌的患者中发现有更高新抗原负荷的患者总生存率更高.
这些研究提示对肿瘤进行基因测序在一定程度上可以预测其生存期, 并且测序结果可能提示对相应治疗的疗效反应.
5 结论
新抗原带来的个性化免疫治疗可能治愈高风险进展患者, 迄今已有较多临床研究取得了可喜的成绩.然而未来还存在巨大的挑战.我们需要更多关于新抗原处理和呈现的数据、改进预测算法, 以选择最理想的免疫原性表位.肿瘤过继免疫疗法中使用的肿瘤浸润淋巴细胞主要来源于肿瘤组织, 制备的过程有创伤且需要一定时间.而疫苗的生产需要更快、更经济、更高效的方法,同时寻找最佳新抗原靶点也是需要解决的问题.探索几种免疫疗法的联合应用, 可以使疗法适用更为广泛肿瘤患者, 并且提高疗效.在恶性肿瘤的疗效预测和生存期分析方面, 新抗原起了一定作用, 发现更为精准的预测靶点, 将为肿瘤的精准化治疗带来新的希望.
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29 01:57:52
中国生殖健康(2020年4期)2021-01-18 02:58:10
中国生殖健康(2020年7期)2020-12-10 07:48:51
中国生殖健康(2018年4期)2018-11-06 07:12:16
海南医学(2016年8期)2016-06-08 05:43:00
西南医科大学学报(2015年1期)2015-08-22 13:01:46
医学研究杂志(2015年6期)2015-07-01 17:41:11
癌变·畸变·突变(2015年3期)2015-02-27 06:15:07
湖北农业科学(2014年11期)2014-09-10 18:06:07
郑州大学学报(理学版)(2014年2期)2014-03-01 04:20:57
