
基于高光谱和深度迁移学习的柑橘叶片钾含量反演
2019-04-01 12:28岳学军凌康杰王林惠岑振钊刘永鑫
农业机械学报 2019年3期
岳学军 凌康杰 王林惠 岑振钊 卢 杨 刘永鑫
(华南农业大学电子工程学院, 广州 510642)
0 引言
钾(K)是柑橘生长过程中必需的营养元素之一,活性极高,对柑橘果实发育和品质形成起到十分重要的作用[1]。我国是柑橘生产大国,快速无损测定柑橘叶片钾含量对柑橘生长健康状况、营养元素胁迫状况、柑橘产量预测和柑橘树的栽培管理、变量施肥施药、农机具研发等具有重要意义[2]。
由于传统的作物钾含量检测方法存在耗时、费力、过程冗长繁琐、时效性差等弊端,因此研究快速无损的钾含量检测方法成为热点[3]。随着高光谱技术的迅速发展,国内外研究者深入研究了作物营养成分光谱检测方法,但主要集中于氮素含量研究,对钾素含量研究较少[4-6]。MENESATTI等[7]基于近红外光谱对柑橘叶片包括钾在内的多种营养元素进行了估测,但缺乏对柑橘不同物候期的高光谱分析与建模。刘艳丽等[8]基于高光谱技术分别建立了iPLS和siPLS模型的柑橘花钾素营养估测模型,获得预测集相关系数最高为0.885 5。朱西存等[9]使用ASD FieldSpec3光谱仪测定苹果花期冠层高光谱数据,并对原始光谱进行了不同形式的光谱变换,获得一阶微分下多元线性回归(Multiple linear regression,MLR)模型的相关系数为0.985 1,平均相对误差为9.8%。赵化兵等[10]对梨树鲜叶的可见/近红外反射光谱进行了S-G(3)滤波,所建立的PLSR钾素含量模型的验证集和预测集决定系数为0.722 7和0.679 1,预测的平均相对误差为6.81%。以上方法中均没有涉及不同物候期光谱特征迁移,研究对象局限于某一时期,没有关注果树不同生长期的营养状况,由于叶片营养成分会不断变化,单一物候期的营养元素含量预测对整株果树生长周期营养状况监测不具备指导作用[11]。同时上述方法所建立的单模型单数据集在特征提取和泛化能力上存在不足,均需收集相应数量的化学测量真实值作为模型训练监督标签,存在成本高、耗时间长、过程繁琐、工作量大缺点,不能实现不同目标域之间的特征迁移和融合[12-14]。
本文采用深度迁移学习方法对柑橘树4个不同物候期的高光谱数据进行自适应特征提取和降维,迁移和融合多种特征,以此建立SSAE-DLNs柑橘叶片钾含量估测模型。
1 材料与方法
1.1 柑橘树种植管理与叶片采集
选择广州市萝岗区蟹庄村柑橘种植示范园4年生的109株柑橘作为试验样本,并进行科学规范的种植管理,保证温度在12~20℃,灌水量1 000 mm左右,pH值为4~5,相对空气湿度为80%左右。采用纯度为45%的钾肥分别于萌芽期、稳果期、壮果促梢期和采果期进行施肥,施肥时间为2015年2、4、6、8月下旬,施肥量占全年施肥总量的30%、30%、25%、15%。保证水肥合理、稳定,灌溉频次为每周2次。分别在2015年4月24日(萌芽期)、6月22日(稳果期)、8月23日(壮果促梢期)、10月23日(采果期)采集柑橘叶片,在每棵树东南西北4个方位的上下两层共采集8片鲜叶,采集位于顶梢起向下数第3和第4片大小均匀的健康叶片。
1.2 高光谱测定
采用美国ASD FieldSpec 3光谱仪采集柑橘鲜叶的光谱反射值,光谱测量范围为350~2 500 nm,光谱数据输出间隔为1 nm,故柑橘叶片的光谱数据为2 151维矢量。具体操作是从叶片基部将叶片完整剪下,平铺于反射率近似为零的黑色橡胶上,采用ASD公司提供的Pro Lamp(A128932)人工光源通过手柄安装在采集支架上,入射光纤探头垂直正对待测叶片,距离叶片6 cm,视场直径0.52 cm,顶角30°,分别在每片叶子的叶尖、叶中和叶基附近采集3次[15]。实际光谱反射值是叶尖至叶基光谱反射值的平均值。每次测量前均用白板校正,采集暗电流25次,白板10次[16]。同一棵树同一生长时期的8片叶片光谱反射率的均值作为柑橘叶片样本的光谱反射率,不同生长时期的同一棵树视为不同样本,共有436个柑橘叶片样本。不同物候期109棵柑橘叶片样本的反射光谱如图1所示。

图1 不同物候期柑橘叶片样本反射光谱Fig.1 Relectance spectra of citrus samples at different growth stages
1.3 叶片钾含量测定

图2 不同物候期钾质量分数箱线图Fig.2 Boxplot of potassium content at different growth stages
光谱测定后,选用火焰光度法测定柑橘叶片钾含量[17],测定方法为将每株柑橘的8片鲜叶均匀混合,待测样品经消煮或浸提,然后稀释,用湿灰化法获取待测容液,再用原子吸收光谱法进行钾素检测。钾质量分数统计数据如图2所示。从图2可知,不同物候期的柑橘叶片钾含量由大到小依次为采果期、萌芽期、壮果促梢期、稳果期。
1.4 高光谱数据预处理方法
在高光谱数据获取过程中,由于测定环境、仪器噪声等因素影响,不可避免存在多重共线性、吸光度的非线性、附加散射变动和基线变动。原始光谱维度高,相邻波段存在较大相关性,造成数据冗余[18]。本文分别采用连续投影算法(Successive projections algorithm,SPA)和小波去噪对原始光谱进行预处理。
SPA是一种常用的光谱特征波长选取算法,能使从光谱变量中选出的变量组共线性最小、冗余信息最少。柑橘在不同物候期对钾素的需求不同,不同物候期的光谱反射率综合表征其生理和物理化学组分状态,通过SPA降维方法,提取光谱中敏感波段[19]。
利用小波去噪方法,首先选定小波基函数和分解层次对原始数据进行分解,然后根据各层阈值去除分解后小波系数较小的成分,最后通过小波逆变换重构信号[20]。试验中采用正交试验和极差分析对小波去噪结果进行筛选,选出小波去噪参数最优水平组合。
1.5 建模原理与评价标准
堆栈稀疏编码机(Stacked sparse autoencoder,SSAE)是一种多层神经网络,能自适应获取特征层,SSAE的特征分享层能够学习到不同任务之间的特征和噪声共性模式,既能专注于特定特征,也能提供共性特征[21]。本文将堆栈稀疏自动编码机堆叠起来组成特征分享层[22],用于捕获多任务中共同特征,减少目标函数岭、脊和鞍点,加快迭代速率,优化和提高深度学习模型预测精度和泛化能力。
设SSAE的稀疏度为ρ,网络激活函数为σ,整个SSAE定义KL(Kullback-Leibler)散度[23]为
(1)

(2)

m——第j个神经元与它前向相连接的神经元数量
z——神经元加权输入值
设SSAE网络权重矩阵为W,网络偏置向量为b,第l层神经元数量为sl,隐含层总数为nl,调和因子为β,原损失函数为J(W,b),稀疏损失函数Js(W,b)为
(3)
DLNs深度学习模型在特征提取上具有自适应性,并且能够融合不同分布概率的光谱特征,提高模型精度、稳定性、泛化能力和迁移能力[24-26]。设源域为Ds,目标域为DT,假设函数为h,正则函数为Ω,最小化损失函数Ψ为
(4)
式中Θ——Ds迁移至DT的网络参数
Xj——目标域第j个输入矩阵
yj——目标域第j个输出向量
nT——目标域样本总数
从总样本数中随机选取80%的样本(349个样本)作为校正集(Calibration set),用于训练模型,剩余20%(87个样本)作为验证集(Validation set),用于评估模型,建模评估采用模型决定系数R2和均方根误差(Root mean square error,RMSE)两个指标[27]。试验数据处理与建模流程为:先进行SPA降维,将提取的特征光谱用于建立MLR模型;同时采用小波去噪和SSAE特征提取和迁移方法,分别建立DLNs模型,对比分析基于传统单一特征与SSAE迁移特征下模型的最终效果。SSAE-DLNs试验中采用基于随机梯度下降的BP算法训练网络,设置传输函数为logisg[28],最大迭代次数为1 200,最佳神经元数量采用格点搜索法(Grid search)和交叉验证法(Cross validation)选得最优值。每个试验方案均取20次运行结果的均值作为最终记录。
2 结果与分析
2.1 最优SPA和MLR建立与验证
通过交叉验证方法,选取最优SPA降维结果。波长按重要性由大到小排列,试验结果如表1所示。

表1 不同物候期钾质量分数特征波段Tab.1 Characteristic bands of potassium content at different growth stages
由表1可知,钾素明显存在特征光谱漂移现象,不同生长期的特征光谱分布情况不同,特征波段数量多、跨度广、构成复杂。随着生长期推移,钾素反射光谱有部分重要的特征波段向高波段漂移。萌芽期特征波长主要集中在1 000 nm内,采果期有部分特征光谱漂移至1 935~2 007 nm和2 496~2 500 nm。
将原始反射光谱进行一阶微分,结果如图3所示。在490~530 nm波段,柑橘叶片钾含量越低,蓝边面积越大,这是由于柑橘缺钾素时,叶绿素含量降低,反射蓝绿光增加;在540~660 nm波段,一阶微分越大,钾含量越大,对应原始反射光谱650 nm处有一个红光吸收谷,随钾含量降低,叶绿素含量下降,光合作用利用的红光减少,吸收谷变浅。780~1 100 nm波段范围内,一阶微分的和越大,钾含量越高,可能与缺钾导致叶片表皮细胞发生变化,叶片厚度增加,叶片栅栏、海绵组织的细胞收缩和局部破裂有关。1 900~2 010 nm波段范围内,一阶微分的和越大,钾含量越高,其内在的生理生化机理还有待研究[29-30]。

图3 柑橘叶片反射光谱一阶微分Fig.3 First order derivative of relectance spectra for citrus
将SPA降维后的数据作为输入量,建立MLR模型[31],结果如表2所示。

表2 不同物候期MLR建模性能评估Tab.2 Performance evaluation of MLR model at different growth stages
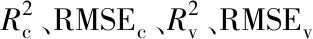

2.2 小波去噪和DLNs建模结果与分析
小波分解过程中涉及小波基函数、分解层数以及光谱重构过程中的阈值方案、阈值调整方案等参数,据文献表明,分解层数大于7时,对建模效果没有明显的提升作用[32]。由于小波去噪涉及因素多,采用正交试验,根据文献[33],正交试验的预选小波基函数选取haar、db4、sym8、coif2,能达到良好效果。正交试验的因素水平[34]如表3所示。

表3 正交试验因素水平Tab.3 Factors and levels of orthogonal test
从表3中可以看出,该正交试验包括3因素4水平和1因素3水平两类。因此,采用拟水平法,将因素D的第3水平进行重复,构成4因素4水平,直接采用正交表L16(45)进行正交试验。虽然拟水平后的表对所有因素来说不具有均衡搭配性质,但仍具有部分均衡搭配的性质,因此拟水平法仍保留了正交表的优点。小波去噪参数正交试验结果见表4。针对5个物候期,采用极差分析法对小波去噪正交试验的结果进行分析,试验结果见表5。
从表4知,第14组水平组合的校正集和验证集模型决定系数均达到最大,分别为0.887 9、0.875 2,比基于原始光谱建模的R2提高了9.25%和8.42%;校正集和验证集模型的均方根误差达到最小,分别为0.343 6、0.314 8,比基于原始光谱建模的RMSE降低了56.95%和61.89%,与表5的分析一致,即最优水平组合是A4B2C3D1,即小波基函数为coif2,分解层数为3,阈值方案为sqtwolog,噪声估计方案为one。本试验采用A4B2C3D1方案对所有光谱进行去噪。
2.3 SSAE和DLNs建模结果与分析
2.3.1SSAE中光谱特征状态
SSAE试验中权重状态见图4。由图4可知,SSAE能够对不同物候期的光谱自适应获取光谱非线性特征,物候期间特征光谱存在交集。在前层单元对后层单元的关系中,萌芽期、稳果期、壮果促梢期、采果期的SSAE神经元归一化活跃度超过0.5,分别占总神经元数的23.36%、37.08%、3.10%、20.96%,平均活跃度分别为0.361 5、0.399 0、0.280 7、0.156 4,4个物候期的前层单元活跃神经元主要分布于67~86、100~105、135~145、14~39、69~81和150~160号神经单元上。
2.3.2同一物候期特征层间迁移与融合
研究柑橘同一物候期光谱在SSAE中每层特征迁移融合与层数之间的关系,依次融合基准层和特征迁移层,得到深度学习网络模型性能如图5所示(图中*表示极大值,横坐标0表示基准层)。
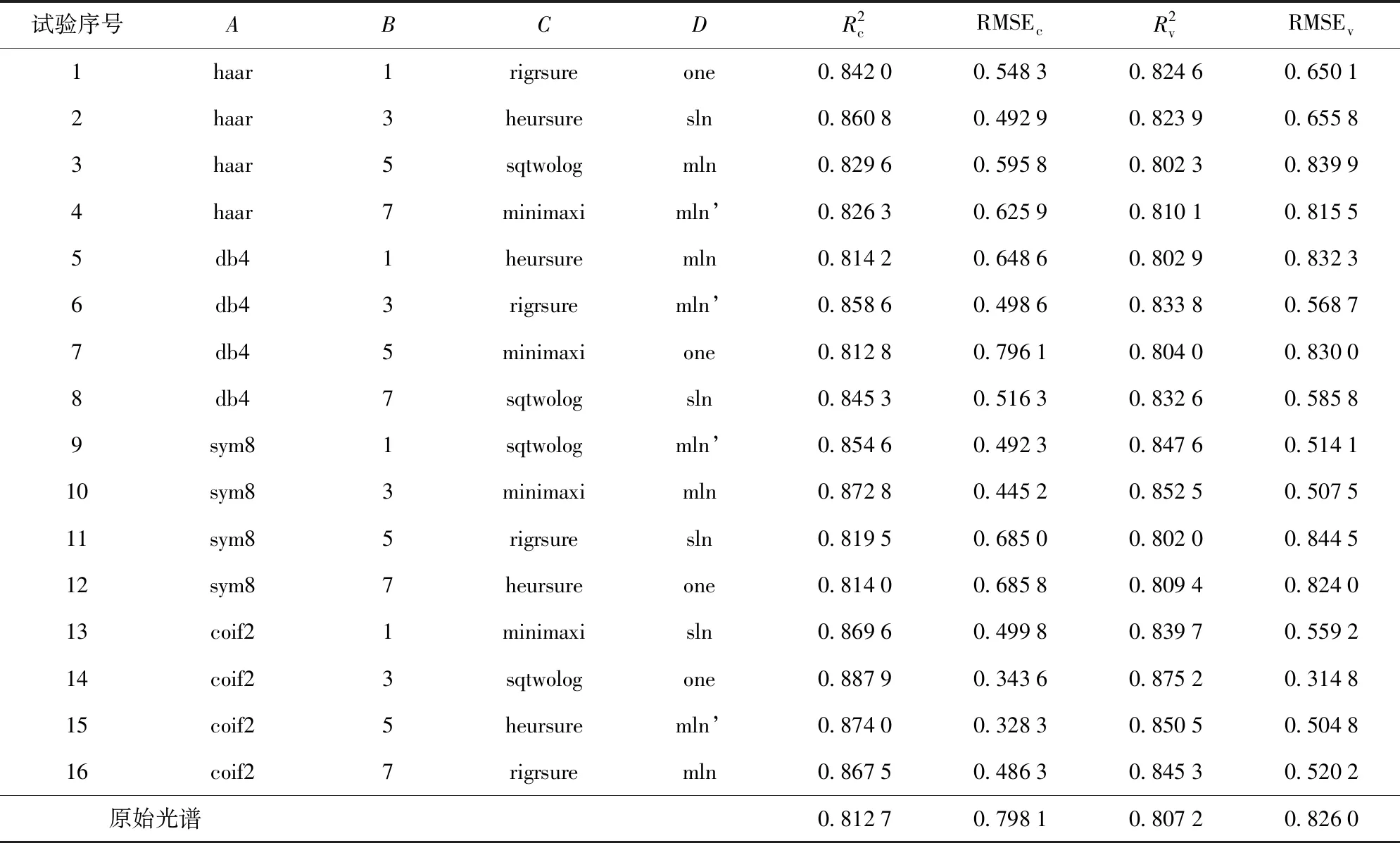
表4 小波去噪正交试验结果Tab.4 Grouping and result of wavelet denoising orthogonal test
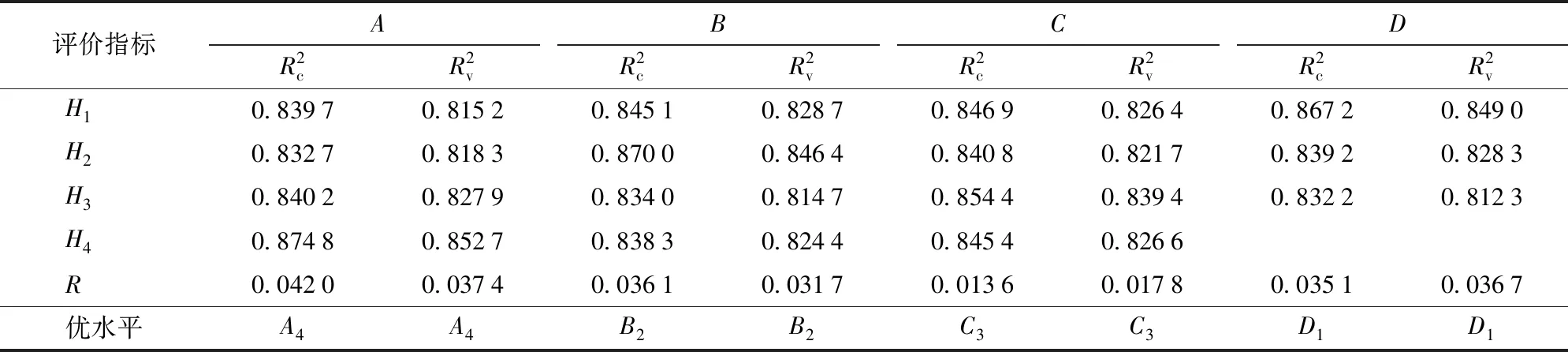
表5 小波去噪正交试验结果极差分析Tab.5 Analysis result of wavelet denoising orthogonal test
注:H1~H4为某一水平的评价指标均值;R为极差。

图5 特征层数对模型性能影响Fig.5 Influence of characteristic layers on model performance
由图5可知,随着迁移层数增加,意味着增加了特征数,DLNs模型性能得到提高。当因SSAE特征迁移层数增加而引入的噪声影响大于所引入的特征数时,模型性能开始下降且不稳定。在特征层数迁移中,萌芽期、稳果期、采果期超过极大值后,开始迅速下降。壮果促梢期超过极大值后在第8层出现第2极大值,考虑到迁移层数过大会引入高增量噪声和增加模型复杂度,影响模型鲁棒性,故壮果促梢期最优层数选择4。所以试验中萌芽期、稳果期、壮果促梢期、采果期最优迁移层数分别为3、1、4、3,R2分别为0.899 9、0.859 8、0.886 9、0.854 7,比原始基准层分别提高了19.82%、9.45%、21.49%、7.21%。
2.3.3不同物候期光谱特征迁移与融合


图6 迁移域数与决定系数关系Fig.6 Relationship between number of transfer domains and determination coefficient
由图6可知,SSAE不同物候期的迁移特征能明显提高目标域的模型性能,1个物候期迁移域便能明显提高基准目标域的性能。随着迁移域增加,目标域性能提高速率减缓,逐步达到饱和状态,意味着新迁移域所引入的相关特征对目标域贡献率减低而变得相对不明显。由图6可知,试验中最佳迁移域为4,对应的萌芽期、稳果期、壮果促梢期、采果期最佳R2分别为0.877 2、0.898 1、0.904 9、0.889 4,比BTD分别提高了16.80%、14.32%、23.96%、11.56%。
根据以上分析,本文对萌芽期、稳果期、壮果促梢期、采果期4个物候期试验选择SSAE的迁移层数分别为3、1、4、3,每个目标域的迁移域数为4。结合2.2节部分,对所有光谱进行小波去噪后,研究和对比多种不同光谱形式的DLNs建模,该模型的最佳稀疏度采用留一交叉验证法来选定,试验结果见表6。
从表6可知,一阶导数谱r′建模效果最好,萌芽期、稳果期、壮果促梢期、采果期和全生长期的校正集模型决定系数分别为0.899 2、0.889 9、0.883 8、0.872 7和0.898 8,稀疏度分别为0.141 1、0.163 3、0.118 9、0.185 6和0.207 8,验证集模型决定系数分别为0.865 1、0.870 4、0.855 1、0.858 0和0.877 1;倒数谱1/r建模效果最差,萌芽期、稳果期、壮果促梢期、采果期和全生长期的校正集模型决定系数分别为0.815 7、0.791 6、0.801 2、0.781 9和0.801 2,验证集模型决定系数分别为0.809 6、0.780 6、0.791 6、0.775 1和0.800 9;原始光谱r的建模效果仅次于一阶导数谱r′,而对数谱lgr和二阶导数谱r″的建模效果皆低于原始光谱r。
2.4 其他模型建模对比分析
本试验将最优的基于SSAE-DLNs和其他模型进行对比,目前在基于高光谱反射率对营养元素反演中的主流模型主要包括支持向量回归(Support vector regression,SVR)、偏最小二乘法回归(Partial least square regression,PLSR)、广义神经网络(General regression neural networks,GRNN)、逐步多元线性回归(Stepwise multiple linear regression,SMLR)等。将全生长期的光谱经过小波去噪后的一阶导数谱r′导进这4个模型,模型参数分别调节到最优,结果如图7所示。

表6 不同光谱变换形式的模型性能评估Tab.6 Model performance evaluation on calibration set with different transformations of spectral data
注:r、r′、r″、1/r、lgr分别为原始光谱、一阶导数谱、二阶导数谱、倒数谱、对数谱。

图7 不同建模方法下钾质量分数实测值和预测值的比较Fig.7 Comparison of measured and predicted potassium contents by using different modeling methods
图7不同模型与表6中最优SSAE-DLNs模型相比,性能均有不同程度下降,主要原因是钾素特征光谱中含有大量复杂的非线性特征,而较为简单的模型无法得到特征变量间更复杂的关系。SSAE模型能够自适应不同物候期的光谱特征并能相应提取有效成分,能对小波去噪后的一阶导数谱具有最大解释能力。DLNs模型能对不同物候期的光谱域进行特征迁移和融合,可以比单模型单任务带来更丰富的信息。因此SSAE-DLNs模型具有更好的预测精度和泛化能力,能自适应不同时期的光谱反演任务。
3 结论
(1)为了降低数据噪声对深度学习网络的影响,采用小波去噪方法对光谱进行预处理。通过正交试验和极差分析,小波去噪最优水平组合是A4B2C3D1,即小波基函数为coif2,分解层数为3,阈值方案为sqtwolog,噪声估计方案为one。
(2)试验采用了SSAE模型实现柑橘不同物候期特征提取和迁移,自适应获取光谱中非线性特征;采用了DLNs模型进行物候期间的光谱特征融合和钾含量反演。SSAE模型的前层单元中神经元平均活跃度分别为0.361 5、0.399 0、0.280 7、0.156 4。SSAE中萌芽期、稳果期、壮果促梢期、采果期的最优迁移层数分别为3、1、4、3,模型决定系数分别为0.899 9、0.859 8、0.886 9、0.854 7,比原始基准层分别提高了19.82%、9.45%、21.49%、7.21%。每个目标域的最优迁移域数为4,萌芽期、稳果期、壮果促梢期、采果期模型最佳决定系数分别为0.877 2、0.898 1、0.904 9、0.889 4,比基准目标域分别提高了16.80%、14.32%、23.96%、11.56%。
(3)比较了不同光谱变换形式的建模效果,将原始光谱分别变换为多种光谱形式,导入SSAE-DLNs模型训练,试验结果表明,一阶导数谱r′建模效果最好,萌芽期、稳果期、壮果促梢期、采果期和全生长期的校正集模型决定系数分别为0.899 2、0.889 9、0.883 8、0.872 7和0.898 8,稀疏度分别为0.141 1、0.163 3、0.118 9、0.185 6和0.207 8,验证集模型决定系数分别为0.865 1、0.870 4、0.855 1、0.858 0和0.877 1。光谱变换形式的建模效果由大到小依次为r′、r、r″、lgr、1/r。

(5)不同物候期的柑橘叶片钾含量由大到小依次为采果期、萌芽期、壮果促梢期、稳果期。不同物候期施肥量表明不同时期叶片钾含量变化呈现一定的规律,但与施肥量之间并不是严格线性关系。
猜你喜欢
农业工程学报(2022年12期)2022-09-09
热带作物学报(2022年6期)2022-07-08
中国特种设备安全(2022年3期)2022-07-08
科技风(2021年19期)2021-09-07
今日农业(2020年19期)2020-12-14
——以长白山区为例
东北林业大学学报(2020年9期)2020-10-21
农业灾害研究(2020年8期)2020-05-11
电子制作(2019年13期)2020-01-14
东坡赤壁诗词(2019年5期)2019-11-14
世界热带农业信息(2019年3期)2019-06-20
